不同水位梯度下拉鲁湿地典型湿草甸植物群落特征
2018-04-08杜红霞
王 丽, 杨 涛, 杜红霞, 张 军
(1. 西安建筑科技大学环境与市政工程学院 西北水资源与环境生态教育部重点实验室 陕西省环境工程重点实验室, 陕西 西安 710055; 2. 中国电建集团 西北勘测设计研究院有限公司 环保与安评设计院, 陕西 西安 710065)
青藏高原地理环境特殊且地处江河源头,其水热条件处于生物极限水平,生态系统极度脆弱[1],该地区大面积的高原湿地具有广泛的生态学意义[2]。高寒湿草甸湿地的水分条件良好,植被覆盖度高,生长茂盛,是良好的放牧场所或刈割草场;由于其土壤湿度及养分适中,当地居民在定居和垦殖过程中,常将其用作建设用地或开垦为农田。但不合理的利用方式导致该地区湿草甸湿地植被退化,面积剧减[3],其中水文条件退化是其退化的关键环节[4-5]。
水文条件是湿地各生态过程运行的关键驱动因子[6-7]。水文条件的变化直接或间接导致植物调节个体生长,改变种群优势,进而引起群落结构及景观格局的动态变化。水位是描述水文特征的最简单直观的指标,其对湿地植被的影响涉及生理、个体、种群、群落及景观的各研究尺度[7]。植被是湿地健康水平的直接表征。不同水文条件下植被的生长及动态变化一直是湿地研究的热点。植物多样性水平能反映植物群落结构及功能特征,而生物量对群落结构的形成有重要影响,是评价群落结构和功能的重要指标[6,8],因此,研究植物多样性和生物量对准确认识植物群落结构及动态变化具有重要意义。
华扁穗草(BlysmussinocompressusTang et F. T. Wang)型高寒湿草甸的植被生长茂盛,草本层发达,耐放牧和践踏,是西藏地区分布较广的一种湿草甸类型。该湿草甸草群低矮茂密,多数地区旺季呈郁闭状态,多以华扁穗草占优势,伴生植物种类较少,主要有矮生嵩草〔Kobresiahumilis(C. A. Mey. ex Trautv.) Sergiev〕、鹅绒委陵菜(PotentillaanserineLinn.)、西藏报春(PrimulatibeticaWatt)和具槽秆荸荠(EleocharisvalleculosaOhwi)等[9]。本研究在拉鲁湿地国家级自然保护区典型华扁穗草湿草甸分布区内设置观测样地,对不同生长期植物群落进行调查,分析水位梯度对高寒湿草甸湿地植物群落物种多样性及地上部分生物量特征的影响,探讨高寒湿草甸湿地的合理利用方式,以期为高寒湿草甸湿地的保护与恢复提供背景数据及理论支撑。
1 研究区概况和研究方法
1.1 研究区概况
拉鲁湿地国家级自然保护区(北纬29°39′46″~29°41′05″、东经91°03′48″~91°06′51″)位于拉萨市区西北角,平均海拔3 645 m,为海拔最高、世界稀有的城市湿地。保护区所在的拉萨河谷属藏南高原温带半干旱季风气候,气温低,雨季、旱季分明。年平均降水量439.3 mm,年平均气温7.6 ℃[10]。保护区内地形简单,地势东北高、西南低,补给水源主要为流沙河、北干渠和地下潜水。土壤主要为泥炭沼泽土和泥炭土,植被主要为泥炭沼泽型草甸。从20世纪60年代中期开始,受挖沟修渠、开荒种地以及城市建设等活动的影响,拉鲁湿地呈严重的退化态势,目前保留面积6.2 km2。另外,由于1990年后修建的横穿拉鲁湿地的中干渠渠底低于湿地地面高程,拉鲁湿地内70%的水流通过该渠流走,导致拉鲁湿地水位下降,加重了拉鲁湿地的退化态势。
1.2 研究方法
采用以空间代替时间的方法,在拉鲁湿地典型湿草甸湿地分布区内,选取处于不同微地貌区域的湿草甸设置观测样地,调查群落组成及水位情况。根据现场情况,共选取5个水位带,水位(即实验期内的平均水位)分别为-15.0、-5.0、0.0、2.5和6.5 cm,其中水位-15.0和-5.0 cm分别为地面以下15.0和5.0 cm。采用直尺测量水位,其中水位面位于地面以下的水位带先用土钻打洞,再用直尺测量水位。
