情绪学习促进无意识信息进入意识*
2018-03-24张秀玲庞兆阳姜云鹏
张秀玲 庞兆阳 姜云鹏 张 明 蒋 毅
(1东北师范大学心理学院, 长春 130024) (2清华大学生物工程医学系, 北京 100084)
(3苏州大学教育学院心理学系, 苏州 215000) (4中国科学院心理研究所, 北京 100101)
1 引言
有效提取环境中的关键信息对个体的生存意义重大。大量研究发现, 与中性信息相比, 人类对带有情绪信息的刺激存在明显的加工优势(Hedger,Adams, & Garner, 2015)。而且这种优势广泛存在于知觉、注意和记忆等认知过程中。情绪影响视知觉的一个最普遍的例子是人们对情绪相关信息的辨别敏感度要高于中性信息(Phelps, Ling, & Carrasco,2006)。类似地, 在注意方面, 带有情绪信息的干扰物比中性的干扰物在视觉搜索中更能捕获注意, 从而降低了被试对目标的搜索速度(Schmidt, Belopolsky,& Theeuwes, 2014)。
近年来, 大量无意识信息加工领域的研究也支持情绪信息的知觉加工优势。Yang, Zald和Blake(2007)使用突破连续闪烁抑制范式(breaking continuous flash suppression, b-CFS)发现, 即使是在无意识的状态下, 恐惧信息仍比其他信息更快地突破噪音的抑制进入意识。此外, 其他不可见的情绪刺激, 例如愤怒的面孔(Gray, Adams, Hedger,Newton, & Garner, 2013)和愤怒的体态(Zhan,Hortensius, & de Gelder, 2015)也能在CFS中更快地突破抑制, 而且在CFS中受到抑制的情绪刺激依然能够捕获注意以及产生情绪启动效应(Almeida,Pajtas, Mahon, Nakayama, & Caramazza, 2013; Jiang,Costello, Fang, Huang, & He, 2006)。同时, 这些研究结果也得到脑成像研究证据的支持, 使用双眼竞争的研究发现被抑制的恐惧面孔显著地激活了杏仁核的腹侧区域(Lerner et al., 2012)。类似地, 采用后掩蔽范式的研究也发现了恐惧的眼睛比高兴的眼睛引起了更强的杏仁核活动(Whalen, 2004)。多数研究者认为这一优势的存在是因为大脑中存在一条绕过视觉联合皮层的从上丘到枕核最终到达杏仁核的皮层下通路, 被称为视觉加工的“低通路”(LeDoux, 2000)。
然而关于情绪信息的无意识加工优势的研究皆采用情绪面孔或情绪场景做为刺激材料, 这就造成研究结论饱受争议。即存在这种可能性:无意识阶段的加工优势有可能不是由刺激本身的情绪效价造成的, 而是由刺激的物理特征造成的。恐惧面孔与中性面孔眼白大小不同, 从而导致眼睛部位的亮度和对比度等方面存在差异。无论是整体的还是局部的亮度、对比度以及空间频率这些物理属性对双眼竞争的影响巨大(Gayet, Paffen, Belopolsky,Theeuwes, & van der Stigchel, 2016), 是我们做实验时需要严格控制的额外变量。Yang等人(2007)虽然发现在无意识条件下, 恐惧面孔比中性面孔突破抑制更快, 但他的研究同时也发现即使是在面孔倒置的情况下, 恐惧面孔仍比中性面孔突破抑制时间更短, 说明所观察到的恐惧面孔知觉加工优势很有可能是由于某些低级的物理特征造成的。事实上, 在情绪的注意优势领域, 有研究采用视觉搜索任务报告了愤怒面孔在干扰物中存在平行加工优势(Hansen & Hansen, 1988), 但是后来的研究控制了刺激的低级物理属性这一额外变量后并没有发现情绪的平行加工优势(Purcell, Stewart, & Skov,1996)。这一事实更提醒研究者考察情绪的视觉加工优势时控制刺激物理属性的重要性。
