河北省森林草原区主要草本植物功能群适宜分布预测
2018-03-26范顺祥郑建伟魏士凯黄选瑞张志东
范顺祥,郑建伟,魏士凯,黄选瑞,张志东
(河北省林木种质资源与森林保护重点实验室,河北农业大学林学院,河北 保定071000)
物种分布与环境因子之间的关系一直是生态学研究的焦点[1]。植物功能群作为对环境变化具有相似响应的物种组合[2],是联系生态系统结构和功能的桥梁,有助于理解复杂的生态系统功能[3]。基于光合作用途径划分的C3和C4草本植物功能群,表现出了对光、水、热等环境因子响应的差异性,是较为理想的植物功能群分类途径[4]。研究表明C3和C4功能群在生物地球化学循环方面扮演着重要的角色[5],分析C3和C4功能群的适宜分布范围及限制因素,对于理解植物-环境关系及阐明相关的生态学过程具有重要的作用。然而,目前环境变化对C3和C4物种分布的影响机制仍然存在不确定性[6]。
C3和C4植物分布对环境变化的响应差异因不同的环境因子和地理分区而异。在美国夏威夷C4植物分布与7月最高温度显著相关[7],而在中国东北C4植物分布则主要与年均温、2和9月降水量相关[8];澳大利亚的C3和C4植物的分布与生长季节降水量表现出高度的相关性[9],而在热带安第斯山脉C3和C4植物的分布则主要受年均温的影响[10];在中国禾本科中的C4物种分布比例随着降水量的增加而显著上升,而在印度次大陆却呈相反的变化趋势[11];在阿根廷C4植物主要分布在低海拔地区,高海拔地区则以C3植物为主[12],而在澳大利亚的新几内亚地区C3和C4植物的物种更替发生在海拔1600~3000 m之间[13]。由此可见,不同地区影响C3和C4植物分布的主导环境因子不同,阐明特定地区C3和C4适宜分布格局及与气候和地形因子的关系仍显得十分必要。物种分布模型是预测物种适宜分布的有效途径。在众多的模型中,生态位模型(MaxEnt)具有样本量要求低,预测变量处理简单、运行灵活、预测精度高等诸多优点[14-15]。由于MaxEnt模型表现出的良好性能和较高的精度,其已被广泛应用于濒危物种分布及适宜性评价[16-17]、生物入侵与防治[18-19]、保护区空缺性分析[20-21]等方面。本研究采取MaxEnt模型进行研究区C3和C4草本植物的潜在适宜分布预测。
塞罕坝国家级自然保护区,处于华北典型森林-草原生态交错带,生态系统复杂多样,物种多样性丰富,具有很好的保护价值。但由于长期森林利用、商品林经营,天然植被资源锐减,景观破碎化加剧,天然植物群落和草本植物物种分布格局已发生改变[22]。理解草本植物物种适宜分布格局及影响机制,对于重构该地区受损景观和保护物种多样性很有必要。本研究在对该区184个草本样方进行分层随机调查的基础上,主要探究以下两方面问题:1) C3、C4草本植物的适宜分布格局是否存在显著差异?2) 影响C3、C4草本植物适宜分布的关键环境因子是否相同?通过阐明C3、C4草本植物的潜在适宜分布格局,为优化该地区的景观格局和物种多样性保护提供科学依据。
1 材料与方法
1.1 研究区概况