以2010年群落中优势种华扁穗草的生长初期(5月初)和生长旺盛期(8月中旬)表示群落的生长初期和生长旺盛期,在每一水位带随机设置5个面积0.5 m×0.5 m的调查样方,记录群落总盖度和高度以及群落组成物种的种名、盖度、高度和频度。根据调查数据计算群落内物种重要值、物种丰富度指数、Simpson多样性指数、Shannon-Wiener多样性指数和Pielou均匀度指数。调查完毕后贴地收割各样方植物地上部分,带回实验室后先称量其鲜质量,再于60 ℃烘干至恒质量,并称量其干质量,然后计算地上部分生物量和地上部分干鲜比。
1.3 数据处理
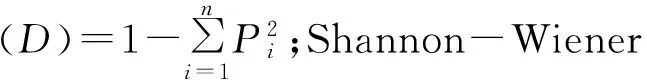
采用SPSS 11.5统计分析软件进行数据分析、单因素方差分析和差异显著性分析(Duncan’s新复极差法),采用Origin 8.5软件制图。
2 结果和分析
2.1 不同水位梯度下拉鲁湿地湿草甸植物群落的物种多样性特征分析
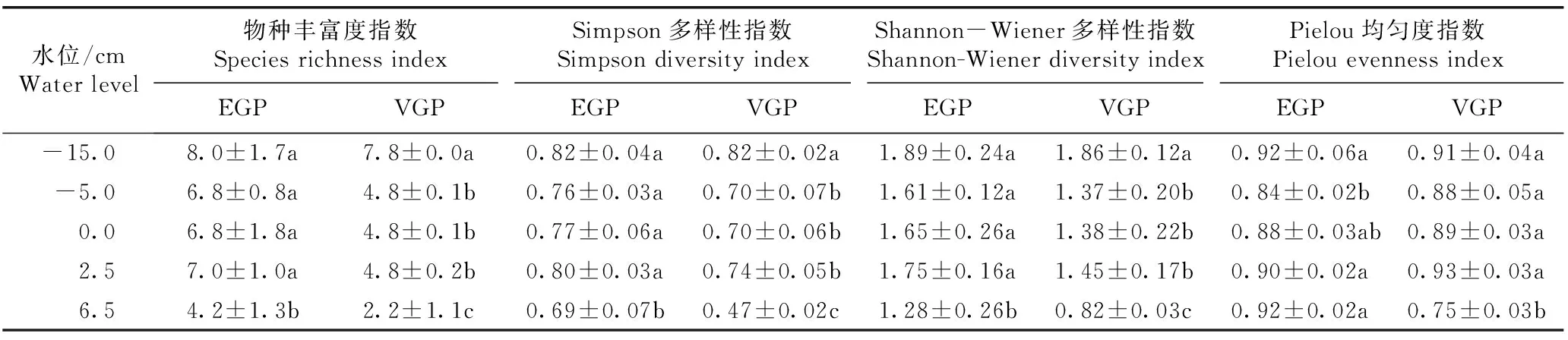
不同水位梯度下拉鲁湿地湿草甸植物群落的物种多样性特征见表1。
2.1.1对物种丰富度指数的影响由表1可见:在生长初期和生长旺盛期,水位对拉鲁湿地湿草甸植物群落物种丰富度指数均有显著影响(P<0.05),物种丰富度指数总体上随着水位的升高而降低。生长初期,该群落的物种丰富度指数在6.5 cm水位下为4.2,显著低于其他水位;在-15.0 cm水位下最高,为8.0。在生长旺盛期,在-5.0、0.0和2.5 cm水位下物种丰富度指数均为4.8,3个水位间物种丰富度指数差异不显著;在-15.0 cm水位下物种丰富度指数为7.8,显著高于其他水位;在6.5 cm水位下物种丰富度指数为2.2,显著低于其他水位。同一水位下,生长旺盛期群落的物种丰富度指数均较生长初期有所降低,但仅在-5.0、2.5和6.5 cm水位下存在显著差异。
2.1.2对Simpson多样性指数的影响由表1还可见:在生长初期和生长旺盛期,水位对拉鲁湿地湿草甸植物群落的Simpson多样性指数均有显著影响。在2个生长期,Simpson多样性指数均在-15.0 cm水位下最高,在6.5 cm水位下最低,总体上随着水位的升高而降低。生长旺盛期群落的Simpson多样性指数的变幅明显大于生长初期。在-5.0~2.5 cm水位范围内,2个生长期群落的Simpson多样性指数在不同水位间均无显著差异。同一水位下,生长旺盛期群落的Simpson多样性指数总体上低于生长初期,但仅在6.5 cm水位下存在显著差异。

水位/cmWaterlevel物种丰富度指数SpeciesrichnessindexSimpson多样性指数SimpsondiversityindexShannon-Wiener多样性指数Shannon-WienerdiversityindexPielou均匀度指数PielouevennessindexEGPVGPEGPVGPEGPVGPEGPVGP-15.08.0±1.7a7.8±0.0a0.82±0.04a0.82±0.02a1.89±0.24a1.86±0.12a0.92±0.06a0.91±0.04a-5.06.8±0.8a4.8±0.1b0.76±0.03a0.70±0.07b1.61±0.12a1.37±0.20b0.84±0.02b0.88±0.05a0.06.8±1.8a4.8±0.1b0.77±0.06a0.70±0.06b1.65±0.26a1.38±0.22b0.88±0.03ab0.89±0.03a2.57.0±1.0a4.8±0.2b0.80±0.03a0.74±0.05b1.75±0.16a1.45±0.17b0.90±0.02a0.93±0.03a6.54.2±1.3b2.2±1.1c0.69±0.07b0.47±0.02c1.28±0.26b0.82±0.03c0.92±0.02a0.75±0.03b
