外源抗坏血酸引发促进Na2SO4胁迫下燕麦幼苗的生长
2018-03-21董秋丽夏方山丁荷星王明亚朱慧森董宽虎杜利霞山西农业大学林学院山西太谷0080山西农业大学动物科技学院山西太谷0080中国农业大学动物科技学院草业科学北京市重点实验室北京009
董秋丽,夏方山,丁荷星,王明亚,朱慧森,董宽虎,赵 祥,杜利霞(.山西农业大学林学院,山西 太谷 0080; .山西农业大学动物科技学院,山西 太谷 0080;.中国农业大学动物科技学院 草业科学北京市重点实验室,北京 009)
土壤盐渍化是当下人类面临的主要生态危机之一[1]。全球有6%的陆地面积及30%的灌溉土地存在着盐碱问题,我国目前约有盐渍化土地3.5×107hm2,其中盐渍化可耕地约有7.6×106hm2,约占总可耕地面积的1/5,并呈逐年增加趋势,致使大面积土地资源难以利用,从而严重地制约了农牧业的可持续发展[2-3]。如何提高植物的抗盐性是实现农牧业增产增收的重要渠道,也是改良盐碱地最为经济有效的手段,因而成为人们长期关注的热点和难点问题[4]。植物种子的萌发至幼苗建成期对外界环境胁迫反应最为敏感,因而对其植株的生长发育具有关键作用[5-6]。土壤盐渍化不仅导致植物种子萌发性能下降,还会滞缓幼苗生长和植株发育,进而降低植物产量及品质,给农牧业生产带来严重的经济损失[1,7]。因此,探究有效促进植物种子萌发与幼苗建成的最佳方法是实现盐渍化土地科学利用的根本途径。种子引发能够有效地改善其在各种生态条件下的活力水平及幼苗的生长能力[8]。采用恰当的生理活性物质引发种子,不仅能激活其萌发前的代谢反应,甚至会直接参与到其萌发过程的代谢反应,从而使其活力水平及幼苗抗逆能力得到有效地提高,并促进其在市场中流通及应用于农牧业生产[9]。目前,采用生理活性物质引发如何提高种子活力及其抗逆性是种子科学领域关注的热点问题。抗坏血酸(AsA)作为植物体内普遍存在的高丰度小分子生理活性物质,参与植物体内众多的物质代谢和氧化还原反应,在种子抵抗氧化胁迫、提高活力、促进细胞分裂和伸长过程发挥着不可替代的作用[9-10]。AsA参与的抗坏血酸-谷胱甘肽循环是整个种子寿命中活跃的动态核心,其快速启动能力被认为是确保种子正常萌发的关键[11]。因此,植物体内AsA的生物学功能研究也越来越受关注[12]。外源AsA能够提高鹰嘴豆(Cicerarietinum)[13]、油菜(Brassicacampestris)[14]、甘蔗(Saccharum)[15]、小麦(Triticumaestivum)[16]及菜豆(Phaseolusvulgaris)[17]等植物耐盐性。但关于外源AsA引发促进盐胁迫下植物幼苗生长的报道较少。
燕麦(Avenasativa)具有耐瘠薄、耐盐碱、抗旱及耐寒等优良品性,作为优质粮饲兼用的特色杂粮作物,在世界干旱和半干旱地区得以普遍种植,其播种面积及总产量仅次于小麦、水稻(Oryzasativa)和玉米(Zeamays)[18-19]。燕麦产草量高,饲用价值优良,能有效解决草地畜牧业生产中冷季缺草问题[20-21]。此外,燕麦还是重要的绿色营养保健作物[22]。