研究区位于河北省承德市围场满族蒙古族自治县境内的塞罕坝国家级自然保护区(42°22′-42°31′ N, 116°53′-117°31′ E)。本区的地貌组合为高原-波状丘陵-漫滩-接坝山地。气候属于温带半湿润气候与温带半干旱气候的过渡区。冬季漫长、春季错后、夏季不明显,昼夜温差大;年均气温-1.5 ℃,极端最高气温30 ℃,极端最低气温-42.8 ℃,≥5 ℃的年积温1930.7 ℃;年均降水量452.6 mm,蒸发量1230 mm,年均无霜期60 d;植被由森林向草原过渡,主要树种有华北落叶松(Larixprincipis-rupprechtii)、蒙古栎(Quercusmongolica)、白桦(Betulaplatyphylla);灌木种主要有山刺玫(Rosadavurica)、山杏(Armeniacasibirica)、柔毛绣线菊(Spiraeapubescens)等;草本主要由地榆(Sanguisorbaofficinalis)、金莲花(Trolliuschinensis)、披针叶苔草(Carexlanceolata)等物种构成[23]。
1.2 物种分布数据
2014年7-9月在研究区进行了群落学调查。调查是基于空间广布性原则,采取分层随机抽样的方法,按照斑块大小设置不同数量的调查样方,样方间隔至少50 m。在研究区38个天然植被斑块(12个草本、11个灌木、15个天然林)中共调查草本样方184个,样方大小为1 m×1 m。记录草本植物的物种名、多度、盖度、平均高度和最大高度,利用GPS记录每个样方的经纬度坐标[23]。
基于光合碳同化途径的不同,可分为C3途径、C4途径和景天酸代谢(CAM)途径,相应的植物因CO2固定的最初产物不同,分为C3、C4和CAM植物[24]。本研究将调查的草本物种划分为C3、C4和CAM三个功能群。由于研究区CAM草本植物仅含有景天三七(Sedumaizoon)、钝叶瓦松(Orostachysmalacophyllus)和八宝景天(Sedumspectabile)等少数物种,样本点达不到MaxEnt模型所需的最小样本数,因此,只对C3和C4功能群进行适宜分布预测。
1.3 环境因子数据
采用高分辨率气候模型ClimateAP生成研究区气候数据。ClimateAP模型是在WorldClim模型和PRISM模型的基础上,进一步采取双线性插值和动态局部回归的方法将气候数据进行尺度下推的工具[25]。该模型可通过地理坐标和海拔生成亚太地区任意点的无尺度(free scale)月、季、年和未来某时间节点的高精度气候数据集。由于预测精度高,该模型已被广泛应用于特定区域内物种分布预测[26-27]。本研究利用取样点和随机生成点的经纬度和海拔信息,利用ClimateAP模型生成了研究区1981-2012年之间的年度气候历史数据值,包括温度和降水的16个气候因子点数据。利用Arcgis 10.1软件的含障碍样条函数法对这些气候因子点数据进行空间插值,最终生成了研究区的栅格气候因子数据集。