近年来有研究做了这方面的努力, 尝试消除低级物理属性的影响, 如采用情绪面孔的图示刺激,Stein和 Sterzer (2012)发现正性情绪图示面孔比负性图示面孔能更快地在CFS中突破抑制。尽管图示刺激能够在颜色和亮度等方面得到匹配, 但是表征不同情绪的图示面孔在眉毛、眼睛以及嘴角弧度上仍然存在细微差异。更为致命的是, 当消除图片中的面孔以及情绪信息而保留嘴角上弯或下弯的线条时得出了相同的结果, 这说明影响图片突破抑制时间的并非是面孔的情绪效价。因此情绪面孔的图示刺激不能用来考察无意识条件下的情绪加工优势。
为了解决上述问题, 本研究采用情绪学习这一方法, 在物理属性上完全匹配的刺激(不同朝向的光栅)与无条件恐惧刺激(unconditioned stimuli,UCS)反复联结, 使原本中性的刺激成为具有情绪效价的条件刺激(conditioned stimuli, CS)。以往很多脑成像研究揭示出情绪学习后初级感觉皮层(如视觉和嗅觉)的活动增强或联结增强, 发生了可塑性的变化(Damaraju, Huang, Barrett, & Pessoa, 2009;Li, Howard, Parrish, & Gottfried, 2008)。这些脑可塑性的证据暗示情绪学习也会引起行为上知觉的易化。而且有一个研究揭示出情绪学习降低了光栅的检测阈限(Padmala & Pessoa, 2008), 虽然阈限的下降即知觉敏感性的提高并不意味着无意识条件下情绪刺激也可以得到加工, 但我们推测情绪学习也许能够促进无意识条件下的情绪加工。
因此, 本研究将以物理属性上完全匹配的不同朝向的光栅(实验 1)以及不同颜色与朝向的整合光栅(实验 2)为刺激材料, 采用情绪学习的手段并结合突破连续闪烁抑制范式(b-CFS)来考察无意识条件下经过情绪学习的刺激是否具有加工优势。
2 实验1:情绪学习促进简单刺激更快地进入意识
实验1通过情绪学习中的厌恶性条件反射, 将中性刺激 Gabor patch与无条件刺激警报声(UCS)反复联结, 使原本不具有情绪意义的中性刺激变为带有情绪效价的条件刺激(CS), 考察经过情绪学习后的刺激是否在无意识中存在加工优势。即在CFS中经过情绪学习的Gabor patch是否比没有经过情绪学习的Gabor patch更快地突破抑制。
2.1 方法
2.1.1 被试
随机选取了20名大学生志愿者, 其中12名女生8名男生, 平均年龄为22.5岁, 视力或矫正视力正常, 且均为右利手。实验前签署知情同意书, 并在实验后给予一定报酬。
2.1.2 实验材料和仪器
实验刺激呈现在21寸纯平CRT显示器Iiyama MA203DT Vision Master Pro 513上, 分辨率为1024×768, 刷新频率为85 Hz。实验程序采用Matlab r2008b和 Psychophysics Toolbox-3编写(Brainard,1997; Pelli, 1997)。实验过程中要求被试将下颌放在颌托上, 使被试眼睛距离屏幕保持在52 cm。通过立体镜使屏幕上左侧的信息投射进左眼, 右侧的信息投射进右眼。
