原产地和入侵地不同细胞型加拿大一枝黄花幼苗期形态特征的比较
2018-04-08许京璇薛丽芳
许京璇, 薛丽芳, 强 胜
(南京农业大学 杂草研究室, 江苏 南京 210095)
加拿大一枝黄花(SolidagocanadensisLinn.)隶属一枝黄花属(SolidagoLinn.)[1],为世界性外来入侵性杂草之一[2-6],原产于北美,广泛分布在加拿大北部至美国南部的东部沿海及中部。目前,该种广泛分布于中国华东、华中、华北和西南等地,并有进一步扩散的趋势[7-9]。在入侵地,加拿大一枝黄花的竞争能力较强,能够适应广泛的环境条件[10-11],可在多样化的栖息地迅速建立种群和传播。加拿大一枝黄花入侵后,入侵地的生物多样性降低,引起入侵地生境与原产地生境的生物均质化[12],破坏入侵地原来的生态系统结构与功能,导致入侵地原有物种局部灭绝,严重破坏入侵地的生物多样性和生态系统的稳定性。加拿大一枝黄花能够迅速入侵的一个重要原因是其植株可产生大量随风传播的种子,并且其种子能够快速萌发并长成具有竞争优势的幼苗,最终定植在裸地或人工干扰生境中[11,13]。
学术界对加拿大一枝黄花的分类和命名一直存在争议[6]。目前,加拿大一枝黄花的分类主要依赖Melville等[14]在1982年提出的形态学性状指标。染色体研究结果表明:加拿大一枝黄花在原产地和入侵地至少有二倍体(2n=18)、四倍体(2n=36)和六倍体(2n=54)3种染色体倍性[15]。由于染色体倍性和产地不同,原产地和入侵地不同细胞型加拿大一枝黄花的形态存在差异,且染色体倍性差异可能是导致加拿大一枝黄花复合种鉴定困难的重要原因。
从种子到幼苗是植物生活史中最关键的阶段之一,对于植物的定植、拓展或占据生存空间具有决定性意义[16-17]。子叶(cotyledon)是植物种子和幼苗的重要组成部分[18-19],子叶生长是植物生活史的重要阶段,并且,子叶的形态特征能够反映植物对环境变化的生存对策[20-22]。此外,幼苗的建立和存活对植物的生存和竞争具有重要意义。
鉴于此,以原产地和入侵地加拿大一枝黄花的二倍体、四倍体和六倍体幼苗为实验材料,对其子叶期幼苗和子叶的形态特征,以及真叶期(包括第一和第二真叶期)第一真叶的形态特征进行了比较分析,以期为一枝黄花属植物的分类提供更多实验依据,并为进一步阐明加拿大一枝黄花的入侵机制提供参考。
1 材料和方法
1.1 材料
供试的加拿大一枝黄花原产地的二倍体(NA2x)、四倍体(NA4x)和六倍体(NA6x)以及入侵地的二倍体(IN2x)、四倍体(IN4x)和六倍体(IN6x)分别采自加拿大魁北克(北纬43°07′12″、西经131°53′38″)、美国克林顿(北纬32°03′36″、西经118°31′12″)、美国克利夫兰(北纬36°10′48″、西经84°04′12″)、俄罗斯符拉迪沃斯托克(北纬46°45′36″、东经71°17′24″)、中国九江(北纬29°46′48″、东经116°08′24″)以及中国南京(北纬32°06′21″、东经118°52′29″)。将原产地和入侵地不同细胞型加拿大一枝黄花5个单株的种子分别置于-20 ℃条件下保存、备用。
1.2 方法
1.2.1育苗方法每个单株选取大小相近且籽粒饱满的种子30粒,用体积分数75%乙醇浸泡1 min,经无菌水漂洗3次后,分别播种在盛有灭菌土的塑料杯(口径7.5 cm、高度8.5 cm)中,置于温度25 ℃、空气相对湿度80%、光照强度110 μmol·m-2·s-1的恒温培养箱内进行培养。同一单株的种子播种在1个塑料杯中,原产地和入侵地各细胞型分别播种5个塑料杯,视为5个重复。
1.2.2指标测定方法种子萌发后第7天(子叶期),在每个塑料杯中选取长势均匀的幼苗5株,用无菌水将根部泥土洗净,使用Olympus SZX7体式显微镜(日本Olympus公司)进行观察,使用Olympus DP71 CCD(日本Olympus公司)进行拍照,并使用Image-Pro Plus软件测量幼苗的全株长、根长、地上部长度及茎直径。其中,全株长为幼苗根尖到幼苗顶端的距离;根长为幼苗地上部与根的分界线到根尖的距离;地上部长度为幼苗地上部与根的分界线到幼苗顶端的距离;茎直径为下胚轴最宽处的直径。
在子叶期,用刀片沿子叶叶柄基部将子叶切下,轻轻展平,使用Image-Pro Plus软件测量子叶的长度、宽度和面积;在第一真叶期(第一真叶完全展开、第二真叶尚未展开)和第二真叶期(第二真叶完全展开),采用相同方法测量第一真叶的长度、宽度和面积。每个塑料杯选取长势均匀的幼苗5株进行测量。
