康县隆肛蛙的两性异形
2018-03-26黄棨通龚大洁
黄棨通, 龚大洁
西北师范大学生命科学学院,兰州730070)
两性异形是指同一物种雌、雄个体在身体大小、颜色和局部形态特征等方面存在差异的现象,它普遍存在于整个动物界(Darwin,1871;Shine,1989;Andersson,1994;徐峰,2015)。早在1871年,达尔文在研究生物进化中就对物种的两性异形进行过研究(Darwin,1871)。近年来的研究发现,生育力假设是对两性异形最好的解释(Shine,1979;廖灏泓等,2013);但是,雌雄两性在个体生长和形态上的差异与动物个体的遗传机制、生长环境、食性、生长速率都有密切关系,这些因素综合作用后表现出雌雄外部形态特征的不同(刘洋,时磊,2009;罗煜,2009)。两性异形现象自提出以来,在不同物种中得到验证,无脊椎动物、鱼类、两栖动物的雌性个体普遍大于雄性(Andersson,1994;Blanckenhorn,2005;胡一中等,2015);鸟类、哺乳动物的雄性个体则比雌性大(Temelesetal.,2000;赵晓进,2006;Ceritetal.,2008);而爬行动物的两性异形则无统一模式(Fairbairn & Blanckenhorn,2007;廖灏泓等,2013)。
两栖动物作为由水生动物向陆生动物进化的过渡性脊椎动物,是动物进化史上的重要组成部分(刘凌云,郑光美,2009),所以两栖动物物种的两性异形也受到较广泛的关注,包括蝾螈Trituruscristatus(Malmgren & Thollesson,1999)、黑斑侧褶蛙Pelophylaxnigromaculatus(郑荣泉等,2002)、滇蛙Babinapleuraden(罗煜,2009)、中华蟾蜍Bufogargarizans(李殿伟等,2014)、华南湍蛙Amolopsricketti(胡一中等,2015)等。不同物种在躯体结构、机能以及行为等方面存在差异,且生长发育过程也各不相同,因此,不同物种两性异形的模式不同。对特定物种进行雌雄性别形态差异分析,不仅有利于了解该物种两性异形的产生机制,也有利于了解其进化选择压力。