1)EGP: 生长初期Early growth period; VGP: 生长旺盛期Vigorous growth period. 同列中不同的小写字母表示差异显著(P<0.05) Different lowercases in the same column indicate the significant difference (P<0.05).
2.1.3对Shannon-Wiener多样性指数的影响由表1还可见:在生长初期和生长旺盛期,水位对拉鲁湿地湿草甸植物群落的Shannon-Wiener多样性指数均有显著影响,Shannon-Wiener多样性指数总体上随着水位的升高而降低。在2个生长期,群落的Shannon-Wiener多样性指数均在-15.0 cm水位下最高,在6.5 cm水位下最低,且生长旺盛期群落的Shannon-Wiener多样性指数的变幅明显大于生长初期。在-5.0~2.5 cm水位范围内,2个生长期群落的Shannon-Wiener多样性指数在不同水位间均无显著差异。同一水位下,生长旺盛期群落的Shannon-Wiener多样性指数均显著低于生长初期。
2.1.4对Pielou均匀度指数的影响由表1还可见:在生长初期,水位对拉鲁湿地湿草甸植物群落的Pielou均匀度指数有显著影响。其中,Pielou均匀度指数在-15.0和6.5 cm水位下最高,在-5.0 cm水位下最低,且随着水位的升高呈先降低后升高的趋势。在生长旺盛期,群落的Pielou均匀度指数在-15.0、-5.0、0.0和2.5 cm水位下无显著差异,但在-5.0~2.5 cm水位范围内,Pielou均匀度指数呈升高趋势,在2.5 cm水位下达到最高;在6.5 cm水位下,Pielou均匀度指数最低,且显著低于其他水位。在-5.0~2.5 cm水位范围内,2个生长期群落的Pielou均匀度指数在不同水位间总体上无显著差异。在-5.0、0.0和2.5 cm水位下,生长旺盛期群落的Pielou均匀度指数略高于生长初期,而在-15.0和6.5 cm水位下则低于生长初期,但仅在6.5 cm水位下Pielou均匀度指数在2个生长期间存在显著差异。
2.2 不同水位梯度下拉鲁湿地湿草甸植物群落地上部分生物量分析
2.2.1对地上部分鲜质量和干质量的影响不同水位梯度下拉鲁湿地湿草甸植物群落的地上部分生物量(地上部分鲜质量和干质量)见图1。由图1可见:在生长初期和生长旺盛期,水位对拉鲁湿地湿草甸植物群落地上部分鲜质量和干质量均有显著影响(P<0.05)。随着水位的升高,2个生长期地上部分鲜质量和干质量均呈“升高—降低—升高”的趋势,且不同水位下地上部分鲜质量的变幅更大。
在生长初期,地上部分鲜质量在6.5 cm水位下最大,显著高于其他水位,在-5.0 cm水位下次之,在-15.0和0.0 cm水位下较小。在生长旺盛期,地上部分鲜质量在-5.0 cm水位下最大,显著高于其他水位,在6.5 cm水位下次之,在0.0和2.5 cm水位下较小。
与地上部分鲜质量相似,在生长初期,地上部分干质量在6.5 cm水位下最大,在-5.0 cm水位下次之,在-15.0和0.0 cm水位下较小。在生长旺盛期,地上部分干质量在-5.0 cm水位下最大,显著高于其他水位,而其他水位间的地上部分干质量无显著差异,在0.0和2.5 cm水位下较小。
2.2.2对地上部分干鲜比的影响不同水位梯度下拉鲁湿地湿草甸植物群落的地上部分干鲜比见图2。由图2可见:在生长初期和生长旺盛期,水位对拉鲁湿地湿草甸植物群落地上部分干鲜比均有显著影响。在2个生长期,地上部分干鲜比总体上随着水位的升高而降低。同一水位下,生长旺盛期的地上部分干鲜比均显著低于生长初期。