根据测量结果计算幼苗根长与全株长的比值以及子叶和第一真叶的长宽比。
1.3 数据统计和分析
采用EXCEL 2010软件进行数据整理和分析;采用SPSS Statistics 21.0软件进行统计学分析,其中,差异显著性检验采用Duncan’s新复极差法。
2 结果和分析
2.1 子叶期幼苗和子叶形态特征的比较
2.1.1幼苗形态特征的比较观察和测量结果(图版Ⅰ-A和表1)表明:在子叶期,原产地和入侵地加拿大一枝黄花四倍体和六倍体的全株长显著大于二倍体,其中,原产地四倍体和六倍体的全株长分别较二倍体增加70.19%和76.24%,且四倍体和六倍体间差异不显著;入侵地四倍体和六倍体的全株长分别较二倍体增加20.50%和28.29%,且六倍体的全株长显著大于四倍体。并且,入侵地加拿大一枝黄花二倍体、四倍体和六倍体的全株长显著大于原产地,分别较原产地增加58.79%、12.43%和15.59%。
在子叶期,原产地和入侵地加拿大一枝黄花四倍体和六倍体的地上部长度显著大于二倍体,其中,原产地四倍体和六倍体的地上部长度分别较二倍体增加35.51%和43.00%,且四倍体和六倍体间差异不显著;入侵地四倍体和六倍体的地上部长度分别较二倍体增加13.36%和14.62%,且四倍体和六倍体间差异不显著。并且,入侵地加拿大一枝黄花二倍体、四倍体和六倍体的地上部长度显著大于原产地,分别较原产地增加33.82%、11.94%和7.26%。
在子叶期,原产地和入侵地加拿大一枝黄花四倍体和六倍体的根长显著大于二倍体,其中,原产地四倍体和六倍体的根长分别较二倍体增加116.24%和120.06%,且四倍体和六倍体间差异不显著;入侵地四倍体和六倍体的根长分别较二倍体增加27.08%和40.86%,且六倍体的根长显著大于四倍体。并且,入侵地加拿大一枝黄花二倍体、四倍体和六倍体的根长显著大于原产地,分别较原产地增加91.72%、12.67%和22.72%。
在子叶期,原产地和入侵地加拿大一枝黄花四倍体和六倍体的根长与全株长的比值显著大于二倍体,其中,原产地四倍体和六倍体的根长与全株长的比值相同,均较二倍体增加25.58%;入侵地四倍体和六倍体的根长与全株长的比值分别较二倍体增加7.69%和9.62%,且六倍体的根长与全株长的比值显著大于四倍体。并且,入侵地加拿大一枝黄花二倍体和六倍体的根长与全株长的比值显著大于原产地,分别较原产地增加20.93%和5.56%;其四倍体的根长与全株长的比值略大于原产地。
在子叶期,原产地和入侵地加拿大一枝黄花四倍体和六倍体的茎直径显著大于二倍体,其中,原产地四倍体和六倍体的茎直径分别较二倍体增加37.93%和34.48%,且四倍体和六倍体间差异不显著;入侵地四倍体和六倍体的茎直径分别较二倍体增加28.13%和34.38%,且四倍体和六倍体间差异不显著。并且,入侵地加拿大一枝黄花二倍体和六倍体的茎直径显著大于原产地,分别较原产地增加10.34%和10.26%;其四倍体的茎直径略大于原产地。
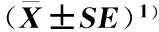
样品2)Sample2)全株长/mmWholeplantlength地上部长度/mmAbove-groundpartlength根长/mmRootlength根长与全株长的比值Ratioofrootlengthtowholeplantlength茎直径/mmStemdiameterNA2x7.28±0.12e4.14±0.06d3.14±0.09e0.43±0.01d0.29±0.00eIN2x11.56±0.26d5.54±0.11c6.02±0.17d0.52±0.01c0.32±0.01dNA4x12.39±0.33c5.61±0.10c6.79±0.26c0.54±0.01bc0.40±0.01bcIN4x13.93±0.39b6.28±0.17ab7.65±0.13b0.56±0.01ab0.41±0.01abNA6x12.83±0.24c5.92±0.22bc6.91±0.25c0.54±0.01bc0.39±0.01cIN6x14.83±0.35a6.35±0.16a8.48±0.18a0.57±0.01a0.43±0.01a
1)同列中不同小写字母表示差异显著(P<0.05) Different lowercases in the same column indicate the significant difference (P<0.05).