燕麦种子含有丰富的不饱和脂肪酸、β-葡聚糖(可溶性膳食纤维)及均衡蛋白质等营养成分,对人类健康起至关重要的作用[23]。燕麦种子油脂含量可达3.1%~11.6%,其不饱和脂肪酸超过80%[24]。由于脂肪衍生物容易酸败或劣变,燕麦种子萌发过程更容易遭受逆境损伤,并延缓其幼苗生长,从而降低其种用价值[25]。土壤盐渍化是严重制约燕麦种子萌发、幼苗生长发育、产量与品质提高的重要因子,尤其春季返盐的影响更大[2,26]。因此,本研究以燕麦种子为材料,分析外源AsA引发对Na2SO4胁迫燕麦幼苗生长性能的影响,以探究最大限度地提高燕麦幼苗耐盐性生长的途径。
1 材料方法
1.1 材料来源
供试燕麦种子来源及信息参照文献[27]。
1.2 AsA引发处理
AsA引发处理具体参照文献[27]进行,每份燕麦种子为20 g。
1.3 发芽试验及指标测定
发芽条件参照国际种子检验协会(International Seed Testing Association,ISTA)的种子检验规程[28]。Na2SO4溶液的浓度为0(CK)、50和100 mmol·L-1,具体试验操作、测定指标与方法均参照文献[27]。
1.4 统计分析
试验数据采用Excel 2010及SAS 8.0软件处理,燕麦幼苗根长、芽长、根鲜重及芽鲜重的多重比较均采用Duncans法进行,最终结果表示为平均值±标准误。
2 结果与分析
2.1 燕麦幼苗芽长在Na2SO4胁迫下响应AsA引发的变化
同浓度AsA引发下,燕麦幼苗芽长随Na2SO4浓度的增加而显著(P<0.05)下降(表1)。Na2SO4浓度为0时,在AsA浓度为0.5~2.0 mmol·L-1时燕麦幼苗芽长显著(P<0.05)高于AsA浓度为0时;Na2SO4浓度为50 mmol·L-1时,随AsA浓度增大燕麦幼苗LS呈现先升后降趋势,浓度为0.5~2.0 mmol·L-1的AsA引发时会显著(P<0.05)高于AsA浓度为0时,并在浓度为1.5 mmol·L-1时达到最大值;Na2SO4浓度为100 mmol·L-1时,燕麦幼苗LS随AsA浓度增大而呈现先升后降趋势,且1.5 mmol·L-1的AsA引发时达到最大值,并显著(P<0.05)高于其他浓度。

表1 燕麦幼苗芽长在Na2SO4胁迫下响应AsA引发的变化Table 1 Responses to AsA-priming on shoot length of oat seedlings under Na2SO4 stress cm
同列不同大写字母表示不同Na2SO4浓度间差异显著(P<0.05),同行不同小写字母表示不同AsA浓度间差异显著(P<0.05)。下同。
Means in the same column with different capital letters are significantly difference among different Na2SO4concentrations at the 0.05 level, in the same row with different lowercase letters are significantly difference among different AsA concentrations at the 0.05 level; similarly for the following tables.