基于研究区等高线、河流、湖泊等矢量数据,采用GIS软件的空间分析模块生成DEM数据,并在此基础上获得了坡度、坡向、海拔及地形位置指数(topographic position index,TPI)等栅格数据[25]。初步共选取20个环境因子(包括4个地形因子和16个气候因子)用于模拟研究区草本植物的潜在分布。
本研究采用的栅格数据,均使用Xian_1980_GK_Zone_20投影坐标系,分辨率10 m×10 m。为减少预测变量的相关性,提高模拟精度和运行效果,对20个环境变量进行了相关性检验,选取了13个相关性较低的变量(|r|<0.80),用于模型运算(表1)。
1.4 适宜分布预测
采用MaxEnt模型(http://www.cs.princeton.edu /~schapire/maxent)预测C3、C4物种的适宜分布。随机选择75%的C3、C4植物分布点数据用于建立模型,而余下的25%分布点数据用于模型验证。在建立模型过程中,采取刀切法[18](Jackknife)和响应曲线分析各环境因子贡献率及对功能群适宜分布的影响。选择第10个百分点的训练存在阈值(10th percentile training presence threshold),进行预测结果的解释和可视化。同时选择交叉验证(crossvalidate)重复运算模型10次,以获得更加准确的验证结果。模型其他参数设为默认值,输出预测栅格结果为以逻辑值形式给出的概率值(0~1)。选择10次模型运行中较好的结果进行下一步适宜区划分。以第10个百分点的训练存在值作为适宜分布阈值[28],低于此阈值的划为不适宜区,高于此阈值的又进一步划分:边缘适宜区(阈值~0.50)、低度适宜区(0.50~0.65)、中度适宜区(0.65~0.80)和高度适宜区(>0.80),通过栅格计算统计各适宜区的面积比例。

表1 构建物种分布模型的环境因子 Table 1 Environmental variables for the species distribution model
采用受试者工作特征曲线(receiver operating characteristic curve, ROC)对模型的精度进行评价。ROC 曲线是以假阳性率(1-特异度,即预测错误“存在”的概率)为横坐标、真阳性率(灵敏度,即预测正确“存在”的概率)为纵坐标所形成的曲线。其曲线下面积(area under curve, AUC)值提供了一个衡量模型预测精度的指标。AUC值不受阈值和发生率的影响,AUC值越大,环境变量与模型的相关性越高,即模型预测精度就越高。当AUC值在0.5~0.7时模型的预测精度较低,在0.7~0.9时预测较准确,大于0.9 时预测精度最高,一般来讲,当模型的AUC值>0.75时,认为构建的模型精度可接受[29]。
2 结果与分析
2.1 预测精度
ROC分析表明,C3、C4植物功能群预测的训练数据AUC值均达到了0.9,测试数据AUC值也达到了0.8(图1),表明两个功能群的MaxEnt模型均具有较好的预测能力,可用于预测C3和C4物种的适宜分布。