康县隆肛蛙Feiranakangxianensis隶属于无尾目Anura叉舌蛙科Dicroglossidae隆肛蛙属Feirana,主要生活于海拔800~2 000 m的山区溪流及附近灌丛中(Yangetal.,2011;费梁等,2012;Wangetal.,2014;中国两栖类,2017),现仅分布于我国甘肃省西秦岭南侧陇南山地中的康县及附近山区。目前,对该物种的研究仅见于分子系统发育(Yangetal.,2011)、集群繁殖和混交制交配制度(Wangetal.,2014)等,有关该物种两性异形的研究尚未见报道。由于该物种的分布区域不仅是我国动物地理区划中古北界与东洋界的过渡区域,同时也是两界物种相互渗透的通道之一(姚明灿,2014),因而,对康县隆肛蛙两性异形的研究,不仅可以为后续研究其性选择和进化机制提供更多基础资料,同时也可以为两界物种相互渗透提供参考。
1 材料和方法
1.1 实验材料
于2017年6月18日—7月5日在甘肃省康县豆坝乡(康县隆肛蛙模式产地)采集到成体康县隆肛蛙,共计90只(雌性48只,雄性42只)。用游标卡尺(精确到0.02 mm)测定头体长(SVL)、头长(HL)、头宽(HW)、吻长(SL)、鼻间距(IND)、眼间距(IOD)、眼径(ED)、鼓膜长(TYD)、前臂及手长(LAL)、手长(HAL)、后肢全长(HLL)、足长(FL),用电子秤(精确到0.01 g)称量体质量(BM)(费梁等,1999)。剪蹼法标记已捕捉的个体,以免重复捕捉,测量标记后放回原处(陈旭等,2013)。
1.2 数据处理方法
采用SPSS 23.0处理数据。首先用Levene和Kolmogorov-Smironov分别检验所有测量数据的方差同质性和正态性。用单因素方差分析比较两性头体长差异,采用Pearson相关性检验分析头体长与其他形态学特征的相关性,对与头体长有显著相关性的形态学特征,以头体长为协变量,用单因素协方差分析比较两性间差异。最后,将两性异形差异明显、与头体长显著相关的形态量度进一步与头体长做一元线性回归分析。数据以平均值±标准误(Mean±SE)表示,显著性水平设为α=0.05(郑荣泉等,2002;贺佳飞等,2006;陈旭等,2013)。