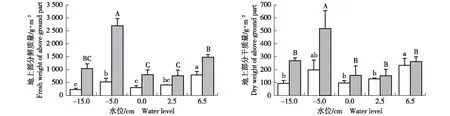
□: 生长初期Early growth period; : 生长旺盛期Vigorous growth period. 不同的小写或大写字母分别表示生长初期或生长旺盛期同一指标在不同水位间差异显著(P<0.05) Different lowercases or capitals indicate the significant difference (P<0.05) of the same index among different water levels in early growth period or vigorous growth period, respectively.图1 不同水位梯度下拉鲁湿地湿草甸植物群落的地上部分生物量Fig. 1 Biomass of above-ground part of wet meadow plant community in Lalu wetland under different water level gradients

□: 生长初期Early growth period; : 生长旺盛期Vigorous growth period. 不同的小写或大写字母分别表示生长初期或生长旺盛期在不同水位间差异显著(P<0.05) Different lowercases or capitals indicate the significant difference (P<0.05) among different water levels in early growth period or vigorous growth period, respectively.图2 不同水位梯度下拉鲁湿地湿草甸植物群落的地上部分干鲜比Fig. 2 Ratio of dry weight to fresh weight of above-ground part of wet meadow plant community in Lalu wetland under different water level gradients
3 讨论和结论
3.1 水位梯度对高寒湿草甸植物群落的影响
水位梯度可通过直接或间接影响土壤环境来作用于湿地植物的繁殖、生长及竞争能力,进而调控植物分布格局及群落结构[7,12]。有关低海拔地区的大量研究结果表明:随着水位的升高,湿地植物群落物种多样性总体上呈降低的趋势[7,13]。而有关高海拔地区的研究较少,对巴音布鲁克[5]及若尔盖[14]高寒沼泽植物群落物种多样性的研究结果显示:植物群落的物种多样性水平随着水位的升高而降低。本研究中,拉鲁湿地湿草甸各水位带的水位在生长季内处于波动状态,其中-15.0和6.5 cm水位带的水位面分别在地下和地上,而-5.0、0.0和2.5 cm水位带的水位面在地面上下波动,属于干湿交替的环境。与其他高原湿地植物群落物种多样性研究中的无积水、季节性积水和长期淹水的划分进行比较[6],本研究中群落物种多样性呈相似的随着水位升高而降低的趋势,水位梯度对拉鲁湿地湿草甸植物群落的物种多样性有显著影响。
水位梯度及水位的波动对湿地植物生物量的影响已有多方面验证[15]。本研究中,拉鲁湿地湿草甸植物群落的地上部分干鲜比随着水位的升高而降低,说明水位梯度对群落地上部分生物量的影响与植物的水分利用策略有关。在适宜的水位范围内,随着水位的升高,植物生物量呈升高的趋势,但超过一定范围后生物量则总体呈降低的趋势[7,16]。本研究中,群落地上部分鲜质量和干质量在-15.0~-5.0 cm和0.0~6.5 cm 2个水位范围内均随着水位的升高而升高。在生长初期,地上部分鲜质量和干质量均在6.5 cm水位下最大,这可能与生长初期竞争作用不明显,生物量主要受群落优势种华扁穗草个体生长优势的影响有关。而在生长旺盛期,地上部分鲜质量和干质量均在-5.0 cm水位下最大,这主要是多物种充分利用资源的结果。群落的物种多样性与生物量的关系并不是绝对的,刘庆[8]认为,青海湖北岸植物群落生物量与物种丰富度的关系受环境因子影响。本研究中,群落的物种多样性均在-15.0 cm水位下最高,而群落的地上部分生物量则在该水位下较低。