2)NA2x: 原产地二倍体 Diploid from native range; IN2x: 入侵地二倍体 Diploid from introduced range; NA4x: 原产地四倍体 Tetraploid from native range; IN4x: 入侵地四倍体 Tetraploid from introduced range; NA6x: 原产地六倍体 Hexaploid from native range; IN6x: 入侵地六倍体 Hexaploid from introduced range.
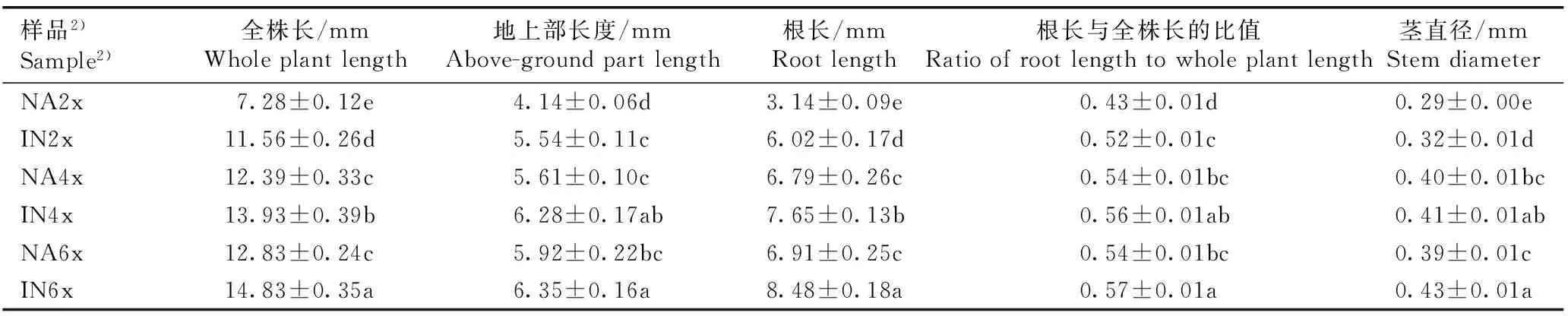
2.1.2子叶形态特征的比较观察和测量结果(图版Ⅰ-B和表2)表明:在子叶期,原产地和入侵地加拿大一枝黄花四倍体和六倍体的子叶长度显著大于二倍体,其中,原产地四倍体和六倍体的子叶长度分别较二倍体增加52.82%和58.46%,且四倍体和六倍体间差异不显著;入侵地四倍体和六倍体的子叶长度分别较二倍体增加20.00%和20.38%,且四倍体和六倍体间差异不显著。并且,入侵地加拿大一枝黄花二倍体和四倍体的子叶长度显著大于原产地,分别较原产地增加35.90%和6.71%;其六倍体的子叶长度略大于原产地。
在子叶期,原产地和入侵地加拿大一枝黄花四倍体和六倍体的子叶宽度显著大于二倍体,其中,原产地四倍体和六倍体的子叶宽度分别较二倍体增加57.45%和58.51%,且四倍体和六倍体间差异不显著;入侵地四倍体和六倍体的子叶宽度分别较二倍体增加26.06%和26.76%,且四倍体和六倍体间差异不显著。并且,入侵地加拿大一枝黄花二倍体、四倍体和六倍体的子叶宽度显著大于原产地,分别较原产地增加51.06%、20.95%和20.81%。
在子叶期,原产地和入侵地加拿大一枝黄花四倍体和六倍体的子叶长度与宽度的比值与二倍体间差异不显著;并且,入侵地加拿大一枝黄花二倍体、四倍体和六倍体的子叶长度与宽度的比值显著小于原产地,分别较原产地减少9.66%、11.82%和13.04%。
在子叶期,原产地和入侵地加拿大一枝黄花四倍体和六倍体的子叶面积显著大于二倍体,其中,原产地四倍体和六倍体的子叶面积分别较二倍体增加133.97%和140.67%,且四倍体和六倍体间差异不显著;入侵地四倍体和六倍体的子叶面积分别较二倍体增加34.48%和42.83%,且四倍体和六倍体间差异不显著。并且,入侵地加拿大一枝黄花二倍体、四倍体和六倍体的子叶面积显著大于原产地,分别较原产地增加123.44%、28.43%和32.60%。
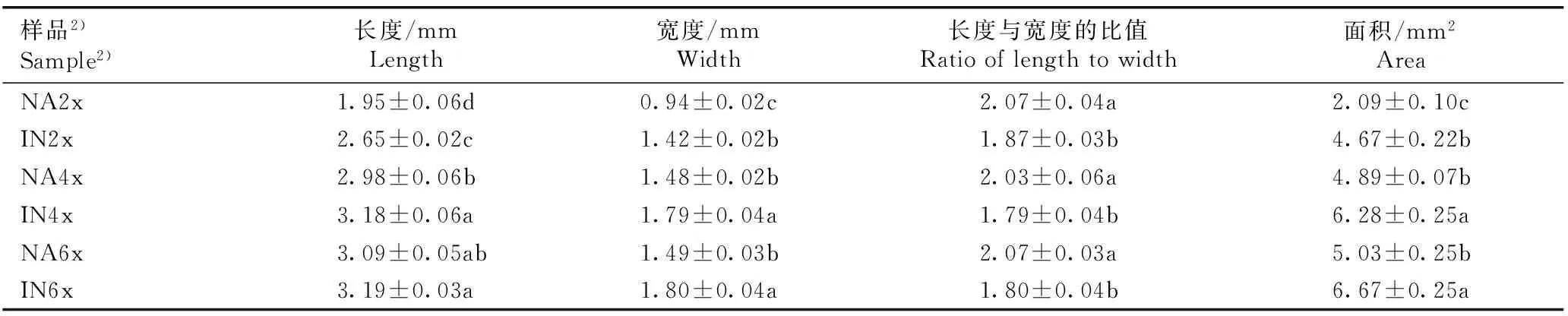