2.2 燕麦幼苗根长在Na2SO4胁迫下响应AsA引发的变化
同浓度AsA引发下,燕麦幼苗根长随Na2SO4浓度的增加而显著(P<0.05)下降(表2)。Na2SO4浓度为0和50 mmol·L-1时,随AsA浓度增加燕麦幼苗根长呈现先升后降趋势,在AsA浓度为0.5~2.0 mmol·L-1时显著(P<0.05)高于其浓度为0时,并在浓度为1.5 mmol·L-1时显著(P<0.05)高于其他浓度;Na2SO4浓度为100 mmol·L-1时,随AsA浓度增加燕麦幼苗根长呈现先升后降趋势,在AsA浓度为0和0.5 mmol·L-1时差异不显著(P>0.05),而浓度为1.5 mmol·L-1时达到最高水平(P<0.05)。
2.3 燕麦幼苗芽鲜重在Na2SO4胁迫下响应AsA引发的变化
同浓度AsA引发下,燕麦幼苗芽鲜重随Na2SO4浓度的增加而显著(P<0.05)下降(表3)。相同浓度Na2SO4胁迫下,随AsA浓度的增加燕麦幼苗芽鲜重均呈现先升后降趋势,并均于1.5 mmol·L-1的浓度时达最大值。Na2SO4浓度为0时,在AsA浓度为0和0.5 mmol·L-1时芽鲜重差异不显著(P>0.05),在AsA浓度为1.0~2.0 mmol·L-1时差异也不显著(P>0.05),但显著(P<0.05)高于其浓度为0和0.5 mmol·L-1时;50 mmol·L-1的Na2SO4胁迫时,浓度为0.5~2.0 mmol·L-1的AsA引发时芽鲜重显著(P<0.05)高于其浓度为0时,但其浓度为1.0~2.0 mmol·L-1时差异不显著(P>0.05);Na2SO4浓度为100 mmol·L-1时,在AsA浓度为0.5~2.0 mmol·L-1时显著(P<0.05)高于其浓度为0时,但其浓度为1.5和2.0 mmol·L-1时差异不显著(P>0.05)。

表2 燕麦幼苗根长在Na2SO4胁迫下响应AsA引发的变化Table 2 Responses to AsA-priming on root length of oat seedlings under Na2SO4 stress cm

表3 燕麦幼苗芽鲜重在Na2SO4胁迫下响应AsA引发的变化Table 3 Responses to AsA-priming on fresh weight of oat seedlings under Na2SO4 stress g
2.4 燕麦幼苗根鲜重在Na2SO4胁迫下响应AsA引发的变化
同浓度AsA引发下,燕麦幼苗根鲜重随Na2SO4浓度增加呈现先升后降趋势(表4),并在Na2SO4浓度为50 mmol·L-1时显著(P<0.05)高于其他浓度,而在浓度为100 mmol·L-1时显著(P<0.05)低于其他浓度。在相同浓度Na2SO4胁迫下,随AsA浓度增加燕麦幼苗根鲜重均呈现先升后降趋势,并在1.5 mmol·L-1的浓度引发时显著(P<0.05)高于0~1.0 mmol·L-1的浓度引发时,但与2.0 mmol·L-1引发时差异不显著(P>0.05);Na2SO4浓度为100 mmol·L-1时,在AsA浓度为0和0.5 mmol·L-1时燕麦幼苗根鲜重差异不显著(P>0.05),Na2SO4浓度为0和50 mmol·L-1时,AsA浓度为0时显著(P<0.05)低于其浓度为0.5~2.0 mmol·L-1时。