异形指数参照Gibbons和Lovich(1990)计算,即:异形指数=1-体型较小性别的平均头体长/体型较大性别的平均头体长。
2 结果
2.1 形态特征的两性异形
康县隆肛蛙体质量、头体长、头长、头宽、吻长、鼻间距、眼间距、眼径、鼓膜长、前臂及手长、手长、后肢全长、足长的比较分析结果显示(表1),雌性形态特征的平均值均略大于雄性。其中,雌性平均头体长为50.52 mm±9.48 mm,雄性平均头体长为46.32 mm±7.02 mm,两性间差异有统计学意义(F=5.509,P=0.021),两性平均头体长比为1.091,异形指数为0.08。
Pearson相关性分析得知,雌雄两性个体的头体长与其他11项形态特征呈显著正相关关系(表2)。协方差分析显示:雌、雄的头长、头宽、吻长、鼻间距、眼间距、眼径、鼓膜长、前臂及手长、手长、后肢全长、足长之间的差异均具有统计学意义,即排除头体长后,雌雄个体在这些形态特征上的差异有统计学意义,且雌性个体的部分形态特征显著大于雄性。一元线性回归分析表明,雌、雄的头长、头宽、吻长、鼻间距、眼间距、眼径、鼓膜长、前臂及手长、手长、后肢全长、足长均与头体长有较明显的线性关系,且雌性吻长、眼间距、眼径、鼓膜长、前臂及手长、手长的斜率大于雄性斜率,其中,以鼓膜长的斜率差异最大(图1,图2)。表明雌性康县隆肛蛙这些局部特征随头体长的生长速率明显大于雄性,尤其是吻长、眼间距、眼径、鼓膜长、前臂及手长、手长的生长速率两性差异最大。
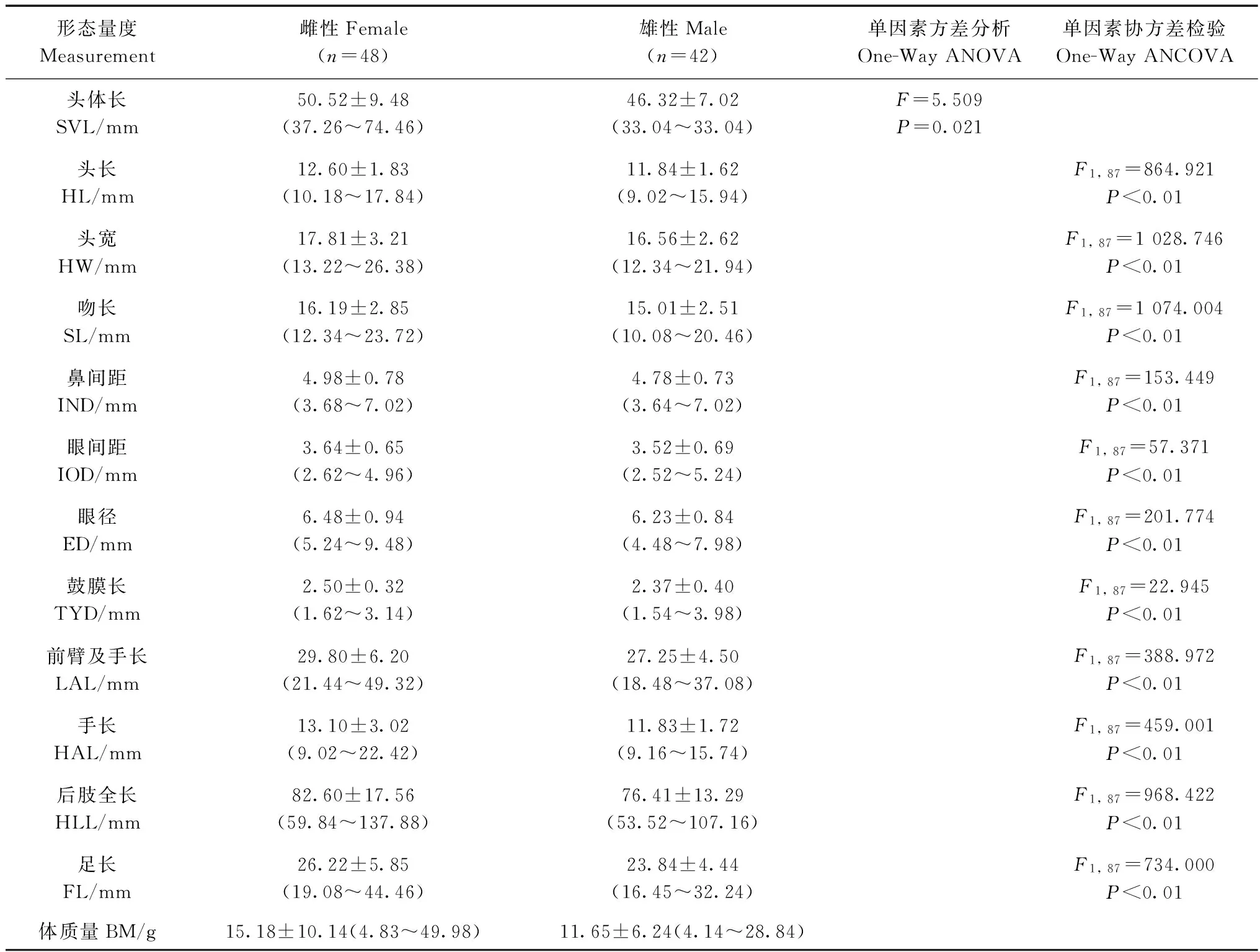
表1 康县隆肛蛙成体形态特征统计Table 1 Morphological characters statistics of adult Feirana kangxianensis