屏幕背景为灰色, 左右各呈现一个13.5°×13.5°的正方形刺激区域, 框粗 1.7°, 距屏幕中央 12.7°,每个刺激区域有一个黑色十字“+”作为中央注视点,大小为 0.5°×0.5°。无条件刺激(UCS)为 100 dB 的警报声(https://www.ee.columbia.edu/~dpwe/sounds/),经Goldwave处理, 警报声将在光栅出现1 s后呈现,持续时间为1 s并包含10 ms的淡入淡出。条件刺激(CS)为直径 4.3°的 Gabor patch, 旋转 45°和 135°分别作为 CS+和 CS–, 并进行被试间平衡, 即一半的被试45°作为CS+, 135°作为CS–, 另一半被试与之相反。
2.1.3 实验设计与程序
实验1为单因素2水平被试内设计。自变量为学习条件:伴随有警报声(UCS)的 Gabor patch为CS+, 而没有伴随警报声的Gabor patch为CS–。以旋转45°和135°的Gabor patch分别作为CS+和CS–,并进行被试间平衡。因变量为刺激的突破抑制时间(suppression time, ST), 即从图片开始呈现到被试知觉到图片的时间间隔。ST与进入意识的速度成反比, 即ST越大进入意识的速度就越慢, 反之越快。
实验 1分为学习阶段和测试阶段两个部分, 并在实验结束后让被试对警报声(UCS)做情绪等级评定, 评定根据不愉快程度、强烈程度和警觉程度三个方面进行, 均为1~5分评定, 分数越高程度越强烈。
学习阶段采用经典条件学习范式。分为2个区组(block), 每个 block包含 40个试次(trial), 共 80个trial。每个试次中会呈现一种角度的刺激, 45°与135°各占一半。每个trial的流程如图1所示, 首先呈现800~1200 ms的中央注视点, 随后呈现200 ms的Gabor patch, 最后是1 s的中央注视点, 如果是CS+条件, 被试在这 1 s内听到令人不愉悦的警报声(UCS)。如果是CS–条件, 被试则不会听到警报声。
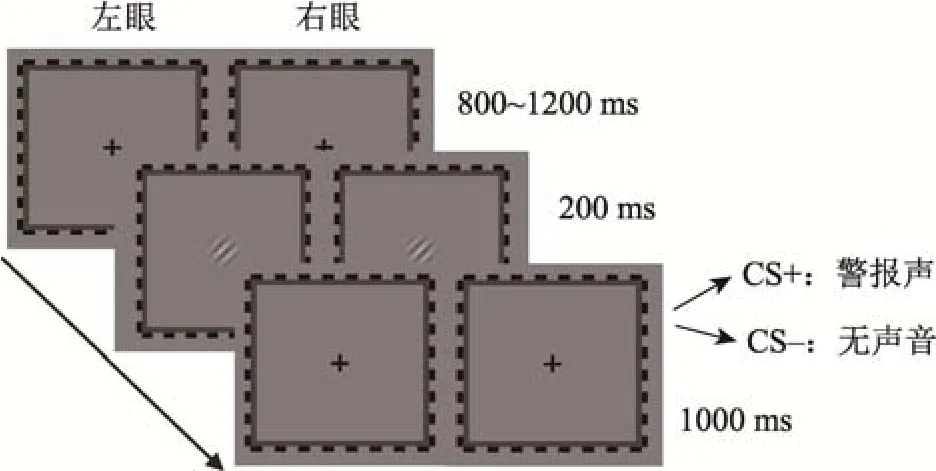
图1 学习阶段流程图。在学习阶段, 给被试的双眼同时呈现相同的Gabor patch。其中一种朝向的Gabor patch伴随警报声(CS+), 而另一种朝向的 Gabor patch则不伴随警报声(CS–)。被试在学习阶段不需要按键反应。
测试阶段分为 3个block, 每个 block包含 40个 trial, 共 120个 trial, 其中靶刺激随机出现在左右眼各60次。实验开始前为使被试适应实验任务,熟悉实验流程, 设置了20次练习。测试阶段采用突破连续闪烁抑制范式(b-CFS)使视觉刺激从无意识进入意识(Jiang, Costello, & He, 2007)。每个trial的流程如图2所示, 首先向两眼呈现800~1200 ms的中央注视点。随后, 向其中一只眼睛呈现标准动态噪音图(蒙德里安图片), 同时向另一只眼睛呈现靶刺激, 且噪音和靶刺激出现的眼睛是随机的。靶刺激呈现的位置在正方形边框内随机, 并在1 s内对比度从0逐渐上升到100%。被试的任务是判断靶刺激的位置, 在“+”的左侧按“←”键, 右侧按“→”键。被试按键反应后进入下一个trial; 若经过12 s后仍无反应, 则自动进入下一个trial。
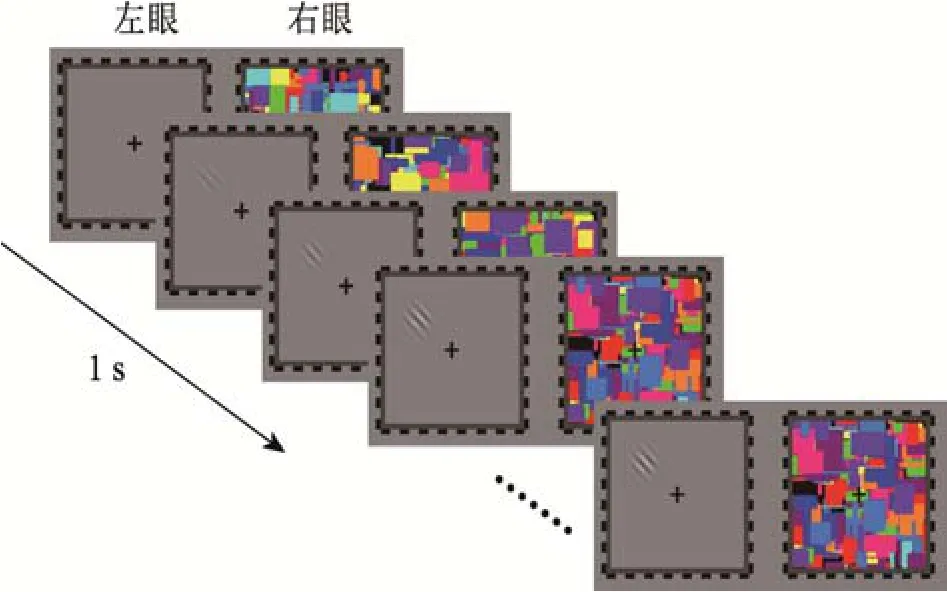
图2 测试阶段流程图。在测试阶段, 给被试的一只眼睛呈现Gabor patch, 而另一只眼睛呈现动态蒙德里安噪音图来抑制目标刺激进入意识。