2.5 燕麦幼苗活力指数在Na2SO4胁迫下响应AsA引发的变化
同浓度AsA引发下,燕麦幼苗活力指数随Na2SO4浓度增加而显著(P<0.05)下降(表5)。Na2SO4浓度为0~100 mmol·L-1时,随AsA浓度的增加燕麦幼苗活力指数均呈现先升后降趋势,但在Na2SO4浓度为0时,燕麦幼苗活力指数随AsA浓度的变化不显著(P>0.05);Na2SO4浓度为50和100 mmol·L-1时,AsA浓度为0时燕麦幼苗活力指数显著(P<0.05)小于AsA浓度为0.5~2.0 mmol·L-1时;Na2SO4浓度为50 mmol·L-1时,1.5 mmol·L-1的AsA引发下燕麦幼苗活力指数与其浓度为2.0 mmol·L-1时差异不显著(P>0.05),但显著(P<0.05)高于0~1.0 mmol·L-1时;Na2SO4浓度为100 mmol·L-1时,1.5 mmol·L-1的AsA引发下燕麦幼苗活力指数显著(P<0.05)高于其他浓度引发。
2.6 燕麦幼苗生长在Na2SO4胁迫下响应AsA引发的双因素方差分析
不同AsA引发及Na2SO4胁迫浓度对燕麦幼苗根长、芽长、根鲜重、芽鲜重及其活力指数的影响均差异极显著(P<0.01)(表6),不同AsA引发浓度与不同Na2SO4胁迫浓度间的交互作用对燕麦幼苗根长、芽长、根鲜重及芽鲜重的影响均差异极显著(P<0.01),而对其活力指数的影响则差异显著(P<0.05)。
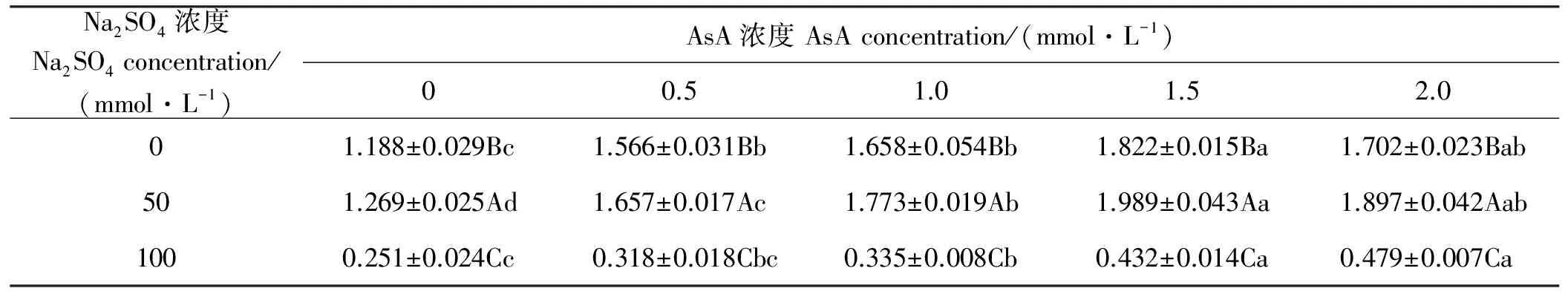
表4 燕麦幼苗根鲜重Na2SO4胁迫下响应AsA引发的变化Table 4 Responses to AsA-priming on fresh weight of roots in oat seed under Na2SO4 stress g
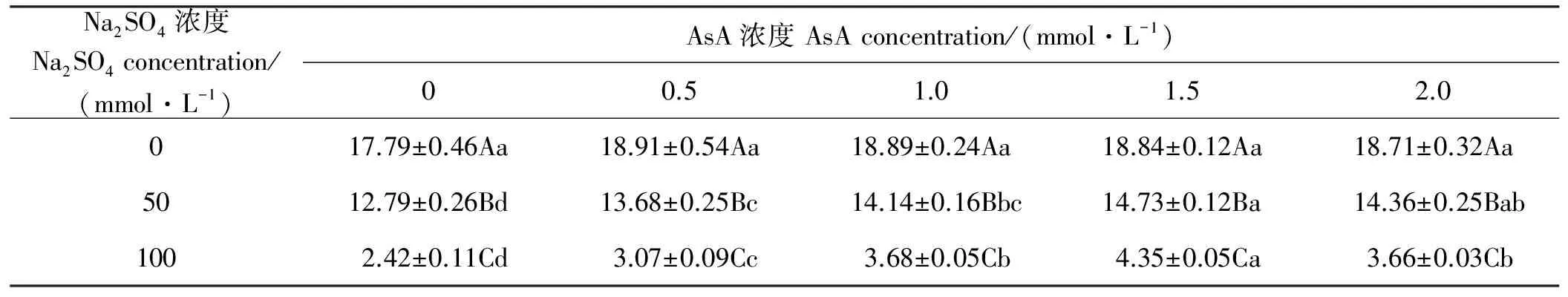
表5 燕麦种子幼苗活力指数在Na2SO4胁迫下响应AsA引发的变化Table 5 Responses to AsA-priming on seedlings vigor index in oat seeds under Na2SO4 stress
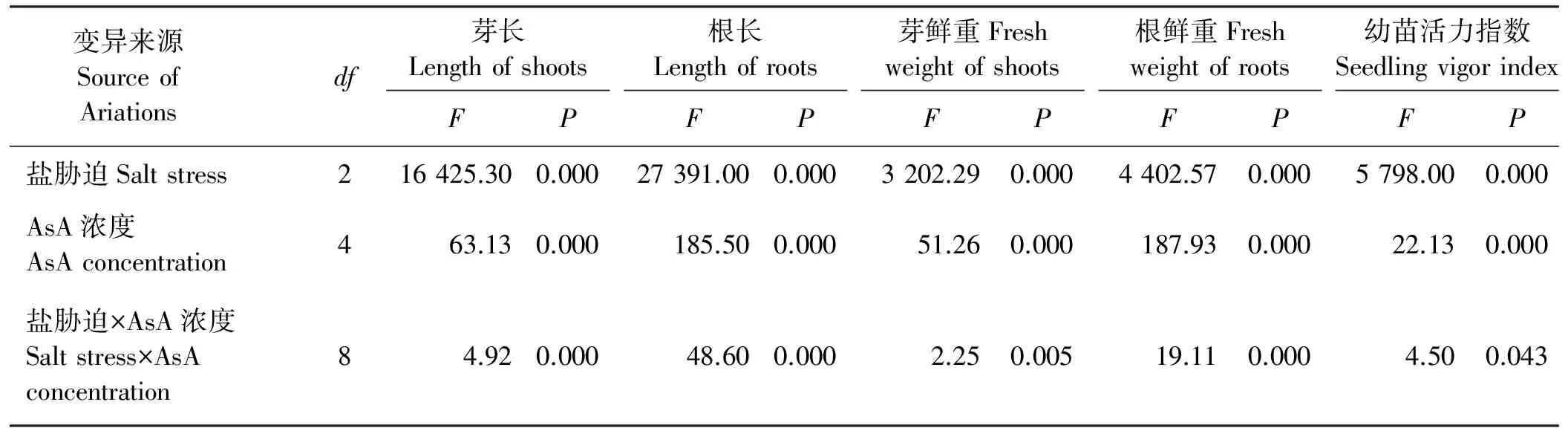
表6 燕麦幼苗生长在Na2SO4胁迫下响应AsA引发的双因素方差分析Table 6 Variance analysis of AsA-priming and Na2SO4 stress on seedling growth of oat seeds
3 讨论