拉鲁湿地国家级自然保护区水文环境退化最重要的原因为保护区南侧中干渠的排水作用[17]。韩大勇等[6]研究了若尔盖高原退化沼泽群落物种多样性沿排水梯度的变化,认为该群落的物种丰富度指数和Shannon-Wiener多样性指数随着水位的升高而降低。该研究涉及的水位变化范围基本包含本研究中涉及的5个水位。虽然本研究中群落的物种多样性指标总体上随着水位的升高而降低,但在-5.0~2.5 cm水位范围内,4个物种多样性指标均呈明显的升高趋势,这可能与本研究主要针对湿草甸植被有关。不同的植物种群均有其特定的生态分布范围[6]。湿草甸湿地是一种处于交替带的湿地类型。在适宜的水位范围内,水位越高越利于湿草甸植物生长,物种多样性也越高,而超出适宜的水位范围后,水位升高时仅耐淹物种可存活。
3.2 植物群落特征对于高寒湿草甸湿地保护与恢复的意义
群落物种多样性和生物量均是表征植物群落结构和动态的量化特征[8]。物种丰富度指数、Simpson多样性指数、Shannon-Wiener多样性指数及均匀度指数是描述或评价植物群落物种多样性特征的主要指标。Simpson多样性指数对物种均匀度更为敏感,而Shannon-Wiener多样性指数对物种丰富度更为敏感[18-19]。本研究中,拉鲁湿地湿草甸植物群落同样表现为不同水位下群落的Simpson多样性指数与Pielou均匀度指数的变化趋势更相似,而Shannon-Wiener多样性指数与物种丰富度指数的变化趋势更接近。同一水位下,物种丰富度指数和Shannon-Wiener多样性指数在不同生长期间总体上差异显著,对植物群落的变化更为敏感,因此,在高寒湿草甸植被相关的监测或评价工作中,物种丰富度指数和Shannon-Wiener多样性指数对群落结构的变化更灵敏。
水文条件的恢复是湿地恢复的首要措施,在拉鲁湿地国家级自然保护区内也一直在进行相关工作[17]。华扁穗草是该保护区湿草甸植物群落的优势种,随着水位的升高,华扁穗草的重要值总体上呈上升的趋势(另文发表)。根据本研究结果,在适宜的水位范围内,水位的升高有利于高寒湿草甸植物群落物种多样性的恢复,6.5 cm水位下群落的物种多样性显著低于其他水位,但是群落地上部分生物量却较高,这主要与6.5 cm水位下优势种华扁穗草的重要值显著高于低水位有关,而在-15.0 cm水位下虽然群落的物种多样性最高,但优势种华扁穗草的重要值及群落地上部分生物量则较低,且群落中出现了藏蓟〔Cirsiumlanatum(Roxb. ex Willd.) Spreng.〕和车前(PlantagoasiaticaLinn.)等旱生物种。在植被恢复工作中,优势种的恢复对于群落重建有重要影响[13],因此,在湿草甸湿地恢复过程中,水位梯度的设置以及群落的物种多样性和生物量指标在植被健康评价中的运用应该根据恢复工作的阶段目标进行衡量。
本研究根据研究区微地貌差异选取了5个水位带,初步对比分析了生长初期和生长旺盛期不同水位梯度间高寒湿草甸植物群落多样性及地上部分生物量的差异。而水位梯度只是湿地水文特征中最简单的一个指标,水位波动频率及幅度等对群落动态同样具有重要影响[20-21],应进一步研究高寒湿草甸植物对水文条件及其引起的土壤特征差异的响应规律,以更全面地认识高寒湿草甸植被生长的影响因子,并有助于高寒湿草甸湿地生态系统的保护与恢复。
参考文献:
[1]姚檀栋, 朱立平. 青藏高原环境变化对全球变化的响应及其适应对策[J]. 地球科学进展, 2006, 21(5): 459-464.
