干旱胁迫对桃叶杜鹃菌根苗生长的影响
2018-03-27熊贤荣龙海燕欧阳嘉晖
熊贤荣 欧 静 龙海燕 欧阳嘉晖 熊 丹
(贵州大学林学院,贵州 贵阳 550025)
干旱是限制植物生长的重要非生物因素之一,我国的干旱和半干旱地区约占全国土地面积的1/2,其限制了园林植物的分布及其在景观中的应用范围[1]。桃叶杜鹃 (Rhododendronannae) 是杜鹃花科杜鹃花属常绿灌木,为高山常绿杜鹃代表种,根系为浅根系,不能伸入土层深处,不耐干旱,干旱制约其引种栽培及园林应用[2]。菌根类植物,相较于非菌根类植物来说,能促进水分的吸收,具有更强的耐旱能力,能够在更为干旱的环境下正常生长和发育。菌根化造林技术越来越受到人们重视,它可以增强苗木的存活率、提高植物的质量[3],因此加强菌根化苗木的研究对园林植物栽培工作具有十分重要的作用。本试验材料以1 a生桃叶杜鹃幼苗,通过接种杜鹃花类菌根 (ERM),进行干旱胁迫处理,通过对桃叶杜鹃幼苗苗高、地径、生物量、根冠比的生长指标的测定,探讨接种ERM真菌对桃叶杜鹃幼苗抗旱性的影响。
1 材料与方法
1.1 试验材料
供试土壤为百里杜鹃土壤腐殖质和石英砂以3∶1的比例混匀后在高压灭菌锅灭菌20 min,灭菌后测得其土壤理化性质为pH 4.77,有机质44.15 g/kg,全氮1.56 g/kg,全磷0.161 g/kg,有效磷7.04 mg/kg,碱解氮268.55 mg/kg,速效钾205.91 mg/kg。供试的2个试验菌株为前期从野生桃叶杜鹃根系分离并筛选出的综合促生能力和侵染率较强的菌株[2,4]: TY29 (AB158314:Trametesochracea),TY35 (AB378554:Pochoniabulbillosa)。
1.2 试验方法
于2015年12月播种后放入人工气候箱内恒温培养 (白天、夜间均为25 ℃,湿度为90%、光照12 h)。2016年4月将桃叶杜鹃幼苗移出人工气候培养箱,移栽至花盆中,每盆移植1株,栽植塑料花盆规格为90 cm × 60 cm × 80 cm,每盆装灭菌土20 g。2016年4月12日,每株幼苗根部各施入菌液5 mL,CK苗浇5 mL不含菌液的PDA液体培养基,在贵州大学林学院苗圃大棚内进行培育。同一个处理随机摆放,各处理之间相互隔离,进行常规育苗方法统一管理。
设4个胁迫梯度,即正常水分 (WW,土壤含水率80%~90%) 作为干旱胁迫的对照,轻度干旱 (MW,土壤含水率65%~75%),中度干旱 (MS,土壤含水率50%~60%)和重度干旱 (SS,土壤含水率35%~45%)。2016年8月,选取生长一致的幼苗进行胁迫处理,每天用称量法保持每盆恒定,每个处理15盆,每盆1株,各处理重复3次。
1.3 测定方法