样品2)Sample2)长度/mmLength宽度/mmWidth长度与宽度的比值Ratiooflengthtowidth面积/mm2AreaNA2x1.95±0.06d0.94±0.02c2.07±0.04a2.09±0.10cIN2x2.65±0.02c1.42±0.02b1.87±0.03b4.67±0.22bNA4x2.98±0.06b1.48±0.02b2.03±0.06a4.89±0.07bIN4x3.18±0.06a1.79±0.04a1.79±0.04b6.28±0.25aNA6x3.09±0.05ab1.49±0.03b2.07±0.03a5.03±0.25bIN6x3.19±0.03a1.80±0.04a1.80±0.04b6.67±0.25a
1)同列中不同小写字母表示差异显著(P<0.05) Different lowercases in the same column indicate the significant difference (P<0.05).
2)NA2x: 原产地二倍体 Diploid from native range; IN2x: 入侵地二倍体 Diploid from introduced range; NA4x: 原产地四倍体 Tetraploid from native range; IN4x: 入侵地四倍体 Tetraploid from introduced range; NA6x: 原产地六倍体 Hexaploid from native range; IN6x: 入侵地六倍体 Hexaploid from introduced range.
2.2 真叶期第一真叶形态特征的比较
2.2.1第一真叶期的比较观察和测量结果(图版Ⅰ-C和表3)表明:在第一真叶期,原产地和入侵地加拿大一枝黄花四倍体和六倍体的第一真叶长度显著大于二倍体,其中,原产地四倍体和六倍体的第一真叶长度分别较二倍体增加22.49%和39.95%,且六倍体的第一真叶长度显著大于四倍体;入侵地四倍体和六倍体的第一真叶长度分别较二倍体增加24.94%和32.71%,且六倍体的第一真叶长度显著大于四倍体。并且,入侵地加拿大一枝黄花二倍体、四倍体和六倍体的第一真叶长度显著大于原产地,分别较原产地增加12.43%、14.69%和6.62%。
在第一真叶期,原产地和入侵地加拿大一枝黄花四倍体和六倍体的第一真叶宽度显著大于二倍体,其中,原产地四倍体和六倍体的第一真叶宽度分别较二倍体增加11.48%和17.62%,且六倍体的第一真叶宽度显著大于四倍体;入侵地四倍体和六倍体的第一真叶宽度相同,均较二倍体增加10.34%。并且,入侵地加拿大一枝黄花二倍体和四倍体的第一真叶宽度显著大于原产地,分别较原产地增加6.97%和5.88%;其六倍体的第一真叶宽度略大于原产地。