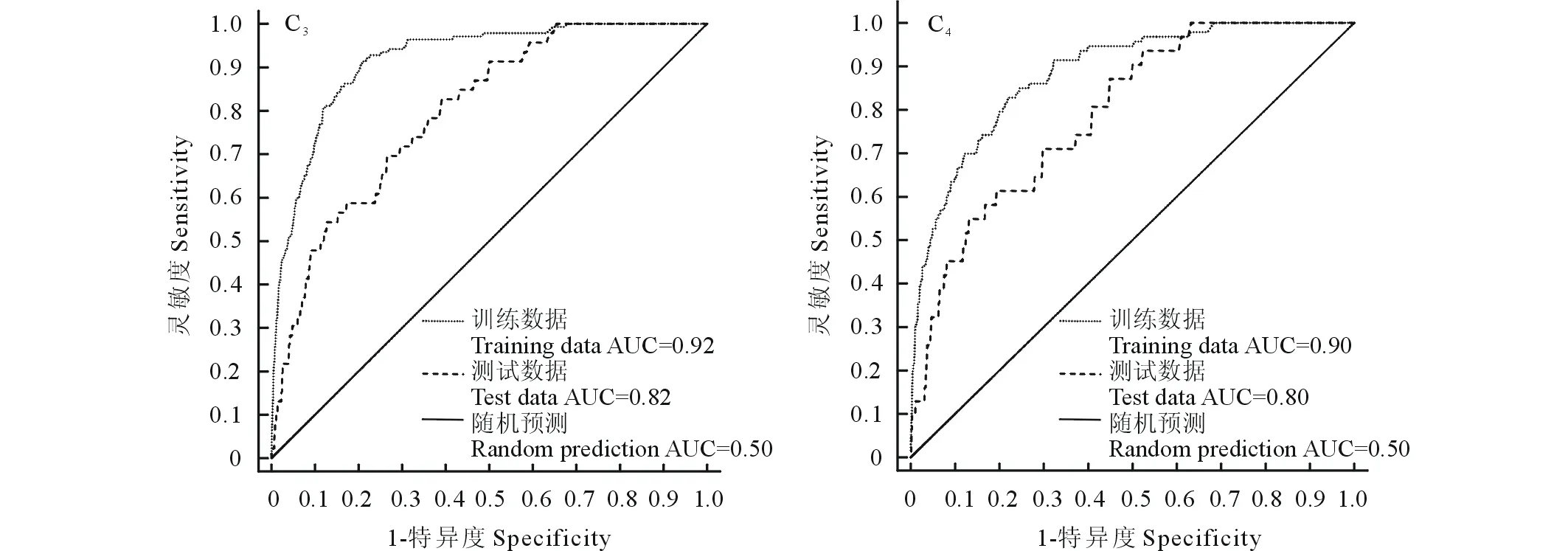
图1 MaxEnt模型精度检验的ROC曲线Fig.1 The receiver operating characteristics curves of test for the MaxEnt models of C3 and C4 herbaceous plant functional groups
2.2 适宜分布格局
C3和C4草本功能群的潜在适宜分布格局呈现明显的差异(图2)。C3适宜分布阈值为0.37,适宜分布面积较大,集中于研究区的中部,占整个研究区面积的61.99%;C4适宜分布阈值为0.40,适宜分布面积较小,零散分布在研究区的中北部,占研究区总面积的19.12%(表2)。
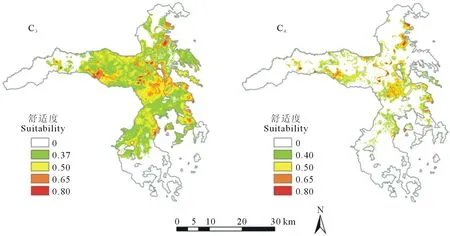
图2 C3、C4草本植物功能群的潜在分布Fig.2 The potential distribution of C3 and C4 herbaceous plant functional groups
2.3 主要环境因子分析
刀切法分析表明,影响C3植物功能群的主要环境因子包括极端最高温(24.92%)、冬季平均降雪量(16.05%)、>5 ℃积温(15.52%)、年平均气温(14.63%)和最冷月均温(14.04%),这5个环境因子累积贡献率为85.16%(图3)。影响C4植物功能群的主要环境变量为极端最高温(21.86%)、>5 ℃积温(15.41%)、年平均气温(14.89%)、年均降水量(14.25%)和冬季平均降雪量(13.94%),这5个环境变量累积贡献率达到80.62%。以上变量能够较好的解释C3和C4植物功能群的分布格局(图3)。
2.4 环境因子对功能群适宜分布的影响
响应曲线分析表明,C3和C4草本植物均有其适宜生长的环境条件(图4)。C3植物发生概率随着冬季平均降雪量(PAS)的增加而增加,而C4植物的发生概率则随着PAS和年均降水量(MAP)的增加而下降(图4a,f);随着年平均气温(MAT)的增加,两个功能群发生概率均增加到一定程度后保持平稳,但C3植物发生概率要高于C4植物(图4b);C3植物发生概率随着极端最高温(EXT)的增加快速下降后趋于平稳,而C4植物发生概率则呈相反的变化趋势(图4c)。C3植物发生概率随>5 ℃积温(DD5)和最冷月平均温度(MCMT)的增加呈下降趋势,但随MCMT的下降幅度更大(图4d,e),与C3植物变化相反,C4植物发生概率随DD5的增加呈增加趋势(图4d)。
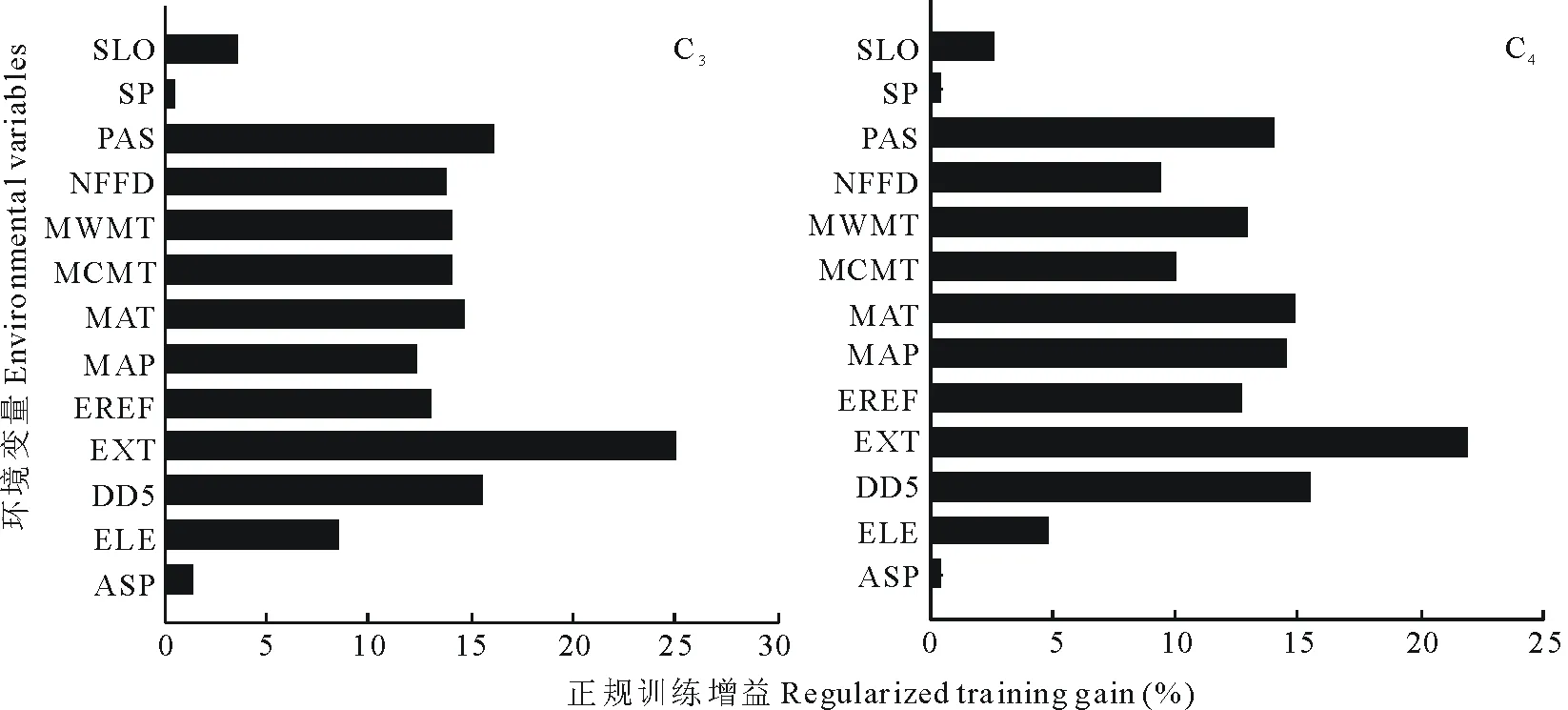
图3 环境变量重要性的刀切法分析Fig.3 Important analysis of environmental variables based on Jackknife test变量代码含义见表1 See Table 1 for environmental factors abbreviations. 下同 The same below.