2.2 结果与分析
2.2.1 声音及刺激的情绪等级评定
被试对警报声(UCS)的情绪等级评定结果显示,这种声音引起了一定程度的负性情绪。将“不愉悦度”的值与 1比较(1代表完全没有感觉到不愉悦),并进行单样本t检验: M = 2.35, SD = 1.27, t(19) =4.76, p < 0.001。将“强度”的值与2进行比较, 并进行单样本t检验: M = 2.85, SD = 0.93, t(19) = 4.07, p =0.001。将“警觉性”的值与2.5进行比较, 并进行单样本t检验: M = 3.1, SD = 1.25, t(19) = 2.14, p <0.05。
2.2.2 正确率
20名被试的正确率基本接近100%, CS+ (M =0.99, SD = 0.03)与 CS– (M = 0.99, SD = 0.02)之间的正确率没有显著差异, t(19) = 0.806, p > 0.05。
2.2.3 突破抑制时间(ST)
剔除突破抑制时间(ST)大于10 s的结果, 这种结果远大于平均值加上 3个标准差。Jiang等人(2007)认为, 如果超过10 s靶刺激仍然没有突破抑制, 那么最终得到的 ST将有可能包含了一些未知因素。总体上看, 剔除的数据量仅占总数的1.2%。对CS+和CS–的ST进行统计分析, 结果如图3所示。CS+的平均值为1.96, 标准差0.72; CS–的平均值为 2.26, 标准差 0.96。两者差异显著, t(19) =–2.82, p < 0.05, d = 0.63。说明CS+比CS–更快地进入了意识。
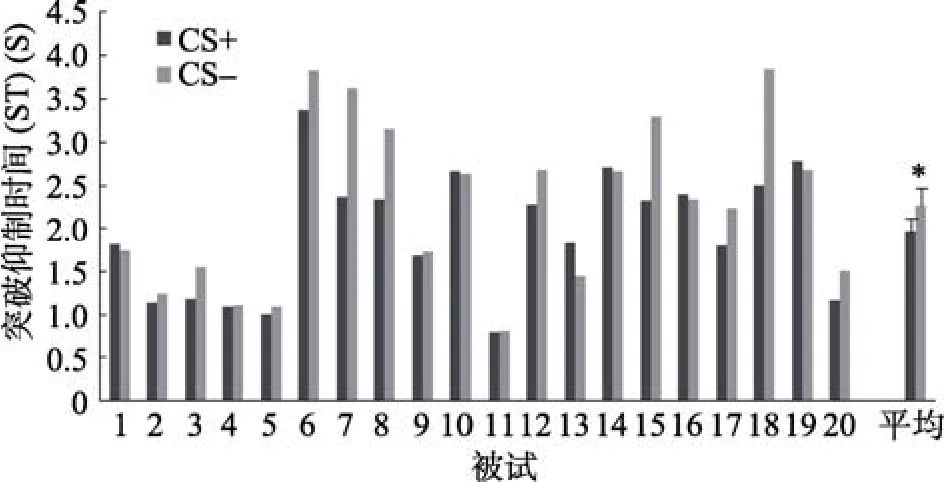
图3 实验1结果。每个被试对于CS+刺激和CS–刺激的突破时间以及所有被试对这两种刺激的平均突破时间, 误差线为标准误(SE)。
3 实验2:情绪学习促进复杂刺激更快地进入意识
实验1中被试对不同朝向的光栅进行了情绪学习, 发现学习过程中伴随警报声的光栅比没有伴随警报声的光栅在随后的无意识任务中能更快地突破抑制、进入意识。朝向是一种简单的视觉属性, 我们会很自然地问, 当条件刺激更复杂时, 即由两个维度(颜色和朝向)整合成的刺激, 是否依然存在这种无意识加工优势效应。如果存在, 这种加工优势是特异性的, 只存在于整合刺激, 还是会泛化到与该复杂刺激具有某一相同属性的其他刺激上。因此实验2采用和实验1相同的实验方法, 材料由原来只在朝向上变化的一维简单刺激变换为在颜色和朝向上变化的二维复杂刺激, 考察经过情绪学习的复杂刺激在无意识加工中的优势模式是怎样的。
3.1 方法
3.1.1 被试
20名大学生志愿者参加实验, 其中 9名男生,平均年龄为 22.23岁, 视力或矫正视力正常, 均为右利手。所有被试在实验前签署知情同意书, 并在实验后给予一定报酬。
3.1.2 实验材料与仪器
实验 2中屏幕背景为黑色, 中央注视点“+”为灰色。实验刺激从实验1中的两种变为了四种, 分别为旋转 45°红色、旋转 45°绿色、旋转 135°红色和旋转 135°绿色的 Gabor patch。通过预实验发现被试对这4种Gabor patch的突破抑制时间没有显著差异,F(3, 57) = 0.91,p> 0.05。其他与实验1同。
3.1.3 实验设计与程序