在第一真叶期,原产地和入侵地加拿大一枝黄花四倍体和六倍体的第一真叶长度与宽度的比值显著大于二倍体,其中,原产地四倍体和六倍体的第一真叶长度与宽度的比值分别较二倍体增加2.99%和11.38%,且六倍体的第一真叶长度与宽度的比值显著大于四倍体;入侵地四倍体和六倍体的第一真叶长度与宽度的比值分别较二倍体增加12.88%和20.25%,且六倍体的第一真叶长度与宽度的比值显著大于四倍体。并且,入侵地加拿大一枝黄花二倍体的第一真叶长度与宽度的比值略小于原产地;其四倍体和六倍体的第一真叶长度与宽度的比值显著大于原产地,分别较原产地增加6.98%和5.38%。
在第一真叶期,原产地和入侵地加拿大一枝黄花四倍体和六倍体的第一真叶面积显著大于二倍体,其中,原产地四倍体和六倍体的第一真叶面积分别较二倍体增加55.90%和87.43%,且六倍体的第一真叶面积显著大于四倍体;入侵地四倍体和六倍体的第一真叶面积分别较二倍体增加37.88%和46.01%,且六倍体的第一真叶面积显著大于四倍体。并且,入侵地加拿大一枝黄花二倍体、四倍体和六倍体的第一真叶面积显著大于原产地,分别较原产地增加37.27%、21.40%和6.94%。
2.2.2第二真叶期的比较观察和测量结果(图版Ⅰ-D和表3)表明:在第二真叶期,原产地和入侵地加拿大一枝黄花四倍体和六倍体的第一真叶长度基本上显著大于二倍体,仅原产地四倍体的第一真叶长度略大于二倍体,其中,原产地四倍体和六倍体的第一真叶长度分别较二倍体增加36.15%和36.38%,且四倍体和六倍体间差异不显著;入侵地四倍体和六倍体的第一真叶长度分别较二倍体增加5.23%和5.05%,且四倍体和六倍体间差异不显著。并且,入侵地加拿大一枝黄花二倍体、四倍体和六倍体的第一真叶长度显著大于原产地,分别较原产地增加34.74%、4.13%和3.79%。
在第二真叶期,原产地加拿大一枝黄花四倍体和六倍体的第一真叶宽度显著大于二倍体,分别较二倍体增加26.32%和28.07%,且四倍体和六倍体间差异不显著;入侵地四倍体和六倍体的第一真叶宽度略大于二倍体,分别较二倍体增加4.15%和4.84%,且四倍体和六倍体间差异不显著。并且,入侵地加拿大一枝黄花二倍体的第一真叶宽度显著大于原产地,较原产地增加26.75%;其四倍体和六倍体的第一真叶宽度略大于原产地。
在第二真叶期,原产地四倍体和六倍体的第一真叶长度与宽度的比值显著大于二倍体,分别较二倍体增加14.12%和12.43%,且四倍体和六倍体间差异不显著;入侵地四倍体和六倍体的第一真叶长度与宽度的比值略大于二倍体,且四倍体和六倍体间差异不显著。并且,入侵地加拿大一枝黄花二倍体的第一真叶长度与宽度的比值显著大于原产地,较原产地增加13.56%;其四倍体和六倍体的第一真叶长度与宽度的比值与原产地相同。