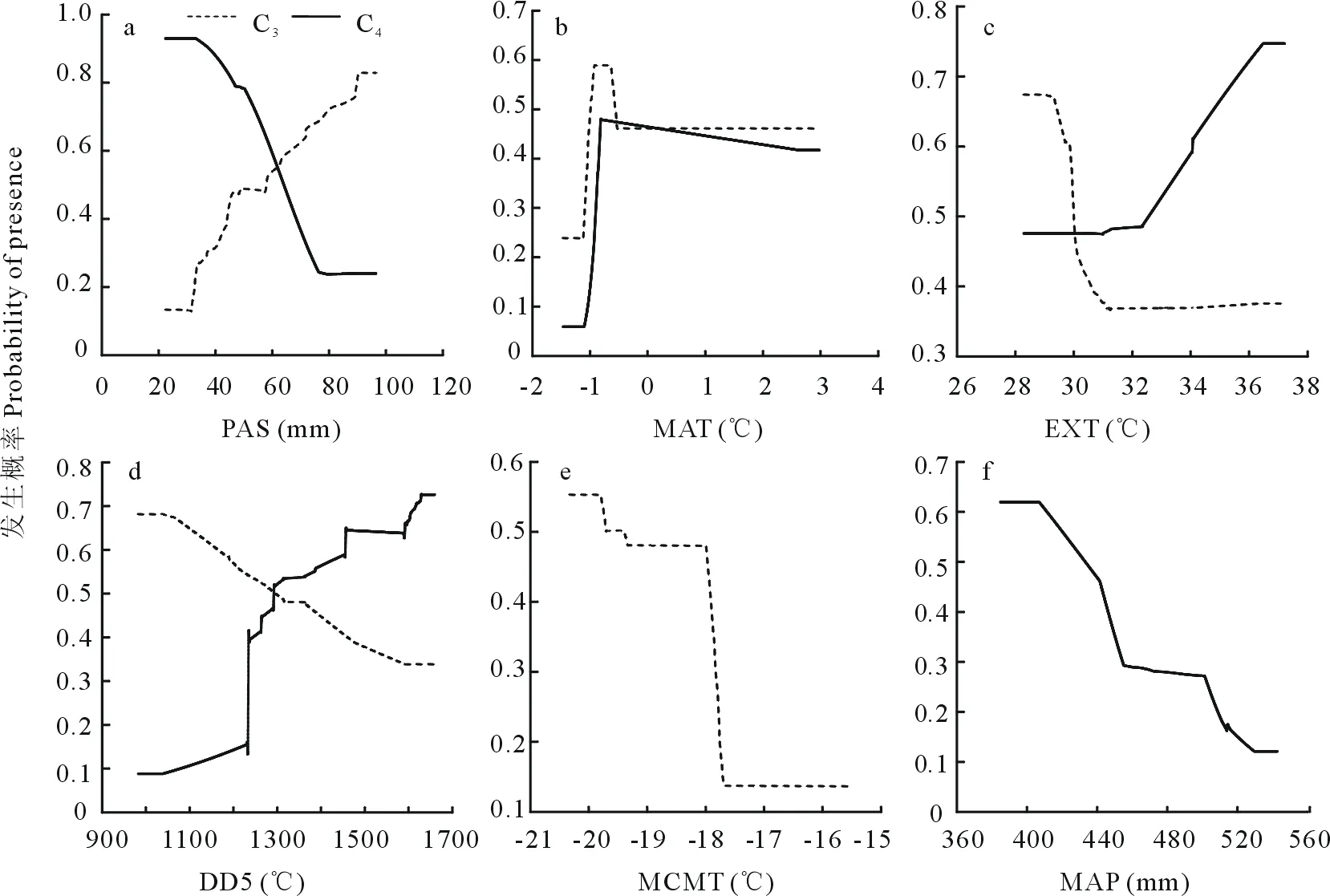
图4 C3、C4草本植物功能群发生概率与主要环境因子的关系Fig.4 Relationships between major environmental variables and the presence probability of the C3 and C4 herbaceous plant functional groups
3 讨论
研究表明C3和C4草本植物功能群的适宜分布格局呈现明显的差异(图2),C3植物适宜分布面积较大,而C4植物分布面积较小,生态位较小。分布格局的差异是C3和C4植物的生理生态特征及环境生态位的不同造成的。
从年均温对C3植物的分布影响来看,C3植物对于较低的年均温有更高的适宜性,在较低的温度下仍能分布,并且其分布概率高于C4植物(图4b)。这与C3植物适合生长在冷凉湿润的生境中的研究结果表现出一致性[6],另一方面,C3植物对于极端低温环境表现出很高的耐受性(图4e),但对极端最高温表现脆弱(图4c)。有研究指出在极寒地区仍有许多C3植物的分布就很好的证明了这一点[30]。并且C3植物发生概率随着>5 ℃积温的增加,呈现出下降趋势,也说明了C3植物对高温的不耐受性(图4d)。塞罕坝地区年平均气温只有-1.5 ℃,冬季漫长、夏季不明显的寒冷大陆性季风型和高原-山地气候特点,造就了C3物种高发生的适宜分布格局。除了温度的影响,冬季降雪量也显著影响了C3植物的适宜分布(图4a)。Bremond等[10]研究指出C3植物的生长季节发生较早,仅有储存在土壤中的春季降水和积雪融水可以利用,因此C3植物生长往往与春、冬季节降雨(雪)量的多少紧密相关。Wertin等[31]也在研究中指出相较于C4植物来说,气候变化引起的土壤水分减少对水分利用效率低的C3植物生长影响更大。本研究随着冬季降雪量的增加,C3植物的分布概率不断增加这一结果充分说明在研究区随着冬季降雪量的增加,显著增加了C3植物生长季土壤中的水分含量,为C3植物的生长提供了必需的水分条件。C3植物与C4植物生理生态差异显著,相对于C4草本物种,C3物种具有高的蒸腾速率、低的光合速率和光饱和点[11]以及低的水分、养分利用效率[4,31],但在低温和冬季降雪量丰富的地区,由前面的分析可知,C3植物却能弥补它的生理生态特征方面的不足并能最大程度地发挥它的光合潜能,适应性更强,分布面积更广。