实验采用 2(光栅朝向:45°和 135°) × 2(光栅颜色:红色和绿色)的被试内实验设计。自变量为 4种条件, 分为整合学习条件(伴随警报声出现的特定朝向和颜色的光栅)、朝向学习条件(只有朝向与整合学习中的刺激相同)和颜色学习条件(只有颜色与整合学习中的刺激相同)以及控制条件(朝向和颜色都和整合学习都不同)。因变量为图片的突破抑制时间(ST)。每种朝向和颜色的Gabor patch所对应的学习类型在被试间进行平衡。
除了实验材料不同, 实验2和实验1的实验流程相同(可参考图1和图2), 实验2也分为学习与测验两个部分。并且实验2中四种光栅中只有一种会伴随警报声, 其他Gabor patch仅出现, 不伴随警报声音。
3.2 结果与分析
3.2.1 正确率
20名被试的正确率基本接近 100%, 整合学习(M= 0.997,SD= 0.01)、朝向学习(M= 0.997,SD=0.01)、颜色学习(M= 0.998,SD= 0.01)和控制条件(M= 0.995,SD= 0.01), 在不同条件上的正确率不存在显著的差异。
3.2.2 突破抑制时间(ST)
剔除突破抑制时间(ST)大于10 s的结果, 这种结果远大于平均值加上3个标准差, 且剔除的数据量不超过总数的 1%。在进行统计分析时, 我们使用“学习效应量”来比较不同学习条件下的差异, 其计算方法如下:
学习效应量 = 学习条件(ST) – 控制条件(ST)这样我们得到3种学习类型的效应量, 使用单样本t检验考察这 3种学习的效应量是否显著(与零比较)。其中, 整合学习的学习效应量M= –0.067,SD= 0.12, 学习效应显著,t(19) = –2.557,p< 0.05; 朝向学习的学习效应量M= –0.021,SD= 0.13, 学习效应不显著,t(19) = –0.701,p> 0.05; 颜色学习的学习效应量M= 0.031,SD= 0.11, 学习效应不显著,t(19) = –1.27,p> 0.05。
4 讨论
本研究通过情绪学习的方法匹配了情绪刺激在物理属性上的差异, 从而排除了以往研究中可能存在的额外变量的干扰, 使我们得以揭示只包含情绪效价的信息在无意识中是否存在加工优势。我们采用突破连续闪烁抑制范式(b-CFS)发现在意识条件下进行的学习可以影响到无意识条件下信息的加工, 即经过情绪学习的刺激能在连续闪烁抑制范式(CFS)中更快地突破抑制。并且我们发现, 这种情绪赋予的无意识加工优势不仅仅存在于简单朝向刺激中, 对于复杂的整合刺激也存在无意识情绪加工优势, 同时也说明在无意识条件下, 颜色和朝向是以捆绑的方式加工的。
我们的结果清晰地表明情绪学习可以促进原本中性的刺激在无意识中的加工, 这和以往情绪学习促进注意和知觉过程的研究结果一致。经过情绪学习的刺激(CS+)在点探测任务(Armony & Dolan,2002)以及视觉搜索任务(Notebaert, Crombez, Van Damme, De Houwer, & Theeuwes, 2011)中都展现出了其在注意领域中的加工优势。如 Notebaert等人(2011)发现尽管 CS+刺激并没达到捕获注意的程度,但是它相对于 CS–刺激确实可以优先获得注意加工。另外, Padmala和Pessoa (2008)发现当阈限水平的刺激和电击匹配后, 被试对该刺激的探测率变高,说明情绪学习可以降低被试对刺激的知觉阈限。除了情绪学习获得的效价, 唤醒度也可能影响知觉探测, Woods, Philbeck和Wirtz (2013)发现具有较高唤醒度水平的被试拥有更高的视觉敏感度。在本研究中, 我们发现了情绪学习促使CS+刺激更快地进入意识, 表明情绪学习有助于个体对刺激的知觉加工, 并进一步说明情绪联结有助于无意识阶段的信息加工。