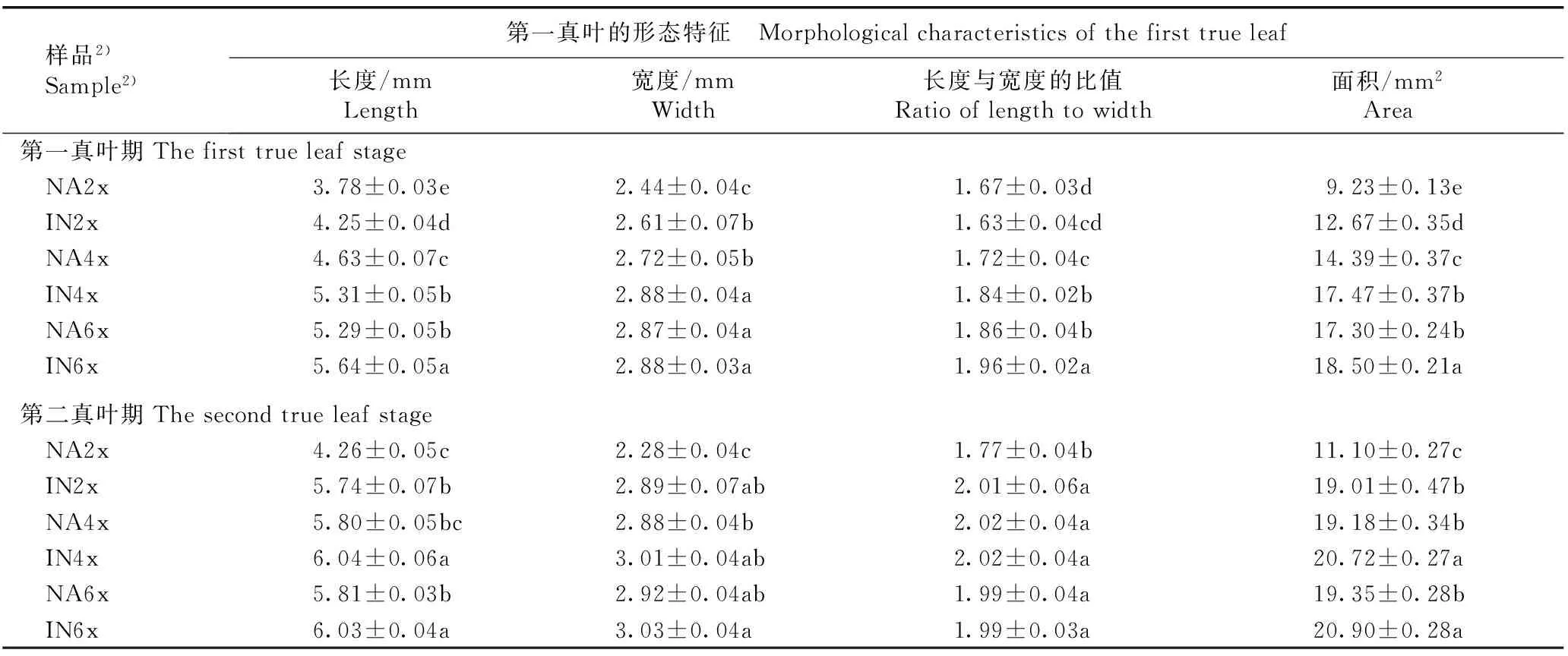
样品2) Sample2)第一真叶的形态特征 Morphologicalcharacteristicsofthefirsttrueleaf长度/mmLength宽度/mmWidth长度与宽度的比值Ratiooflengthtowidth面积/mm2Area第一真叶期Thefirsttrueleafstage NA2x3.78±0.03e2.44±0.04c1.67±0.03d9.23±0.13e IN2x4.25±0.04d2.61±0.07b1.63±0.04cd12.67±0.35d NA4x4.63±0.07c2.72±0.05b1.72±0.04c14.39±0.37c IN4x5.31±0.05b2.88±0.04a1.84±0.02b17.47±0.37b NA6x5.29±0.05b2.87±0.04a1.86±0.04b17.30±0.24b IN6x5.64±0.05a2.88±0.03a1.96±0.02a18.50±0.21a第二真叶期Thesecondtrueleafstage NA2x4.26±0.05c2.28±0.04c1.77±0.04b11.10±0.27c IN2x5.74±0.07b2.89±0.07ab2.01±0.06a19.01±0.47b NA4x5.80±0.05bc2.88±0.04b2.02±0.04a19.18±0.34b IN4x6.04±0.06a3.01±0.04ab2.02±0.04a20.72±0.27a NA6x5.81±0.03b2.92±0.04ab1.99±0.04a19.35±0.28b IN6x6.03±0.04a3.03±0.04a1.99±0.03a20.90±0.28a
1)同列中不同小写字母表示差异显著(P<0.05) Different lowercases in the same column indicate the significant difference (P<0.05).
2)NA2x: 原产地二倍体 Diploid from native range; IN2x: 入侵地二倍体 Diploid from introduced range; NA4x: 原产地四倍体 Tetraploid from native range; IN4x: 入侵地四倍体 Tetraploid from introduced range; NA6x: 原产地六倍体 Hexaploid from native range; IN6x: 入侵地六倍体 Hexaploid from introduced range.