从各环境因子对C4植物的分布影响来看,C4植物分布概率随着年均温度的升高呈现出增加趋势,但在相对低温环境下分布概率增加趋势要弱于C3植物(图4b)。这与C4植物不耐低温,在生长季节平均最低温度低于8~12 ℃的地区很少发生光合作用[32]的研究结果类似。与C3植物相比,C4植物在高温环境下,气孔开度较小,能通过抑制光呼吸保持高光效的能力[24],这一生理活性特征优势,使其在高温环境下有更高的耐受性和适应能力,这可从本研究随着极端最高温变化的响应曲线得到很好的证明(图4c)。植物在生长发育过程中必须从环境中摄取一定的热量才能完成整个发育过程。有研究表明,C4植物只出现在>10 ℃积温超过1200 ℃的区域[11],而在小于约1250 ℃的>5 ℃的积温区,C4草本植物发生概率较低(图4d)。除了温度,降水量也是限制C4草本植物分布的关键因素[33]。但从本研究结果来看,C4草本植物高发生概率均出现在年均降水量(图4f)和冬季降雪量(图4a)比较低的区域。C4植物适应干燥及土壤贫瘠的环境[6]可以说是与其具有高的水分和养分利用效率及抗逆性较强的生理生态特征密不可分的[4,31]。Osborne等[34]在研究中指出C3植物喜欢生长在郁闭度较高的森林,而C4草本植物则更多的分布在干燥开阔的生境中的结果也进一步验证了这一点。但也有研究表明C4草本植物分布与夏季降水量多少呈正相关[33]。总之,在研究区C4草本植物潜在适宜分布格局往往与高温、降水少等C3草本植物不宜生长的环境条件相联系,虽然其适宜分布面积相对C3草本植物要小,但是随着全球气候变暖的进一步加剧,温度升高可能会导致将来研究区C4植物的优势度逐渐提高,分布面积也将进一步扩大。
本研究对C3、C4植物功能群分布的研究仅选取了气候因子和地形因子,而从其他研究中可知,土壤因素[35]、物种之间的相互作用[24]、CO2浓度[9]等都对C3和C4植物的分布具有显著影响。另外,采取直接功能途径,即直接利用功能性状(植物形态、生理和物候等特征)预测物种对环境变化的响应,将更有利于揭示植物分布与环境因素的耦合关系,因此基于功能性状数据考虑更多环境因子预测物种的分布格局将是今后进一步努力的方向。本研究通过对草本植物进行功能群划分后,结合气候和地形因子,利用MaxEnt模型对研究区C3和C4功能群的潜在分布预测仍取得了较好的效果。研究结果对于揭示物种分布与环境因子的关系,保护和恢复研究区重要C3和C4草本植物资源具有一定的理论和现实意义。
References:
[1] Guisan A, Zimmermann N E. Predictive habitat distribution models in ecology. Ecological Modelling, 2000, 135(2/3): 147-186.