干旱胁迫开始后,每隔10天对幼苗苗高进行1次测量,共测3次,每个处理随机抽取10株幼苗用卷尺 (精确到0.1 cm) 测量,3次重复,即前期苗高 (8月10日)、中期苗高 (8月20)、后期苗高 (8月30日)。胁迫30天后,每个处理取出5株幼苗侵入水中,小心冲洗,取出完整的根系,采用多功能根系分析测量仪 (MRS-9600TFU2L) 进行根系扫描获取图像,用万深根系分析软件 (Microtek ScanWizard EZ) 对根系图片进行各项指标分析,包括叶面积、总根长、表面积、体积、平均直径和根尖数。植株收获时菌根侵染率按Phillips等[5]方法测定,并按公式 (1)~(4) 计算各指标。
菌根侵染率=(菌根侵染的根段数/检测
的根段总数) × 100%
(1)
根冠比=根干质量/地上部干质量
(2)
菌根依赖性=(接种植株平均干质量/对照
植株平均干质量) × 100%
(3)
菌根贡献率=(接种植株平均干质量-不接
种植株平均干质量)/ 接种植
株平均干质量 × 100%
(4)
1.4 数据分析
采用Execl 2010记录数据并绘图, SPSS 20.0软件对数据进行单因素方差分析和Duncan多重比较分析。
2 结果与分析
2.1 菌根的侵染率、依赖性及贡献率
不同菌株接种桃叶杜鹃幼苗的菌根侵染率、菌根依赖性及菌根贡献率见表1。

表1 不同菌株接种桃叶杜鹃幼苗的菌根侵染率、菌根依赖性及菌根贡献率Table 1 Mycorrhizal colonsation rate、mycorrhizal dependency and mycorrhizal contribution rate of different strains inoculation of R.annae seedlings
注:不同小写字母表示差异显著;不同大写字母表示差异极显著。
由表1可知,不同ERM菌株接种桃叶杜鹃幼苗,其菌根侵染率不同,其中TY35的侵染率最高为45%,TY29的侵染率为27.5%,未接种的CK处理幼苗根系侵染率为0,极显著低于 (P< 0.01) 接种处理幼苗根系的侵染率。不同菌株对宿主植物根系表现出的亲和力具有差异性,即反映出了宿主植物对不同菌根真菌的依赖性不同。接种苗的菌根依赖性均大于100%,TY35达到147.89%;接种幼苗的菌根贡献率为13.65%~32.38%。
2.2 干旱胁迫对根系形态的影响