[2]白军红, 欧阳华, 徐惠风, 等. 青藏高原湿地研究进展[J]. 地理科学进展, 2004, 23(4): 1-9.
[3]琼次仁, 拉琼. 拉萨市拉鲁湿地的初步研究[J]. 西藏大学学报, 2000, 15(4): 40-41.
[4]王丽, 张军, 杜红霞, 等. 火烧及微地貌对高原湿草甸优势种华扁穗草地上部灰分含量及热值的影响[J]. 植物资源与环境学报, 2016, 25(4): 1-7.
[5]王铭, 曹议文, 王升忠, 等. 水位和草丘微地貌对巴音布鲁克高寒沼泽植物群落物种多样性的影响[J]. 湿地科学, 2016, 14(5): 635-640.
[6]韩大勇, 杨永兴, 杨杨. 若尔盖高原退化沼泽群落植物多样性及种间相关性沿排水梯度的变化[J]. 植物生态学报, 2012, 36(5): 411-419.
[7]姚鑫, 杨桂山, 万荣荣, 等. 水位变化对河流、湖泊湿地植被的影响[J]. 湖泊科学, 2014, 26(6): 813-821.
[8]刘庆. 青海湖北岸环境梯度上植物群落的生物量与物种多样性及其相互关系[J]. 西北植物学报, 2000, 20(2): 259-267.
[9]中华人民共和国农业部畜牧兽医司, 全国畜牧兽医总站. 中国草地资源[M]. 北京: 中国科学技术出版社, 1996: 324-325.
[10]巴桑, 普布, 马正学, 等. 西藏拉鲁湿地夏季和秋季浮游藻类群落特征[J]. 湿地科学, 2012, 10(4): 404-416.
[11]张相锋, 马闯, 董世魁, 等. 不同草灌配比对泌桐高速公路护坡植物群落特征的影响[J]. 草业学报, 2009, 18(4): 27-34.
[12]杨娇, 厉恩华, 蔡晓斌, 等. 湿地植物对水位变化的响应研究进展[J]. 湿地科学, 2014, 12(6): 807-813.
[13]WANG L, SONG C, HU J, et al. Responses of regeneration diversity of theCarexlasiocarpacommunity to different water levels[J]. Chinese Geographical Science, 2010, 20: 37-42.
[14]崔丽娟, 马琼芳, 郝云庆, 等. 若尔盖高寒沼泽植物群落与环境因子的关系[J]. 生态环境学报, 2013, 22(11): 1749-1756.
[15]LI S, PEZESHKI S R, GOODWIN S. Effects of soil moisture regimes on photosynthesis and growth in cattail (Typhalatifolia)[J]. Acta Oecologica, 2004, 25: 17-22.
[16]徐金英, 陈海梅, 王晓龙. 水深对湿地植物生长和繁殖影响研究进展[J]. 湿地科学, 2016, 14(5): 725-732.
[17]摆万奇, 土艳丽, 李建川, 等. 筑坝在湿地恢复中的作用: 以拉萨市拉鲁湿地为例[J]. 资源科学, 2010, 32(9): 1666-1671.
[18]许晴, 张放, 许中旗, 等. Simpson指数和Shannon-Wiener指数若干特征的分析及“稀释效应”[J]. 草业科学, 2011, 28(4): 527-531.
[19]马克平, 黄建辉, 于顺利, 等. 北京东灵山地区植物群落多样性的研究: Ⅱ丰富度、均匀度和物种多样性指数[J]. 生态学报, 1995, 15(3): 268-277.
[20]王丽, 宋长春, 胡金明, 等. 毛苔草地下构件对不同水文情势和水文经历的生长响应[J]. 应用生态学报, 2008, 19(10): 2194-2200.
[21]ALVAREZ-COBELAS M, CIRUJANO S. Multilevel responses of emergent vegetation to environmental factors in a semiarid floodplain[J]. Aquatic Botany, 2007, 87: 49-60.