注: 野外测量的体质量具有较大差异, 不作为两性异形的形态指标
Notes: Due to the large differences in field measurements, body mass is not used as a morphological indicator of sexual dimorphism
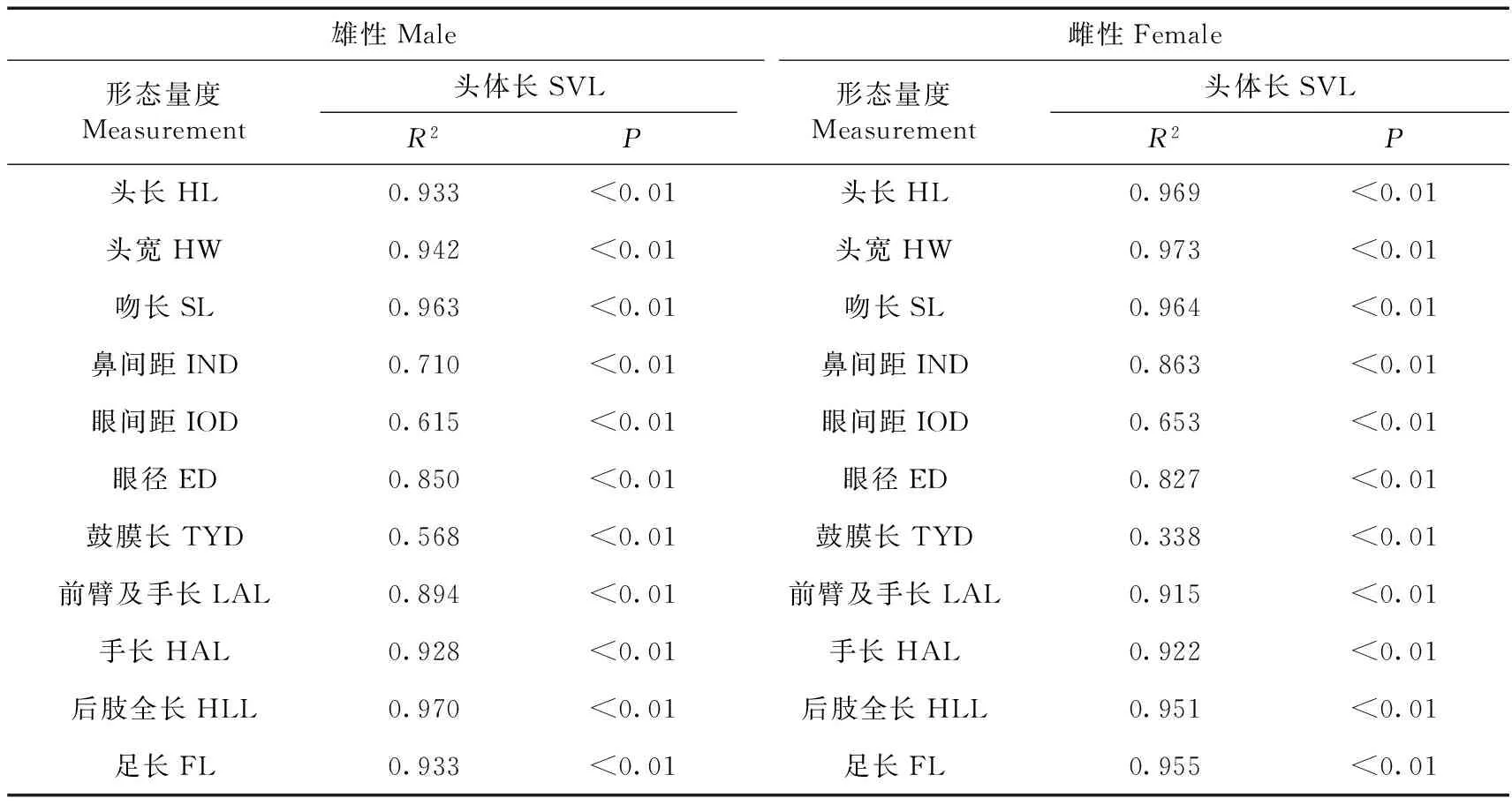
表2 康县隆肛蛙形态特征相关性分析Table 2 Correlation analysis on morphological characters of Feirana kangxianensis
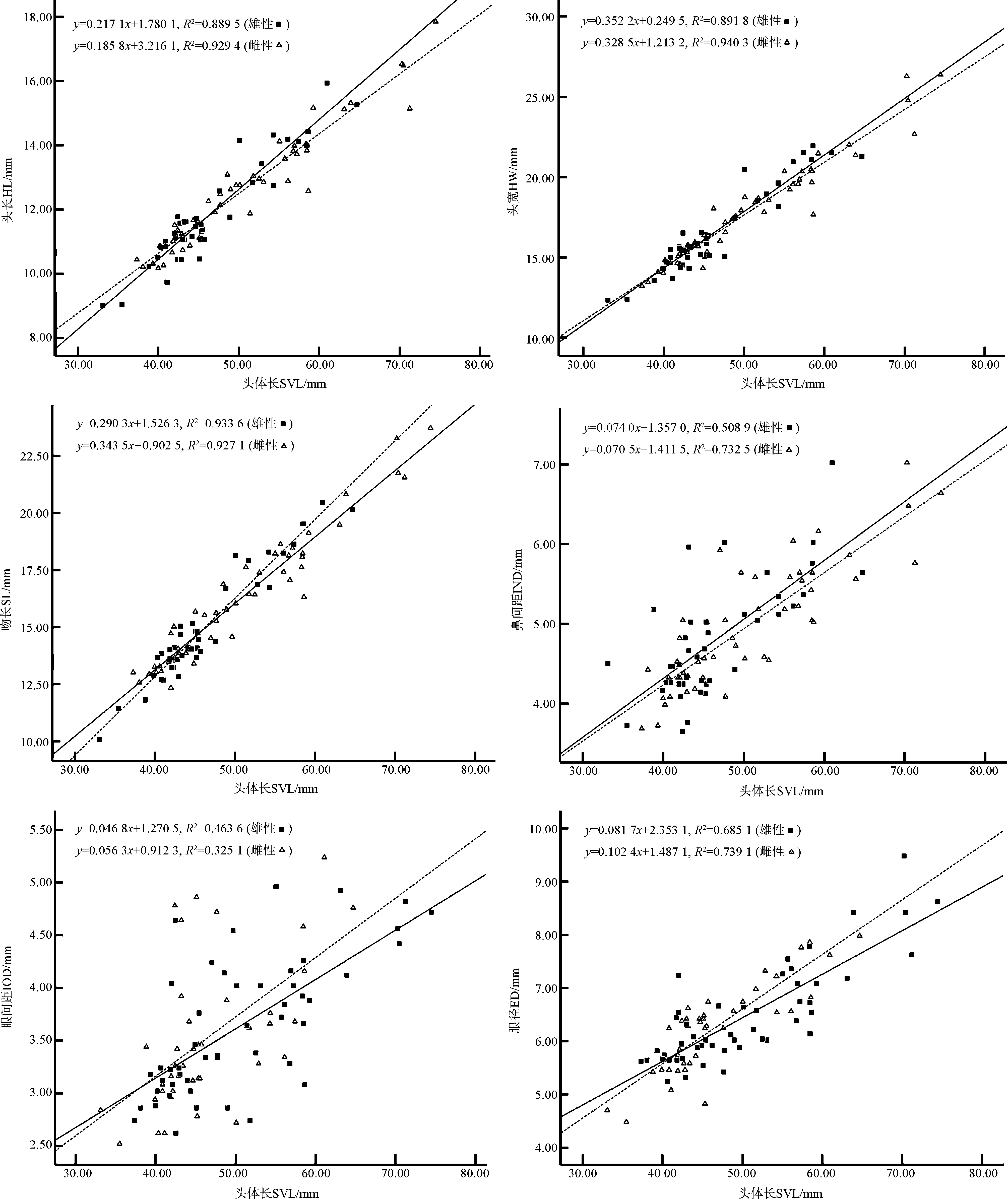
图1 康县隆肛蛙形态特征与头体长的关系Fig. 1 The relationship between morphological characters and snout-vent length of Feirana kangxianensis
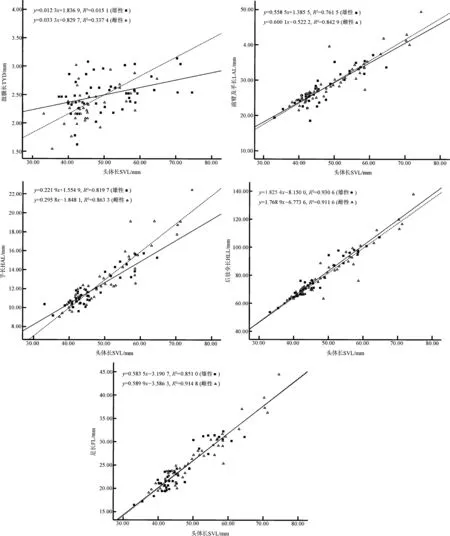
图2 康县隆肛蛙形态特征与头体长的关系Fig. 2 The relationship between morphological characters and snout-vent length of Feirana kangxianensis
3 讨论
两性异形是动物进化发育中普遍存在的现象(Darwin,1871;Andersson,1994;廖灏泓等,2013),而两栖类作为研究动物两性异形极好的生物类群,其个体的两性异形可归为3类:雄性体型大于雌性、雌性体型大于雄性、雌雄体型差异不显著(Powell & Ressell,1985;廖灏泓等,2013)。研究显示,两栖动物中约95%的无尾类和80%的有尾类存在两性异形,而90%的无尾类和61%的有尾类表现为雌性体型大于雄性,蚓螈类为雌雄同型(Shine,1989;李成,2000;胡一中等,2015)。