由图1可知,在任何一种水分胁迫下,接种处理的桃叶杜鹃幼苗总根长均显著大于未接种处理 (P< 0.05),对照组总根长为57.82~131.46 cm,接种组为112.48~212.93 cm。正常水分条件下,TY29和TY35的总根长较CK分别增加了25.7%和61.9%,说明接种ERM真菌能够增加桃叶杜鹃幼苗根系总根长,促进根系生长。轻度胁迫下,各处理总根长均有所增加,且菌根苗的增幅大于CK。随着胁迫加剧,干旱抑制了根系的伸长与生长,总根长随着土壤含水量的降低而降低。各处理根系长度最大值均出现在轻度胁迫条件下,最小值出现在重度胁迫下,且TY35的根系长度大于TY29。这说明适度的干旱胁迫能够刺激桃叶杜鹃根系的生长,以伸入土壤更深处吸收水分,提高其适应干旱的能力,但当干旱超过一定限度时,则会严重抑制根系的伸长生长,根系总长度下降。
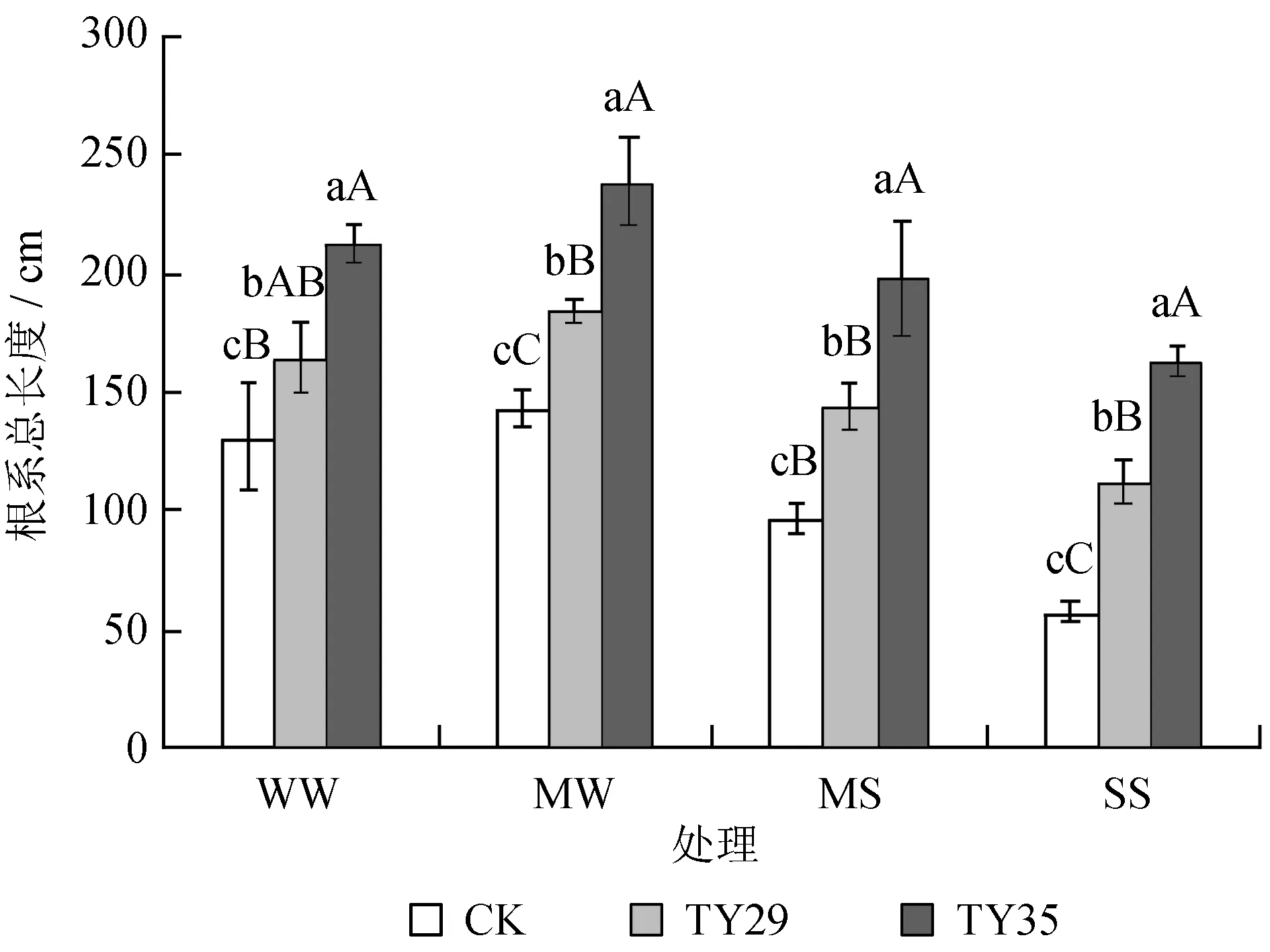
图1干旱胁迫下不同ERM真菌对桃叶杜鹃幼苗总根长的影响
Fig.1 Effects of inoculation with different ericoid mycorrhizal strains on total length ofR.annaeroots under drought stress
由图2可知,接种ERM真菌显著提高了桃叶杜鹃幼苗根系的表面积 (P< 0.05),相同水分胁迫下,接种幼苗的根系表面积均大于未接种处理。在轻度、中度和重度胁迫时,接种幼苗的根系表面积与CK之间差异达极显著水平 (P< 0.01)。正常水分下,各处理根系表面积为13.81~23.29 cm2,其中TY35的根系表面积最大,CK最小。各处理苗的表面积在轻度胁迫时均有所增加,且菌根苗的增幅大于对照苗,而后随着干旱胁迫的加剧呈下降趋势。在重度干旱胁迫下,各处理根系表面积显著降低 (P< 0.05),达到最小值,CK、TY29和TY35根系表面积依次为5.19、11.04和17.21 cm2,与正常水分相比,接种处理与非接种处理之间差异达极显著性水平 (P< 0.01)。
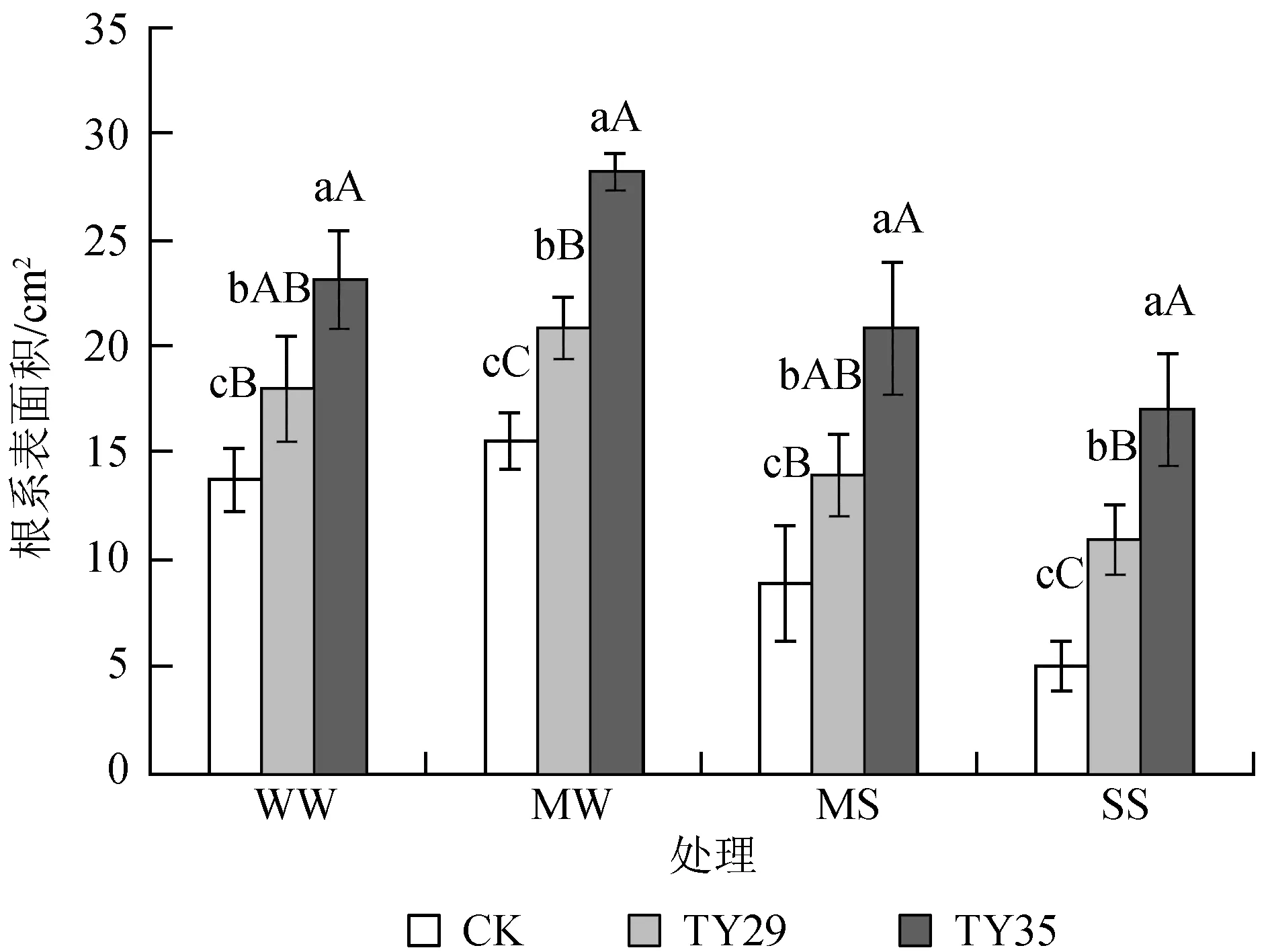
图2干旱胁迫下不同ERM真菌对桃叶杜鹃幼苗根系表面积的影响
Fig.2 Effects of inoculation with different ericoid mycorrhizal strains on surface area ofR.annaeroots under drought stress
由图3可知,各处理根系体积随土壤干旱程度加重而逐渐减小。在正常水分条件下,根系体积最大,CK、TY29和TY35的根系体积依次为0.27、0.32和0.44 cm3,TY35与TY29和CK之间的差异均达到显著性水平 (P< 0.05),TY29与CK之间差异虽未达显著性水平,但接种TY29的幼苗根系体积仍大于CK根系体积。可见,不同的菌株对桃叶杜鹃幼苗根系的促进作用具有一定的差异性,以TY35对根系的促生效应最为明显。在中度与重度胁迫下,根系体积降幅明显,但接种处理幼苗的根系体积仍显著大于未接种处理 (P< 0.05);重度干旱时,各处理苗的根系体积均达到最小值,接种处理之间无显著性差异,可能是由于土壤过度干旱抑制了真菌孢子的侵染,导致菌根效应降低,幼苗根系体积下降。