本实验结果和许多研究发现情绪刺激在无意识中存在加工优势的结果一致。大部分研究创造无意识条件主要采用的是后掩蔽以及CFS等范式, 一方面发现了处于无意识状态下的情绪刺激仍能得到加工, 例如产生情绪启动效应(Almeida et al.,2013); 另一方面它们比中性刺激可以引起更强的杏仁核的活动。但是以往的研究采用的情绪刺激多为愤怒面孔(Gray et al., 2013)等本身具有情绪效价的刺激。在本研究中, 我们采用CFS范式则发现了这种优势也可通过情绪学习获得。关键是, 本研究充分地控制了情绪刺激在物理属性上的差异。正如前言所说, 面孔刺激难以匹配其在眼睛区域亮度、对比度和空间频率上的差异。而我们的研究采用不同朝向的光栅进行情绪学习, 并进行被试间的平衡,保证刺激在亮度、对比度和空间频率上完全匹配。
我们不仅发现简单刺激存在情绪学习引起的无意识加工优势, 对于复杂的、朝向和颜色整合的刺激也存在这种优势。而且这种优势具有特异性,即只存在于和学习阶段完全相同的刺激中(朝向和颜色都一致), 而不会迁移到与其相似(仅仅有一个维度一致)的客体上。值得注意的是, 这一结果与Rajimehr (2004)的结果相一致, 该研究使用高空间频率因此被试无法分辨朝向的 Gabor patch, 考察无意识条件下的朝向适应。结果发现, 被试只对与适应刺激完全相同的探测刺激产生了特异性的朝向后效, 而对不同颜色但相同的朝向的刺激却没有产生朝向后效。这些结果说明在无意识条件下, 被试加工的是朝向和颜色整合的刺激, 而不仅仅是朝向或颜色。同时这些结果也为无意识捆绑假说(unconscious binding hypothesis) (Lin & He, 2009)提供了进一步的支持。
大脑视觉皮层的可塑性变化可能让情绪学习后的刺激更快地突破抑制。除了前言中提到的研究(Padmala & Pessoa, 2008), 在另一项研究中也发现情绪学习增强了早期视觉皮层功能性联结(Damaraju et al., 2009), 说明情绪学习除了影响我们所熟悉的杏仁核的活动, 也影响初级视觉皮层的活动。类似地, 在听觉的经典条件作用研究中, 也发现了以听觉区域为主的可塑性改变(Bieszczad & Weinberger,2010)。这种可塑性存在的普遍性暗示我们观察到的情绪学习引起的无意识加工优势可能与初级视觉皮层的活动相关。
本研究发现经过情绪学习的刺激在无意识中的加工优势, 其神经机制也可能与杏仁核有关。Jiang和He (2006)采用CFS范式发现在无意识条件下, 与中性面孔相比, 恐惧面孔能引起更强的杏仁核的活动。而且在无意识条件下, 经过情绪学习的刺激比没有经过经过学习的刺激也可以引起更强的杏仁核活动(Morris, Öhman, & Dolan, 1998)。最近的研究则发现杏仁核不仅能对情绪刺激产生反应, 它还在个体情绪学习尤其是恐惧反应习得过程中起了重要的作用(Knight, Nguyen, & Bandettini,2005), 因此本研究中得出的结果可能是因为经过情绪学习的刺激在无意识条件下激活了杏仁核, 同时视觉皮层也发生了功能上的改变, 从而使刺激更快地突破了抑制。但是杏仁核以及初级视觉皮层是否在情绪学习中同时激活并存在功能上的相互联结尚无文献进行考察。