土壤盐渍化是限制植物生长发育最主要、最常见的非生物胁迫。盐胁迫不仅会导致燕麦种子萌发能力下降,还会造成其幼苗生长、产量及品质的降低,甚至植株死亡[2,29]。本研究中,随Na2SO4浓度增加,燕麦幼苗芽长、根长、芽鲜重和活力指数均显著(P<0.05)下降,这说明Na2SO4胁迫降低了燕麦幼苗的生长能力。这与前人的研究结果相似,盐胁迫会抑制燕麦幼苗芽和根的伸长及生物量的增加,这种抑制现象与盐胁迫的强度正相关[1-2,30]。因此,燕麦幼苗生长能力下降是限制其在盐碱地种植的最关键因素。然而,燕麦根鲜重的变化与其盐胁迫的浓度高低有关。本研究中,Na2SO4浓度为50 mmol·L-1时燕麦幼苗根鲜重最高,而其浓度为100 mmol·L-1时最低,这说明50 mmol·L-1的Na2SO4胁迫促进了根的增重,这可能主要依赖于根的增粗来实现的。刘建新等[26]研究发现,50 mmol·L-1的NaCl胁迫会增加燕麦幼苗的根干重,而其浓度大于100 mmol·L-1时会降低其根干重。燕麦幼苗根系在盐胁迫下依赖增粗而实现的增重,可能有利于其选择性吸收及运输矿质离子[30]。
盐胁迫导致的脂质过氧化损伤是制约燕麦幼苗生长的最重要因素[26,31]。然而,AsA不仅能直接清除植物体内过度积累的活性氧,还具有调节植物的生长及延缓其衰老等功能[32]。所以,AsA能促进种子萌发及幼苗生长[33],并能有效提高其抗逆性[34]。研究发现,内源AsA的含量水平与燕麦幼苗的耐盐性生长具有密切关系[26]。本研究表明,相同Na2SO4胁迫浓度下,外源AsA引发能够有效提高燕麦幼苗芽长、根长、芽鲜重、根鲜重和活力指数,这说明外源AsA引发可以有效地促进Na2SO4胁迫下燕麦幼苗的生长。研究盐胁迫下紫花苜蓿(Medicagosativa)[35]、油菜[36]和黄芩(Scutellariabaicalensis)[37]等植物种子的萌发及其幼苗的生长发现,外源AsA引发能促进盐胁迫下植物种子的萌发及其幼苗的生长发育。外源AsA促进盐胁迫下植物种子萌发及幼苗生长的原因主要有两个:一是通过提高其可溶性糖和脯氨酸等渗透调节物质含量来缓解盐胁迫对种子萌发及幼苗生长的抑制作用[36-37];二是通过增强酶促或非酶促抗氧化途径的作用来有效清除胁迫产生的活性氧[38]。然而,外源AsA引发提高盐胁迫燕麦种子活力的机理是否与此相同仍有待于进一步探讨。
植物幼苗耐盐性生长的增强程度与外源AsA的浓度关系密切。江绪文等[37]研究发现适宜浓度(0.50 mmol·L-1)的AsA能提高黄芩幼苗对盐胁迫的适应能力,从而起到缓解盐胁迫对其幼苗生长的抑制作用。范美华等[36]也发现,20 mg·L-1的AsA处理对30%海水胁迫下油菜幼苗生长的缓解效果最好。本研究表明,除Na2SO4与AsA浓度的交互作用对燕麦幼苗活力指数的影响差异显著(P<0.05)外,Na2SO4、AsA浓度及其交互作用对燕麦幼苗芽长、根长、芽鲜重及根鲜重的影响均差异极显著(P<0.01),从而说明Na2SO4胁迫浓度和外源AsA引发浓度均对燕麦幼苗的生长具有密切关系,1.5 mmol·L-1的AsA引发燕麦种子对其幼苗在Na2SO4胁迫下的生长具有最好的促进效果。
4 结论
Na2SO4胁迫会抑制燕麦幼苗的生长,而AsA引发可缓解Na2SO4胁迫对燕麦幼苗生长的抑制作用,这种缓解作用与AsA和Na2SO4的浓度及其交互作用密切相关,采用1.5 mmol·L-1的AsA引发对Na2SO4胁迫的缓解效果最好。