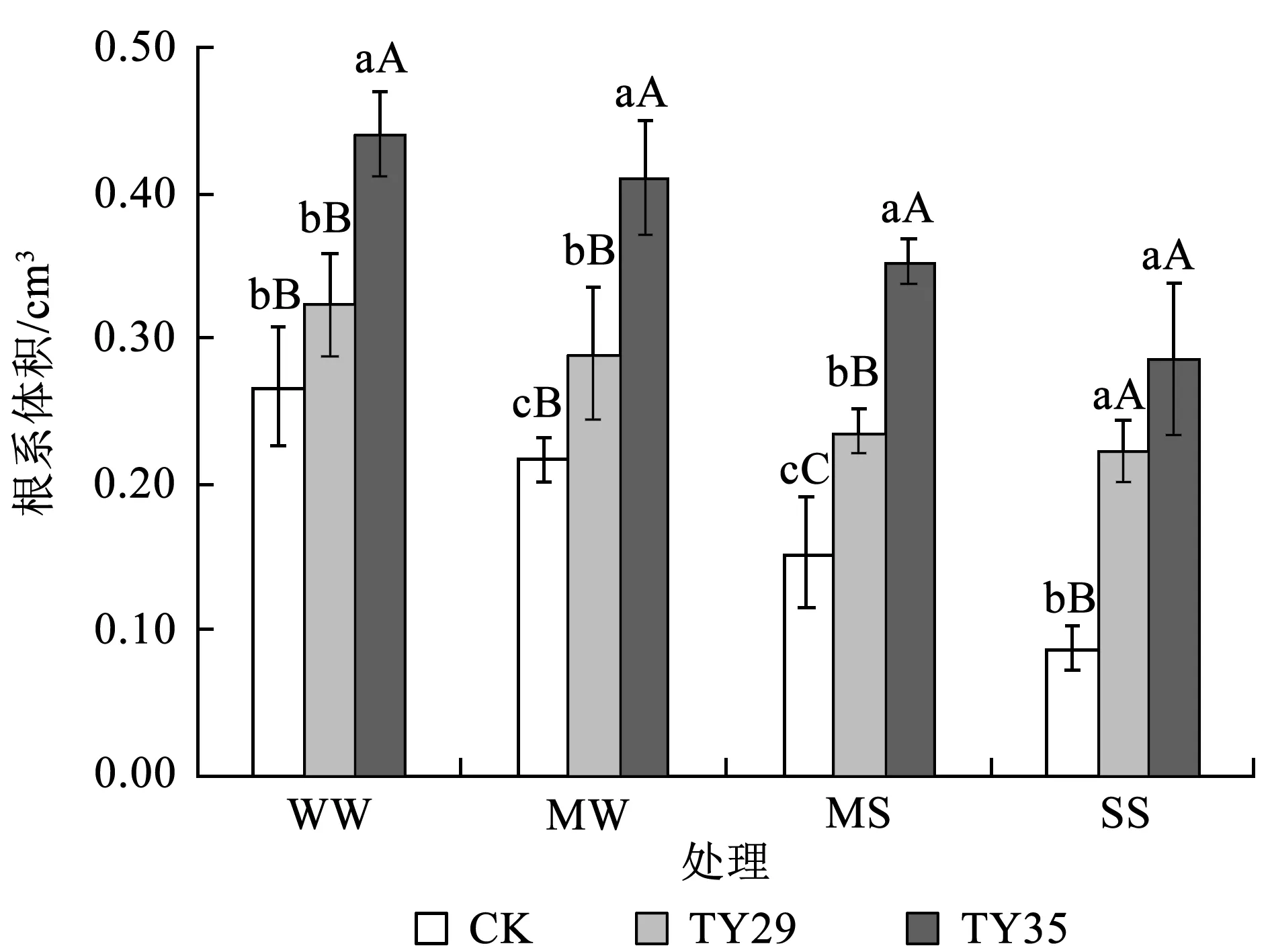
图3干旱胁迫下不同ERM真菌对桃叶杜鹃幼苗根系体积的影响
Fig.3 Effects of inoculation with different ericoid mycorrhizal strains on roots volume ofR.annaeunder drought stress
由图4可知,接种处理降低了幼苗根系的平均直径,在各水分胁迫条件下,CK根系平均直径均为最大,而TY35的根系直径均为最小。正常水分下,CK与接种处理之间差异显著 (P< 0.05),而接种处理之间差异未达显著水平。随着干旱胁迫加剧,各处理平均直径均有不同程度的增加,中度胁迫时达到最大值,CK、TY29和TY35的根系平均直径依次为0.43、0.37和0.32 mm;重度干旱胁迫时,CK、TY29和TY35的根系平均直径又逐渐下降,接种处理与CK之间无显著性差异。
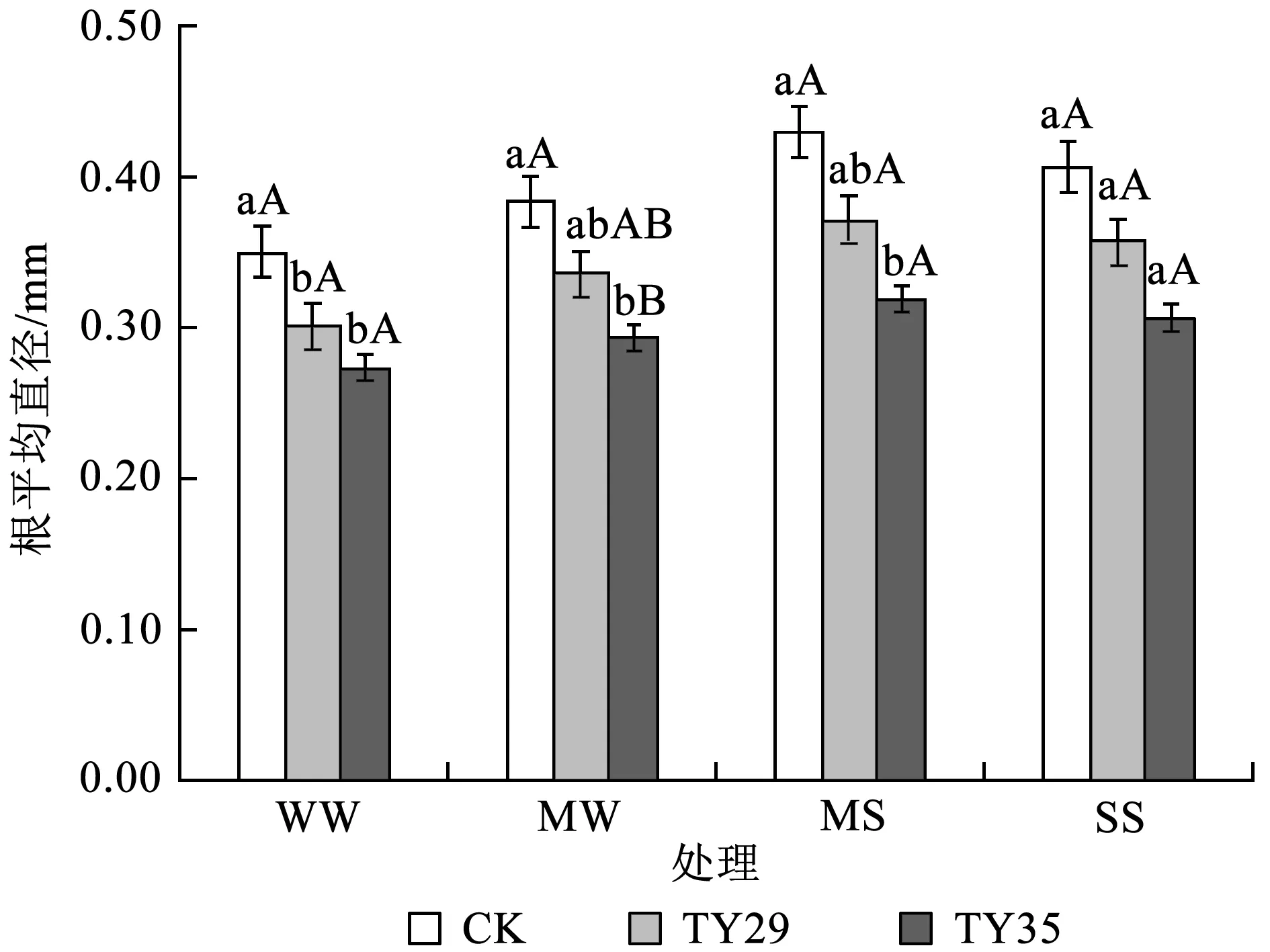
图4干旱胁迫下不同ERM真菌对桃叶杜鹃幼苗根平均直径的影响
Fig.4 Effects of inoculation with different ericoid mycorrhizal strains on average diameter ofR.annaeroots under drought stress
由图5可知,根尖数量的变化趋势大致与总根长和根系表面积呈一致性,都随着干旱胁迫的加剧而下降。在相同的水分胁迫条件下,接种菌根的根系根尖数量均显著大于未接种根系 (P< 0.05)。同一菌根不同水分胁迫下,TY29与CK根尖数量的最大值都出现在正常水分条件下,后随着土壤水分的减少而逐渐降低,而TY35则呈现出先增后减的趋势,在轻度胁迫时达到最大值,重度胁迫时达到最小。
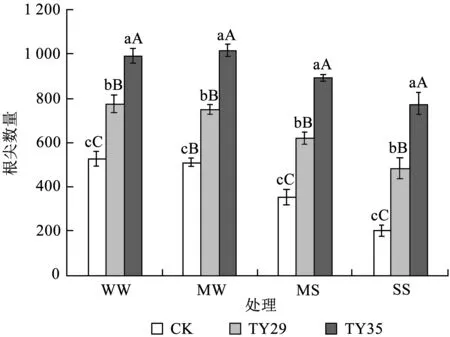
图5干旱胁迫下不同ERM真菌对桃叶杜鹃幼苗根尖数量的影响
Fig.5 Effects of inoculation with different ericoid mycorrhizal strains on number of root tips ofR.annaeroots under drought stress
2.3 干旱胁迫对苗高的影响