在第二真叶期,原产地和入侵地加拿大一枝黄花四倍体和六倍体的第一真叶面积显著大于二倍体,其中,原产地四倍体和六倍体的第一真叶面积分别较二倍体增加72.79%和74.32%,且四倍体和六倍体间差异不显著;入侵地四倍体和六倍体的第一真叶面积分别较二倍体增加9.00%和9.94%,且四倍体和六倍体间差异不显著。并且,入侵地加拿大一枝黄花二倍体、四倍体和六倍体的第一真叶面积显著大于原产地,分别较原产地增加71.26%、8.03%和8.01%。
3 讨论和结论
与二倍体相比,多倍体植株一般具有器官巨型性特点,如茎粗壮,叶片、种子、花和果实增大等[23-24]。植物的生长和形态性状(尤其是与光能捕获、生物量制造及资源利用有关的性状)是评价植物资源有效利用的重要指标[25]。总体来看,在子叶期,原产地和入侵地加拿大一枝黄花四倍体和六倍体的全株长、地上部长度、根长、根长与全株长的比值、茎直径以及子叶的长度、宽度和面积显著高于二倍体;并且,入侵地各细胞型的上述指标显著高于原产地。进入真叶期(包括第一和第二真叶期)后,原产地和入侵地加拿大一枝黄花四倍体和六倍体的第一真叶长度、宽度、长度与宽度的比值和面积也显著高于二倍体;并且,入侵地各细胞型的上述指标总体上显著高于原产地。另外,入侵地各细胞型子叶期的子叶长度与宽度的比值显著低于原产地,说明原产地加拿大一枝黄花的子叶较入侵地更为狭长。
高生长速率可加快适宜条件下植物的建群速度,促使其在短时间内占据更大的资源空间,从而较土著种更具竞争优势,实现成功入侵。多倍体营养组织的生长速率较高,具有明显的竞争优势[26-31]。入侵植物多倍体常具有较高的遗传多样性[32],利于形成入侵能力较强的新基因型植物[33],并可产生丰富的后代表型,从而为自然选择提供丰富的材料。表型可塑性是指同一基因型个体为适应环境变化而产生的不同表型[34-35],是生物界普遍存在的现象之一,在入侵植物快速适应环境的过程中起到关键作用,可提高植物的竞争力[36]。相关研究结果表明:植物多倍体化过程可产生大量的不符合经典遗传学规律的可遗传的表观遗传变异[37],这些变异虽然没有改变基因组的DNA序列,但是能够影响不同性状的基因表达,并可随细胞的有丝分裂或减数分裂遗传给后代[38]。Senchina等[39]认为,表观遗传变异可引起多倍体植物产生一些重要变化,包括基因加倍后表达的多样性、遗传学和细胞学上的二倍化以及基因组间的相互协调等。加拿大一枝黄花多倍体拥有多组染色体,基因重组数多,杂合性强,促使其在入侵初期能够快速适应新环境,并快速生长、繁殖和进化,容易突破奠基者效应实现成功定居,这对加拿大一枝黄花成功入侵具有重要意义。
综上所述,入侵地加拿大一枝黄花多倍体的幼苗具有明显的生长优势,狭长的子叶是其二倍体的特征,可以作为早期鉴别的依据。
参考文献:
[1]李扬汉. 中国杂草志[M]. 北京: 中国农业出版社, 1998: 372.
[2]WEBER E. Morphological variation of the introduced perennialSolidagocanadensisL.sensulato(Asteraceae) in Europe[J]. Botanical Journal of the Linnean Society, 1997, 123: 197-210.
[3]VAN KLEUNEN M, RAMPONI G, SCHMID B. Effects of herbivory simulated by clipping and jasmonic acid onSolidagocanadensis[J]. Basic and Applied Ecology, 2004, 5: 173-181.
[4]WEBER E. Biological flora of Central Europe:SolidagoaltissimaL.[J]. Flora, 2000, 195: 123-134.
[5]MEYER A, SCHMID B. Experimental demography of rhizome populations of establishing clones ofSolidagoaltissima[J]. Journal of Ecology, 1999, 87: 42-54.
[6]董梅, 陆建忠, 张文驹, 等. 加拿大一枝黄花——一种正在迅速扩张的外来入侵植物[J]. 植物分类学报, 2006, 44(1): 72-85.
[7]吴海荣, 强胜. 加拿大一枝黄花生物生态学特性及防治[J]. 杂草科学, 2005(1): 52-56.
[8]LU J Z, WENG E S, WU X W, et al. Potential distribution ofSolidagocanadensisin China[J]. Acta Phytotaxonomica Sinica, 2007, 45(5): 670-674.
[9]李振宇, 解焱. 中国外来入侵种[M]. 北京: 中国林业出版社, 2002: 170.
[10]JIN L, GU Y, XIAO M, et al. The history ofSolidagocanadensisinvasion and the development of its mycorrhizal associations in newly-reclaimed land[J]. Functional Plant Biology, 2004, 31: 979-986.
[11]黄华, 郭水良. 外来入侵植物加拿大一枝黄花繁殖生物学研究[J]. 生态学报, 2005, 25(11): 2795-2803.
[12]CHEN G Q, ZHANG C B, MA L, et al. Biotic homogenization caused by the invasion ofSolidagocanadensisin China[J]. Journal of Integrative Agriculture, 2013, 12(5): 835-845.
[13]杨如意, 昝树婷, 唐建军, 等. 加拿大一枝黄花的入侵机理研究进展[J]. 生态学报, 2011, 31(4): 1185-1194.
[14]MELVILLE M R, MORTON J K. A biosystematic study of theSolidagocanadensis(Compositae) complex. Ⅰ. The Ontario populations[J]. Canadian Journal of Botany, 1982, 60: 976-997.