References:
[1] 高战武,蔺吉祥,邵帅,盛后财,范春燕,屈吉宇.复合盐碱胁迫对燕麦种子发芽的影响.草业科学,2014,31(3):451-456.
Gao Z W,Lin J X,Shao S,Sheng H C,Fan C Y,Qu J Y.Effect of complex salt-alkali stresses on seed germination of oat.Pratacultural Science,2014,31(3):451-456.(in Chinese)
[2] 刘凤歧,刘杰淋,朱瑞芬,张悦,郭勇,韩贵清,唐凤兰.4种燕麦对NaCl胁迫的生理响应及耐盐性评价.草业学报,2015,24(1):183-189.
Liu F Q,Liu J L,Zhu R F,Zhang Y,Guo Y,Han G Q,Tang F L.Physiological responses and tolerance of four oat varieties to salt stress.Acta Prataculturae Sinica,2015,24(1):183-189.(in Chinese)
[3] 李培英,孙宗玖.33份偃麦草种质芽期耐盐性评价.草业科学,2015,32(4):593-600.
Li P Y,Sun Z J.Evaluation on the salt resistance of germplasm resources of 33Elytrigriarepensduring seed germination period.Pratacultural Science,2015,32(4):593-600.(in Chinese)
[4] 刘畅,于涛,高战武,于达夫,蔺吉祥.燕麦对松嫩草地三种主要盐分胁迫的生理适应策略.生态学报,2016,36(21):6786-6793.
Liu C,Yu T,Gao Z W,Yu D F,Lin J X.Physiologically adaptive strategies of oat seedling to the three main salts in the soils of the Songnen Plains grasslands.Acta Ecologica Sinica,2016,36(21):6786-6793.(in Chinese)
[5] 夏方山,毛培胜,闫慧芳,王明亚.水杨酸对植物种子及幼苗抗逆性的影响.草业科学,2014,31(7):1367-1373.
Xia F S,Mao P S,Yan H F,Wang M Y.Effect of salicylic acid on stress resistance of seeds and seedling.Pratacultural Science,2014,31(7):1367-1373.(in Chinese)
[6] 张红香,周道玮.种子生态学研究现状.草业科学,2016,33(11):2221-2236.