干旱胁迫下接种ERM菌株对桃叶杜鹃幼苗苗高的影响见表2。由表2可知,接菌和水分胁迫处理对桃叶杜鹃的苗高均产生了显著的影响。同一时期不同水分胁迫下,菌根苗的苗高均大于CK,且差异显著 (P< 0.05)。正常水分条件下,各时期均以TY35的苗高最大,在前期TY35和TY29,较CK分别增加了24.2%和17.7%,且差异显著 (P< 0.05),但接菌苗之间差异不显著。中期时,TY35和TY29较CK分别增加了24.7%和14.4%,后期则较CK增加了22.7%和11.4%,且各处理之间差异显著 (P< 0.05)。不同时期苗高均有不同程度的增加,且后期的增幅明显小于前期和中期,说明随着胁迫时间的延长,各处理的生长都受到了一定的影响。
在不同水分胁迫条件下,菌根苗的生长均呈现出先升高后降低的趋势,在轻度胁迫时达到最大,在重度胁迫时达到最低值,而CK则呈现逐步下降的趋势。在胁迫后期,各处理苗高均随着胁迫程度的加剧逐渐降低,但菌根苗的苗高下降幅度均小于CK,TY35的下降幅度最小。这说明在一定程度的干旱下,接菌能够促进桃叶杜鹃幼苗的生长从而提高其抵御干旱的能力,且不同的菌根苗会表现出一定的差异性,其中以接种TY35的幼苗效果较好。

表2 干旱胁迫下接种ERM菌株对桃叶杜鹃幼苗苗高的影响Table 2 Effects of inoculation with different ericoid mycorrhizal strains on height of R.annae seedling under drought stress
注:不同小写字母表示差异显著;不同大写字母表示差异极显著。
2.4 干旱胁迫对叶面积的影响
从图6可知,同一水分胁迫下,接菌苗的叶面积均大于CK苗,且差异显著 (P< 0.05)。正常水分时,TY35和TY29叶面积较CK分别增加了66.4%和31.2%。在轻度和中度胁迫下,也均以接种TY35的苗木叶面积最大,且与其他菌株处理呈极显著差异 (P< 0.01)。这说明了接菌对桃叶杜鹃的生长和叶片的萌发起到了较好的促生作用,不同的ERM菌株促生效果不尽相同。
随着干旱胁迫的加剧,桃叶杜鹃幼苗的总叶面积逐渐降低,重度干旱胁迫时,各处理叶面积达到最小值,与正常水分相比,CK下降了41.3%,TY29和TY35分别下降了44.5%和38.0%,TY35的降幅最小,与其他处理苗呈极显著差异 (P< 0.01),TY29的降幅较大,与CK相比无显著性差异,但其叶面积仍然大于CK苗。这一结果说明,干旱胁迫对植物的叶面积产生了一定的影响,相较菌根苗而言,CK对干旱胁迫更为敏感,而菌根苗在干旱胁迫时仍能保持较高的叶面积以保证光合作用的进行,从而提高幼苗的抗旱能力。

图6干旱胁迫下不同ERM真菌对桃叶杜鹃叶片叶面积的影响