本研究中,雌性康县隆肛蛙体型略大于雄性。由于雌性体型与繁殖输出能力呈正相关性(郑荣泉等,2002;李春瑜等,2013),表明雌性可以通过增加腹腔容纳量来提升繁殖能力和繁殖适合度,这符合生育力选择假设;这一结果也与黑斑侧褶蛙(郑荣泉等,2002)、虎纹蛙Hoplobatrachusrugulosus(林植华,计翔,2005)、金线侧褶蛙Pelophylaxplancyi(寿鹿等,2005)、多疣狭口蛙Kaloulaverrucosa(贺佳飞等,2006)、泽陆蛙Fejervaryamultistriata(施林强等,2011)等相同。因而,导致成体康县隆肛蛙两性体型出现差异的原因可能是:(1)体型通常与性成熟相关(Howard,1981)。当雌性个体大于雄性,说明雌性个体性成熟相对于雄性较晚,而雄性较早的性成熟也更有利于个体遗传物质的传递(李春瑜等,2013)。研究发现,康县隆肛蛙的形态特征均存在两性差异,其中,雌性仅吻长、眼间距、眼径、鼓膜长、前臂及手长、手长显著大于雄性,说明雌性成体体型的增大是部分增大,而不是全身增大。而雄性部分形态特征较小,说明雄性个体在生长发育过程中可能将更多能量分配于繁殖,限制了其个体生长。(2)较大的雌性个体具有较高的生育能力,因而雌性个体有向较大体型方向进化的趋势(郑荣泉等,2002;李春瑜等,2013)。(3)雌性较大的体型有利于提高产卵率,从而提高两栖类的生存率。寿鹿等(2005)指出两栖类雌性的体长和产卵量具有正相关性,即雌性体长越大,繁殖能力越强。(4)野外观察发现,康县隆肛蛙在交配时,雄性个体爬在雌性个体背上并沿溪流岸边寻找产卵处,雄性的体质量完全由雌性负担。因而,雌性较大的体型有利于增强对雄性个体的背负能力;雄性较小的个体也有利于减轻雌性个体的繁殖负重,降低雌性繁殖时的死亡率。