Zhang H X,Zhou D W.Current status in seed ecology.Pratacultural Science,2016,33(11):2221-2236.(in Chinese)
[7] 王霞霞,李龑,唐杰伟,欧阳灿彬,朱德建,徐智明.盐胁迫对燕麦种子萌发期蛋白质组影响的研究.华北农学报,2015,30(3):48-53.
Wang X X,Li Y,Tang J W,Ouyang C B,Zhu D J,Xu Z M.Analysis of oat seed protein during seed germination under salt stress.Acta Agriculturae Boreali-Sinica,2015,30(3):48-53.(in Chinese)
[8] Xia F S,Chen L L,Yan H F,Sun Y,Li M L,Mao P S.Antioxidant and ultrastructural responses to priming with PEG in aged,ultra-dry oat seed.Seed Science and Technology,2016,44(3):1-13.
[9] Bewley J D,Bradford K J,Hilhorst H W M,Nonogali H.Seeds:Physiology of Development,Germination and Dormancy.3rd.New York:Springer,2013:143-147.
[10] Lee Y P,Baek K H,Lee H S,Kwak S S,Bang J W,Kwon S Y.Tobacco seeds simultaneously over-expressing Cu/Zn-superoxide dismutase and ascorbate peroxidase display enhanced seed longevity and germination rates under stress conditions.Journal of Experment Botany,2010,61(9):2499-2506.
[11] Tullio M C D,Arrigoni O.The ascorbic acid system in seeds:To protect and to serve.Seed Science Research,2003,13(4):249-260.
[12] 俞乐,刘拥海,袁伟超,周丽萍,彭长连.植物抗坏血酸积累及其分子机制的研究进展.植物学报,2016,51(3):396-410.
Yu L,Liu Y H,Yuan W C,Zhou L P,Peng C L.Recent advances in the study of accumulation of ascorbic acid and its molecular mechanism in plants.Chinese Bulletin of Botany,2016,51(3):396-410.(in Chinese)
[13] Beltagi M S.Exogenous ascorbic acid (vitamin C) induced anabolic changes for salt tolerance in chick pea (CicerarietinumL.) plants.African Journal of Plant Science,2008,2(10):118-123.
[14] Khan A,Iqbal I,Shah A,Nawaz H,Ahmad F,Ibrahim M.Alleviation of adverse effects of salt stress in brassica (Brassicacampestris) by pre-sowing seed treatment with ascorbic acid.American-Eurasian Journal of Agricultural and Environmental Science,2010,7(5):557-560.
[15] Ejaz B,Sajid Z A,Aftab F.Effect of exogenous application of ascorbic acid on antioxidant enzyme activities,proline contents, and growth parameters ofSaccharumspp. hybrid cv. HSF-240 under salt stress.Turkish Journal of Biology,2012,36(6):630-640.
[16] 常云霞,徐克东,周琳,陈龙.抗坏血酸对盐胁迫下小麦幼苗生长抑制的缓解效应.麦类作物学报,2013,33(1):151-155.
Chang Y X,Xu K D,Zhou L,Chen L.Ascorbic acid mitigating the inhibition of salt stress to wheat seedling growth.Journal of Triticeae Crops,2013,33(1):151-155.(in Chinese)
[17] Saeidi-Sar S,Abbaspour H,Afshari H,Yaghoobi S R.Effects of ascorbic acid and gibberellin A3 on alleviation of salt stress in common bean (PhaseolusvulgarisL.) seedlings.Acta Physiologiae Plantarum,2013,35(3):667-677.
[18] 郭红媛,贾举庆,吕晋慧,翟志文,靳艳婷,杨武德.燕麦属种质资源遗传多样性及遗传演化关系ISSR分析.草地学报,2014,22(2):344-351.