[2] Noble I R, Gitay H. A functional classification for predicting the dynamics of landscapes. Journal of Vegetation Science, 1996, 7(3): 329-336.
[3] Yang X H, Bao Y J, Han G D,etal. Plant functional group and their applications in ecology research. Journal of Dalian Nationalities University, 2009, 11(5): 397-400, 409.
杨晓慧, 鲍雅静, 韩国栋, 等. 植物功能群及其在生态学研究中的应用. 大连民族学院学报, 2009, 11(5): 397-400, 409.
[4] Han M, Yang L M, Zhang Y G,etal. The biomass of C3and C4plant function groups inLeymuschinensiscommunities and theirs response to environmental change along Northeast China transect. Acta Ecologica Sinica, 2006, (6): 1825-1832.
韩梅, 杨利民, 张永刚, 等. 中国东北样带羊草群落C3和C4植物功能群生物量及其对环境变化的响应. 生态学报, 2006, (6): 1825-1832.
[5] Nelson D M, Hu F S, Scholes D R,etal. Using spiral (single pollen isotope ratio analysis) to estimate C3- and C4-grass abundance in the paleorecord. Earth and Planetary Science Letters, 2008, 269(1/2): 11-16.
[6] Pau S, Edwards E J, Still C J. Improving our understanding of environmental controls on the distribution of C3and C4grasses. Global Change Biology, 2013, 19(1): 184-196.
[7] Angelo C L, Daehler C C. Temperature is the major driver of distribution patterns for C4and C3BEP grasses along tropical elevation gradients in Hawaii, and comparison with worldwide patterns. Botany, 2015, 93(1): 9-22.
[8] Tang H P. The C4plant distribution and its correlation with environmental factors in Northeast China transect (NECT) area. Chinese Science Bulletin, 1999, (4): 416-421.
唐海萍. 中国东北样带(NECT)的C4植物分布及其与环境因子的相关性. 科学通报, 1999, (4): 416-421.
[9] Murphy B P, Bowman D. Seasonal water availability predicts the relative abundance of C3and C4grasses in Australia. Global Ecology and Biogeography, 2007, 16(2): 160-169.
[10] Bremond L, Boom A, Favier C. Neotropical C3/C4grass distributions-present, past and future. Global Change Biology, 2012, 18(7): 2324-2334.
[11] Niu S L, Jiang G M, Li Y G. Environmental regulations of C3and C4plants. Acta Ecologica Sinica, 2004, (2): 308-314.
牛书丽, 蒋高明, 李永庚. C3与C4植物的环境调控. 生态学报, 2004, (2): 308-314.
[12] Cavagnaro J B. Distribution of C3and C4grasses at different altitudes in a temperate arid region of Argentina. Oecologia, 1988, 76(2): 273-277.
[13] Sage R F, Wedin D A, Li M. The biogeography of C4photosynthesis: patterns and controlling factors//C4plant biology. San Diego: Academic Press, 1999: 313.
[14] Hernandez P A, Franke I, Herzog S K,etal. Predicting species distributions in poorly-studied landscapes. Biodiversity and Conservation, 2008, 17(6): 1353-1366.
[15] Elith J, Graham C H, Anderson R P,etal. Novel methods improve prediction of species’ distributions from occurrence data. Ecography, 2006, 29(2): 129-151.
[16] Yi Y, Cheng X, Yang Z,etal. MaxEnt modeling for predicting the potential distribution of endangered medicinal plant (H.ripariaLour) in Yunnan, China. Ecological Engineering, 2016, 92: 260-269.
[17] Xu J, Cao B, Bai C K. Prediction of potential suitable distribution of endangered plantKingdoniauniflorain China with MaxEnt. Chinese Journal of Ecology, 2015, (12): 3354-3359.
徐军, 曹博, 白成科. 基于MaxEnt濒危植物独叶草的中国潜在适生分布区预测. 生态学杂志, 2015, (12): 3354-3359.
[18] Zhang X A, Sui X Y, Lü Z,etal. A prediction of the global habitat of two invasive fishes (PseudorasboraparvaandCarassiusauratus) from East Asia using MaxEnt. Biodiversity Science, 2014, (2): 182-188.
张熙骜, 隋晓云, 吕植, 等. 基于MaxEnt的两种入侵性鱼类(麦穗鱼和鲫)的全球适生区预测. 生物多样性, 2014, (2): 182-188.
[19] Yue M F, Feng L, Cui Y,etal. Prediction of the potential distribution and suitability analysis of the invasive weed,Bidensalba(L.) DC. Journal of Biosafety, 2016, (3): 222-228.
岳茂峰, 冯莉, 崔烨, 等. 基于MaxEnt模型的入侵植物白花鬼针草的分布预测及适生性分析. 生物安全学报, 2016, (3): 222-228.