康县隆肛蛙雌性个体的体质量、头体长、头长、头宽、吻长、鼻间距、眼间距、眼径、鼓膜长、前臂及手长、手长、后肢全长、足长等形态指标均大于雄性个体,且差异有统计学意义。康县隆肛蛙异形指数(0.08)和两性平均头体长比(1.091)远小于花臭蛙Ranaschmackeri(异形指数0.496,雌、雄平均头体长比1.818)、大绿臭蛙Odorranamargaretae(异形指数0.476,雌、雄平均头体长比1.814)以及凹耳蛙Odorranatormota(异形指数0.418,雌、雄平均头体长比1.716)(李成,2000;郑荣泉等,2002;寿鹿等,2005;贺佳飞等,2006)。由此可见,康县隆肛蛙属于两性异形程度较小的两栖动物。
本研究结果表明,康县隆肛蛙具有两性异形现象,且雌性大于雄性,符合生育力假设。但也有研究表明,两性差异有可能与生态位、出生率和死亡率、食性及生长阶段中物质和能量分配有关(Powelletal.,1985)。当雄性个体将更多能量、资源等用于繁殖时,雄性个体将缩短成熟期而更早进入繁殖期,这将导致雄性个体小于雌性个体。因此,对康县隆肛蛙两性异形与雌雄两性在食性、能量分配的联系有待进一步研究。
陈旭, 张方, 陈潘, 等. 2013. 凹耳蛙的两性异形[J]. 四川动物, 32(1): 73-77.
费梁, 叶昌媛, 江建平. 2012. 中国两栖动物及其分布彩色图鉴[M]. 成都: 四川科学技术出版社: 480.
费梁. 1999. 中国两栖动物图鉴[M]. 郑州: 河南科学技术出版社.
贺佳飞, 周伟, 李明会, 等. 2006. 多疣狭口蛙的两性异形[J]. 中南林学院学报, 26(1): 77-81.
胡一中, 代亚如, 程岩岩, 等. 2015. 华南湍蛙的两性异形[J]. 浙江师范大学学报(自然科学版), 38(1): 23-27.
李成. 2000. 臭蛙类第二性征的多样性与进化研究[J]. 两栖爬行动物学研究, 8: 28-33.
李春瑜, 洪美玲, 王力军, 等. 2013. 海陆蛙的两性异形和雌性繁殖特征[J]. 动物学杂志, 48(1): 16-21.
李殿伟, 郝婧玮, 金志民, 等. 2014. 中华蟾蜍两性异形研究[J]. 牡丹江师范学院学报(自然科学版), 18(4): 35-37.