虽然本研究的结果发现了情绪学习对无意识信息加工的影响, 但是这种情绪学习的影响持续时间如何, 是仅仅只能维持几十分钟, 还是会在一天之后依然存在, 甚至可以延续到一周以上?这一问题有待于进一步的研究。我们发现较为复杂的客体经过情绪学习也存在无意识加工优势, 但是, 这种整合刺激仅仅具有两个不同维度的特征, 对于更复杂的客体如面孔, 是否经过情绪学习后也存在无意识加工优势, 尚待进一步的研究。
5 结论
(1) 经过情绪学习的中性刺激比没有经过情绪学习的中性刺激在无意识加工中存在优势。
(2) 这种优势效应不仅存在于简单的一维刺激中, 在复杂的二维刺激中也存在这种加工优势, 而且这种效应不会泛化到与条件刺激相似的客体上。
Almeida, J., Pajtas, P. E., Mahon, B. Z., Nakayama, K., &Caramazza, A. (2013). Affect of the unconscious: Visually suppressed angry faces modulate our decisions.Cognitive,Affective, & Behavioral Neuroscience,13, 94–101.
Armony, J. L., & Dolan, R. J. (2002). Modulation of spatial attention by fear-conditioned stimuli: An event-related fMRI study.Neuropsychologia, 40, 817–826.
Bieszczad, K. M., & Weinberger, N. M. (2010). Learning strategy trumps motivational level in determining learninginduced auditory cortical plasticity.Neurobiology of Learning and Memory, 93, 229–239.
Brainard, D. H. (1997). The psychophysics toolbox.Spatial Vision, 10, 433–436.
Damaraju, E., Huang, Y. M., Barrett, L. F., & Pessoa, L. (2009).Affective learning enhances activity and functional connectivity in early visual cortex.Neuropsychologia,47,2480–2487.
Gayet, S., Paffen, C. L. E., Belopolsky, A. V., Theeuwes, J., &van der Stigchel, S. (2016). Visual input signaling threat gains preferential access to awareness in a breaking continuous flash suppression paradigm.Cognition,149, 77–83.
Gray, K. L. H., Adams, W. J., Hedger, N., Newton, K. E., &Garner, M. (2013). Faces and awareness: Low-level, not emotional factors determine perceptual dominance.Emotion,13, 537–544.