[15]蔡华, 王荣富. 加拿大一枝黄花的核型分析及B染色体初报[J]. 激光生物学报, 2006, 15(3): 245-248.
[16]彭闪江, 黄忠良, 彭少麟, 等. 植物天然更新过程中种子和幼苗死亡的影响因素[J]. 广西植物, 2004, 24(2): 113-121.
[17]张佳宁, 刘坤. 植物调节萌发时间和萌发地点的机制[J]. 草业学报, 2014, 23(1): 328-338.
[18]CRAINE J M, LEE W G, BOND W J, et al. Environment constraints on a global relationship among leaf and root traits of grasses[J]. Ecology, 2005, 86(1): 12-19.
[19]张业. 油松天然林林内主要植物种的叶功能性状研究[D]. 北京: 北京林业大学生态学院, 2012: 10-13.
[20]WESTOBY M. A leaf-height-seed (LHS) plant ecology strategy scheme[J]. Plant and Soil, 1998, 199: 213-227.
[21]张红香. 种子发芽生态研究[D]. 长春: 东北师范大学城市与环境科学学院, 2008: 13-14.
[22]郑伟. 植物幼苗生长对策研究[D]. 长春: 东北师范大学城市与环境科学学院, 2011: 17-19.
[23]BRETAGNOLLE F, THOMPSON J D, LUMARET R. The influence of seed size variation on seed germination and seedling vigour in diploid and tetraploidDactylisglomerataL.[J]. Annals of Botany, 1995, 76: 607-615.
[24]GARBUTT K, BAZZAZ F A. Leaf demography, flower production and biomass of diploid and tetraploid populations ofPhloxdrummondiiHook. on a soil moisture gradient[J]. New Phytologist, 1983, 93(1): 129-141.
[25]NAVAS M L, MOREAU-RICHARD J. Can traits predict the competitive response of herbaceous Mediterranean species?[J]. Acta Oecologica, 2005, 27(2): 107-114.
[26]李君, 强胜. 多倍化是杂草起源与演化的驱动力[J]. 南京农业大学学报, 2012, 35(5): 64-76.
[27]GRIME J P. Vegetation classification by reference to strategies[J]. Nature, 1974, 250: 26-31.
[28]WESTOBY M, FALSTER D S, MOLES A T, et al. Plant ecological strategies: some leading dimensions of variation between species[J]. Annual Review of Ecology and Systematics, 2002, 33: 125-159.
[29]BLUMENTHAL D M, HUFBAUER R A. Increased plant size in exotic populations: a common-garden test with 14 invasive species[J]. Ecology, 2008, 88(11): 2758-2765.
[30]VAN KLEUNEN M, WEBER E, FISCHER M. A meta-analysis of trait differences between invasive and non-invasive plant species[J]. Ecology Letters, 2009, 13: 235-245.
[31]RAPP R A, WENDEL J F. Epigenetics and plant evolution[J]. New Phytologist, 2005, 168(1): 81-91.
[32]ANDREAKIS N, KOOISTRA W H C F, PROCACCINI G. High genetic diversity and connectivity in the polyploid invasive seaweedAsparagopsistaxiformis(Bonnemaisoniales) in the Mediterranean, explored with microsatellite alleles and multilocus genotypes[J]. Molecular Ecology, 2009, 18(2): 212-226.
[33]MARRS R A, SFORZA R, HUFBAUER R A. When invasion increases population genetic structure:a study withCentaureadiffusa[J]. Biological Invasions, 2008, 10(4): 561-572.
[34]马俊改, 石福臣. 养分条件对互花米草表型可塑性的影响[J]. 生态学杂志, 2011, 30(3): 459-463.
[35]杜乐山. 加拿大一枝黄花的表型可塑性与适应性研究[D]. 北京: 北京林业大学自然保护区学院, 2014: 10-14.
[36]刘海燕. 不同地理种群加拿大一枝黄花的表型可塑性及对遮光和盐胁迫的响应[D]. 临汾: 山西师范大学生命科学学院, 2015: 10-14.
[37]WENDEL J F. Genome evolution in polyploids[J]. Plant Molecular Biology, 2000, 42(1): 225-249.
[38]GUIMIL S, DUNAND C. Patterning ofArabidopsisepidermal cells: epigenetic factors regulate the complex epidermal cell fate pathway[J]. Trends in Plant Science, 2006, 11(12): 601-609.
[39]SENCHINA D S, ALVAREZ I, CRONN R C. Rate variation among nuclear genes and the age of polyploidy inGossypium[J]. Molecular Biology and Evolution, 2003, 20(4): 633-643.