Fig.6 Effects of inoculation with different ericoid mycorrhizal strains on leaf area ofR.annaeleaves under drought stress
2.5 干旱胁迫对生物量与根冠比的影响
水分胁迫下ERM菌株对桃叶杜鹃幼苗生物量和根冠比的影响见表3。由表3可知,接种苗的地上、地下部分干质量与总生物量均较CK提高,在同一水分条件下,均以TY35菌根苗的生物量最大,其次是TY29。这说明接种菌株能够促进幼苗对营养物质的吸收,从而增加其干物质的积累。在正常水分与轻度水分胁迫下,接种苗TY35的总生物量与TY29和CK之间差异显著 (P< 0.05),而接菌苗TY29和CK之间差异未达显著水平,但其生物量仍然高于CK。中度与重度干旱胁迫时,CK地下部分干质量显著低于接种处理 (P< 0.05)。说明在干旱胁迫条件下,接种菌株对植物地下部分的生长促进作用更加明显,使幼苗根系能够有效的从土壤中获取营养物质及水分。随着胁迫程度的加剧,各处理总生物量都随之下降,正常水分时,各处理生物量均为最大值,在重度胁迫时达到最小值,但TY35与TY29的总生物量仍分别高出CK的总生物量33.9%和82.4%,TY35与CK间差异显著 (P< 0.05)。
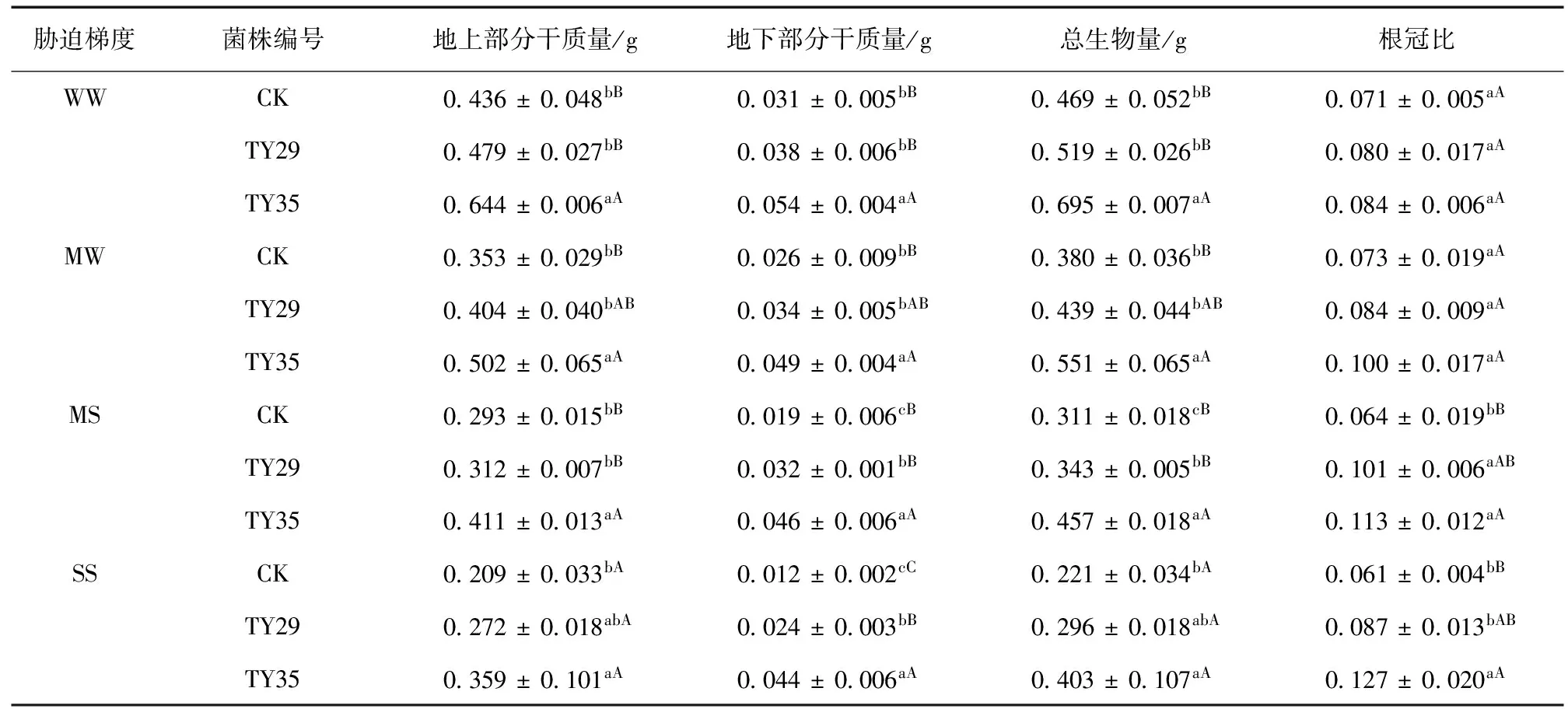
表3 水分胁迫下ERM菌株对桃叶杜鹃幼苗生物量和根冠比的影响Table 3 Effects of inoculation with different ericoid mycorrhizal strains on plant biomass and shoot ratio of R.annae seedling under water stress
注:不同小写字母表示差异显著;不同大写字母表示差异极显著。
同时,桃叶杜鹃幼苗的根冠比也受到了干旱胁迫和菌株的双重影响,相同水分条件下,接菌苗的根冠比均高于CK,且在中度胁迫时差异显著 (P< 0.05),TY35与TY29较CK的根冠比分别高出了57.8%和75.7%。同一菌株不同水分胁迫下,接菌苗的根冠比随着土壤含水量的降低表现出升高的趋势,TY35的增幅大于TY29,而CK的根冠比呈现出先增后减的趋势,在轻度胁迫时有所上升,其后逐渐下降,这可能是由于幼苗接种后能够依赖其庞大的菌丝网,扩大其吸收水分的面积,同时促进根系的生长,进而提高幼苗抵御干旱的能力,而过度的干旱胁迫却严重阻碍了对照苗根系的生长,抑制其苗木生长。
3 结论与讨论