[20] Wen C, Gu L, Wang H,etal. GAP analysis on national nature reserves in China based on the distribution of endangered species. Biodiversity Science, 2015, (5): 591-600.
闻丞, 顾垒, 王昊, 等. 基于最受关注濒危物种分布的国家级自然保护区空缺分析. 生物多样性, 2015, (5): 591-600.
[21] Li B V, Pimm S L. China’s endemic vertebrates sheltering under the protective umbrella of the giant panda. Conservation Biology, 2016, 30(2): 329-339.
[22] Qin P Y, Yang H J, Jiang F L,etal. Quantitative classification of natural plant communities in the Saihanba National Nature Reserve, Hebei Province, China. Chinese Journal of Applied Ecology, 2016, (5): 1383-1392.
秦朋遥, 杨会娟, 蒋凤玲, 等. 河北省塞罕坝保护区天然植物群落数量分类. 应用生态学报, 2016, (5): 1383-1392.
[23] Tian X M, Yan H X, Yuan Y,etal. Response of species richness to the fragmentation of vegetation landscape and its spatial variation scales in Saihanba Nature Reserve. Scientia Silvae Sinicae, 2016, (12): 13-21.
田晓敏, 闫海霞, 袁业, 等. 塞罕坝自然保护区物种丰富度对植被景观破碎化的响应及其空间尺度差异. 林业科学, 2016, (12): 13-21.
[24] Gong C M, Ning P B, Wang G X,etal. A review of adaptable variations and evolution of photo-synthetic carbon assimilating pathway in C3and C4plants. Chinese Journal of Plant Ecology, 2009, (1): 206-221.
龚春梅, 宁蓬勃, 王根轩, 等. C3和C4植物光合途径的适应性变化和进化. 植物生态学报, 2009, (1): 206-221.
[25] Wang T, Wang G, Innes J,etal. Climatic niche models and their consensus projections for future climates for four major forest tree species in the Asia-Pacific region. Forest Ecology and Management, 2016, 360: 357-366.
[26] Wang G, Wang T, Kang H,etal. Adaptation of Asia-Pacific forests to climate change. Journal of Forestry Research, 2016, 27(3): 469-488.
[27] Wang T, Hamann A, Spittlehouse D L,etal. ClimateWNA-high-resolution spatial climate data for Western North America. Journal of Applied Meteorology and Climatology, 2012, 51(1): 16-29.
[28] Zhang Z D, Zang R G. Predicting the distribution of potential natural vegetation based on species functional groups in fragmented and species-rich forests. Plant Ecology and Evolution, 2013, 146(3): 261-271.
[29] Wang Y S, Xie B Y, Wan F H,etal. Application of ROC curve analysis in evaluating the performance of alien species’ potential distribution models. Biodiversity Science, 2007, (4): 365-372.
王运生, 谢丙炎, 万方浩, 等. ROC曲线分析在评价入侵物种分布模型中的应用. 生物多样性, 2007, (4): 365-372.
[30] Rao Z G, Chen F H, Zhang X,etal. Spatial-temporal variations and driving forces of the relative abundance of C3/C4plants in global terrestrial vegetation since the Last Glacial Age. Chinese Science Bulletin, 2012, (18): 1633-1645.
饶志国, 陈发虎, 张晓, 等. 末次冰期以来全球陆地植被中C3/C4植物相对丰度时空变化基本特征及其可能的驱动机制. 科学通报, 2012, (18): 1633-1645.
[31] Wertin T M, Reed S C, Belnap J. C3and C4plant responses to increased temperatures and altered monsoonal precipitation in a cool desert on the Colorado Plateau, USA. Oecologia, 2015, 177(4): 997-1013.
[32] Long S P. C4photosynthesis at low temperatures. Plant Cell and Environment, 1983, 6(4): 345-363.
[33] Paruelo J M, Lauenroth W K. Relative abundance of plant functional types in grasslands and shrublands of North America. Ecological Applications, 1996, 6(4): 1212-1224.
[34] Osborne C P, Freckleton R P. Ecological selection pressures for C4photosynthesis in the grasses. Proceedings Biological Sciences, 2009, 276: 1753.
[35] Wang P, Yin L J, Li J D. Ecological distribution and physiological adaptation to saline-alkali environment of C3and C4plants in Northeastern China prairie area. Chinese Journal of Applied Ecology, 1997, (4): 407-411.
王萍, 殷立娟, 李建东. 东北草原区C3、C4植物的生态分布及其适应盐碱环境的生理特性. 应用生态学报, 1997, (4): 407-411.