Hansen, C. H., & Hansen, R. D. (1988). Finding the face in the crowd: an anger superiority effect.Journal of Personality and Social Psychology,54, 917–924.
Hedger, N., Adams, W. J., & Garner, M. (2015). Fearful faces have a sensory advantage in the competition for awareness.Journal of Experimental Psychology: Human Perception and Performance, 41, 1748–1757.
Jiang, Y., Costello, P., Fang, F., Huang, M., & He, S. (2006). A gender- and sexual orientation-dependent spatial attentional effect of invisible images.Proceedings of the National Academy of Sciences of the United States of America,103,17048–17052.
Jiang, Y., Costello, P., & He, S. (2007). Processing of invisible stimuli: Advantage of upright faces and recognizable words in overcoming interocular suppression.Psychological Science,18, 349–355.
Jiang, Y., & He, S. (2006). Cortical responses to invisible faces: Dissociating subsystems for facial-information processing.Current Biology, 16, 2023–2029.
Knight, D. C., Nguyen, H. T., & Bandettini, P. A. (2005). The role of the human amygdala in the production of conditioned fear responses.NeuroImage,26, 1193–1200.
LeDoux, J. E. (2000). Emotion circuits in the brain.Annual Review of Neuroscience, 23, 155–158.
Lerner, Y., Singer, N., Gonen, T., Weintraub, Y., Cohen, O.,Rubin, N., … Hendler, T. (2012). Feeling without seeing?Engagement of ventral, but not dorsal, amygdala during unaware exposure to emotional faces.Journal of Cognitive Neuroscience,24, 531–542.
Li, W., Howard, J. D., Parrish, T. B., & Gottfried, J. A. (2008).Aversive learning enhances perceptual and cortical discrimination of indiscriminable odor cues.Science,319,1842–1845.
Lin, Z. C., & He, S. (2009). Seeing the invisible: The scope and limits of unconscious processing in binocular rivalry.Progress in Neurobiology, 87, 195–211.
Morris, J. S., Öhman, A., & Dolan, R. J. (1998). Conscious and unconscious emotional learning in the human amygdala.Nature,393, 467–470.
Notebaert, L., Crombez, G., Van Damme, S., De Houwer, J.,& Theeuwes, J. (2011). Signals of threat do not capture, but prioritize, attention: A conditioning cpproach.Emotion, 11,81–89.
Padmala, S., & Pessoa, L. (2008). Affective learning enhances visual detection and responses in primary visual cortex.Journal of Neuroscience,28, 6202–6210.
Pelli, D. G. (1997). The videotoolbox software for visual psychophysics: Transforming numbers into movies.Spatial Vision,10, 437–442.
Phelps, E. A., Ling, S., & Carrasco, M. (2006). Emotion facilitates perception and potentiates the perceptual benefits of attention.Psychological Science, 17, 292–299.
Purcell, D. G., Stewart, A. L., & Skov, R. B. (1996). It takes a confounded face to pop out of a crowd.Perception,25,1091–1108.
Rajimehr, R. (2004). Unconscious orientation processing.Neuron,41, 663–673.
Schmidt, L. J., Belopolsky, A. V., & Theeuwes, J. (2014).Attentional capture by signals of threat.Cognition and Emotion,29, 687–694.
Stein, T., & Sterzer, P. (2012). Not just another face in the crowd: Detecting emotional schematic faces during continuous flash suppression.Emotion,12, 988–996.
Whalen, P. J. (2004). Human amygdala responsivity to masked fearful eye whites.Science, 306, 2061.
Woods, A. J., Philbeck, J. W., & Wirtz, P. (2013). Hyper- arousal decreases human visual thresholds.PLoS One, 8, e61415.
Yang, E., Zald, D. H., & Blake, R. (2007). Fearful expressions gain preferential access to awareness during continuous flash suppression.Emotion,7, 882–886.
Zhan, M., Hortensius, R., & de Gelder, B. (2015). The body as a tool for anger awareness-differential effects of angry facial and bodily expressions on suppression from awareness.PLoS One,10, e0139768.