不同ERM菌株对桃叶杜鹃幼苗菌根侵染率、依赖性和贡献率具有差异性。本试验中CK、TY29和TY35菌株对桃叶杜鹃的幼苗侵染率分别为0%、27.5%和45.0%,1年生幼苗的侵染总体水平不高,这可能与ERM真菌选择、侵染时间和定居时间有关[6]。各菌根苗依赖性不同,TY35的菌根依赖性以及菌根贡献率都高于TY29,说明了不同的ERM菌株对桃叶杜鹃幼苗根系的亲和力具有差异性。综合各菌株处理对桃叶杜鹃幼苗生长的促生效应来看,菌株侵染率的高低与促生作用成正相关关系。
菌根真菌能够通过改变宿主植物根系形态,提高植物对土壤水分和矿质元素的吸收,进而促进植物的生长和抗逆性[7]。本试验中,各水分胁迫下,桃叶杜鹃菌根苗根的总长度、表面积与根尖数量均显著高于CK (P< 0.05),菌根苗根体积高于CK,除正常水分外,差异达到显著性水平 (P< 0.05)。与何跃军等[8]对香樟幼苗,张中峰等[8]对青冈栎幼苗的研究结果相似,表明接种ERM真菌改变了桃叶杜鹃根系形态,促进幼苗根系生长,以提高幼苗抗旱性。在轻度胁迫时,桃叶杜鹃各处理根系总根长、表面积均较其他水分胁迫处理高,且菌根苗显著高于CK (P< 0.05),表明适度的干旱能够刺激植物根系的生长,通过增强植物根系的吸水能力,适应干旱环境。随着土壤水分不断减少,对照苗的根冠比先增后减,菌根苗的根冠比逐渐上升,表明干旱胁迫下,接种ERM菌株能够通过优化桃叶杜鹃幼苗生物量的配置格局,促使植物将更多的生物量分配到根系中吸收水分[9],以缓解植物缺水状况。本研究中,任何一种水分胁迫下,菌根苗根系平均直径均小于CK,根尖数量显著大于CK (P< 0.05)。说明接种菌根真菌诱导菌根化植物根系变得更细小,使植物根系更易从土壤中吸收水分和养分[10]。桃叶杜鹃接种后根系平均直径降低、根尖数量显著增加,这些都将有利于幼苗根系增强对水分及养分的吸收作用,提高幼苗抗旱性。
有研究表明,接种ERM菌株能显著提高桃叶杜鹃幼苗的苗高、地径、地上与地下部分生物量及总生物量[11]。本研究表明,在干旱胁迫下,接种ERM菌株显著提高了幼苗的苗高、叶面积 (P< 0.05),增加了幼苗地上部分与地下部分的生物量积累。随着胁迫加剧,CK苗高、叶面积逐渐降低,菌根苗高在轻度胁迫时有所升高,而后随之下降,菌根苗叶面积下降幅度均小于CK,且TY35的降幅最小。可能是因为ERM菌株通过与根系形成的庞大菌丝网,扩大了根系的吸收面积和范围,增加其对矿质营养元素以及水分的吸收,从而减轻幼苗受干旱胁迫的程度,改善幼苗的生长状况。
综上所述,ERM能和桃叶杜鹃形成良好的共生关系,对桃叶杜鹃的生长起到一定的促进作用。在水分胁迫下,ERM真菌能通过影响桃叶杜鹃幼苗根系形态,扩大根系吸收面积,来改善幼苗的水分及营养状况,促进桃叶杜鹃幼苗的生长,提高桃叶杜鹃幼苗适应干旱的能力。不同的菌株促生效果不同,综合幼苗的生长形态指标来看,以接种TY35对幼苗生长的促生效果较好。菌根能够积极对水分胁迫做出响应,使宿主植物具有更强的干旱耐受性,因此菌根技术运用具有重大的意义,但目前国内关于菌根植物在干旱胁迫下的响应多处在个体生理水平上,还缺乏对抗旱分子机制方面系统深入的研究。