廖灏泓, 徐峰, 杨维康. 2013. 两栖爬行动物两性大小异形研究进展[J]. 生态学杂志, 32(11): 3082-3093.
林植华, 计翔. 2005. 浙江丽水虎纹蛙形态特征的两性异形和食性[J]. 动物学研究, 26(3): 255-262.
刘凌云, 郑光美. 2009. 普通动物学(第4版)[M]. 北京: 高等教育出版社.
刘洋, 时磊. 2009. 奇台沙蜥生长过程中的两性异形[J]. 四川动物, 28(5): 710-713.
罗煜. 2009. 滇侧褶蛙的分布、食性、生长及性二型性研究[D]. 贵阳: 贵州师范大学.
施林强, 张秀琴, 马小梅. 2011. 泽陆蛙(Fejervaryalimnocharis)两性异形的个体发育和雌体繁殖[J]. 生态学杂志, 30(4): 717-723.
寿鹿, 杜卫国, 舒霖. 2005. 金线侧褶蛙和泽陆蛙的两性异形与生育力[J]. 生态学报, 25(4): 664-668.
徐峰, 廖灏泓, 杨维康. 2015. 旱地沙蜥的两性异形[J]. 四川动物, 34(6): 859-862.
姚明灿. 2014. 中国两栖动物地理分布格局研究[D]. 长沙: 中南林业科技大学.
赵晓进, 季青, 胡春红, 等. 2006. 太行山猕猴部分活体形态指标的异速生长分析[J]. 河南师范大学学报(自然科学版), 34(4): 139-143.
郑荣泉, 杜卫国, 寿鹿. 2002. 黑斑侧褶蛙的两性异形和雌性繁殖特征[J]. 动物学杂志, 37(4): 14-19.
中国两栖类. 2017. “中国两栖类”信息系统[EB/OL]. [2017-08-31]. http://www.amphibiachina.org/.
Cerit H, Avanus K, 李亨. 2008. 鸟类性别鉴别方法的研究进展[J]. 中国家禽, 30(18): 30-33.
Andersson MB. 1994. Sexual selection[M]. Princeton: Princeton University Press.
Blanckenhorn WU. 2005. Behavioral causes and consequences of sexual size dimorphism[J]. Ethology, 111(11): 977-1016.
Darwin C. 1871. The descent of man, and selection in relation to sex[M]. London: John Murray.
Fairbairn DJ, Blanckenhorn WU. 2007. Sexual size dimorphism in mammals[M]. New York: Oxford University Press: 16-26.
Gibbons JW, Lovich JE. 1990. Sexual dimorphism in turtles with emphasis on the slider turtle (Trachemysscripta) [J]. Herpetological Monographs, 4(1): 1-29.
Howard RD. 1981. Sexual dimorphism in bullfrogs[J]. Ecology, 62(2): 303-310.
Malmgren JC, Thollesson M. 1999. Sexual size and shape dimorphism in two species of newts,TrituruscristatusandT.vulgaris(Caudata: Salamandridae)[J]. Journal of Zoology London, 249(2): 127-136.
Powell GL, Ressell AP. 1985. Growth and sexual size dimorphism in Alberta populations of the eastern short-horned lizard,Phrynosomadouglassibrevirostre[J]. Canadian Journal of Zoology, 63(1): 139-154.
Shine R. 1979. Sexual selection and sexual dimorphism in the Amphibia[J]. Copiea: 297-306.
Shine R. 1989. Ecological causes for the evolution of sexual dimorphism: a review of the evidence[J].The Quarterly Review of Biology, 64(4): 419-461.
Temeles EJ, Pan IL, Brennan JL,etal. 2000. Evidence for ecological causation of sexual dimorphism in hummingbird[J]. Science, 289(5478): 441-443.
Wang J, Xie F, Wang G,etal. 2014. Group-spawning and simultanous polyandry of a stream-dwelling frogFeiranakangxianensis[J]. Asian Herpetological Research, 5(4): 240-244.
Yang X, Wang B, Jiang JP,etal. 2011. A new species of the genusFeirana(Amphibia: Anura: Dicroglossidae) from the western Qinling Mountains of China[J]. Asian Herpetological Research, 2(2): 72-86.