[1] 陈立明, 尹艳豹. 干旱区园林植物抗旱机制研究进展[J]. 安徽农业科学, 2015, 43(4): 73-76.
[2] 谌端玉, 欧静, 王丽娟, 等. 干旱胁迫对接种ERM真菌桃叶杜鹃幼苗叶绿素含量及荧光参数的影响[J]. 南方农业学报, 2016, 47(7): 1164-1170.
[3] 刘现刚, 郭素娟. 菌根化技术及其应用研究进展[J]. 林业实用技术, 2011(2): 3-5.
[4] 欧静, 韦小丽, 何跃军, 等. 接种ERM真菌对桃叶杜鹃幼苗的促生效应及生理生化影响[J]. 林业科学, 2013, 49(7): 48-56.
[5] Phillips J M, Hayman D S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection[J]. Transactions of the British Mycological Society, 1970, 55(1): 158-161.
[6] 欧静, 刘仁阳, 陈训, 等. 菌根真菌对桃叶杜鹃种子萌发及幼苗移植的影响[J]. 南方农业学报, 2013, 44(2): 293-298.
[7] 张中峰, 张金池, 黄玉清, 等. 水分胁迫和接种菌根真菌对青冈栎根系形态的影响[J]. 生态学杂志, 2015, 34(5): 1198-1204.
[8] 何跃军, 钟章成. 水分胁迫和接种丛枝菌根对香樟幼苗根系形态特征的影响[J]. 西南大学学报 (自然科学版), 2012, 34(4): 33-39.
[9] Stewart J D, Lieffers V J. Preconditioning effects of nitrogen relative addition rate and drought[J]. Canadian Journal of Forest Research, 1993, 23(23): 1663-1671.
[10] Wurzburger N, Higgins B P, Hendrick R L. Ericorrhizal root fungi and their multicopper oxidases from a temperate forest shrub[J]. Ecology and Evolution, 2012, 2(1): 65-79.
[11] 尹丽娟. 云锦杜鹃根系内生真菌营养特性及其接种效应分析[D]. 福州: 福建农林大学, 2008.