Guo H Y,Jia J Q,Lyu J H,Zhai Z W,Jin Y T,Yang W D.Analyses of genetic diversities and evolutions ofAvenaL. germplasm resources by ISSR.Acta Agrestia Sinica,2014,22(2):344-351.(in Chinese)
[19] 肖雪君,周青平,陈有军,杜忠,柏晓玲,田莉华,彭先琴.播种量对高寒牧区林纳燕麦生产性能及光合特性的影响.草业科学,2017,34(4):761-771.
Xiao X J,Zhou Q P,Chen Y J,Du Z,Bai X L,Tian L H,Peng X Q.Effect of seeding rate on production performance and photosynthetic characteristics ofAvenasativacv. LENA in alpine pastoral regions.Pratacultural Science,2017,34(4):761-771.(in Chinese)
[20] 郑曦,魏臻武,武自念,李伟民,陈斐,刘倩,潘玲.不同燕麦品种(系)在扬州地区的适应性评价.草地学报,2013,21(2):272-279.
Zheng X,Wei Z W,Wu Z N,Li W M,Chen F,Liu Q,Pan L.Adaptability evaluation of differentAvenasativavarieties in Yangzhou area.Acta Agrestia Sinica,2013,21(2):272-279.(in Chinese)
[21] 周青平,颜红波,梁国玲,贾志峰,刘文辉,田莉华,陈有军,陈仕勇.不同燕麦品种饲草和籽粒生产性能分析.草业学报,2015,24(10):120-130.
Zhou Q P,Yan H B,Liang G L,Jia Z F,Liu W H,Tian L H,Chen Y J,Chen S Y.Analysis of the forage and grain productivity of oat cultivars.Acta Prataculturae Sinica,2015,24(10):120-130.(in Chinese)
[22] 曲祥春,何中国,郝文媛,栾天浩,李玉发.我国燕麦生产现状及发展对策.杂粮作物,2006,26(3):233-235.
Qu X C,He Z G,Hao W Y,Luan T H,Li Y F.Productive status and developing countermeasure of oat in China.Rain Fed Crops,2006,26(3):233-235.(in Chinese)
[23] Shewry P R,Piironen V,Lampi A M,Nystöm L,Li L,Rakszegi M,Delcour J A.Phytochemical and fiber components in oat varieties in the HEALTHGRAIN diversity screen.Journal of Agricultural and Food Chemistry,2008,56(21):9777-9784.
[24] Cognat C,Shepherd T,Verrall S R,Stewart D.Comparison of two headspace sampling techniques for the analysis of off-flavour volatiles from oat based products.Food Chemistry,2012,134(3):1592-1600.
[25] Heini R L,Oksman-Caldentey K M,Latva-Kala K,Lehtinen P,Poutanen K.Effect of drying treatment conditions on sensory profile of germinated oat.Cereal Chemistry,2001,78(6):707-714.
[26] 刘建新,王金成,贾海燕.燕麦幼苗对盐胁迫和碱胁迫的生理响应差异.水土保持学报,2015,29(5):331-336.
Liu J X,Wang J C,Jia H Y.Different between physiological responses ofAvenanudaseedling to salt and alkali stresses.Journal of Soil and Water Conservation,2015,29(5):331-336.(in Chinese)
[27] 夏方山,李晓禹,董秋丽,王明亚,朱慧森,杜利霞,佟莉蓉.抗坏血酸引发对NaCl胁迫燕麦幼苗生长的影响.山西农业大学学报(自然科学版),2017,37(11):761-766.
Xia F S,Li X Y,Dong Q L,Wang M Y,Zhu H S,Du L X,Tong L R.Effects of ascorbic acid priming on seedling growth of oat seeds under NaCl stress.Journal of Shanxi Agricultural University(Natural Science Edition), 2017,37(11):761-766.(in Chinese)
[28] ISTA.International Rules for Seed Testing.Bassersdorf:Zurich,2015.
[29] 卢培娜,刘景辉,李倩,申逸杰,严威凯,薛国兴.盐碱地不同燕麦品种的品质及产量比较.麦类作物学报,2016,36(11),1510-1516.
Lu P N,Liu J H,Li Q,Shen Y J,Yan W K,Xue G X.Comparison of quality and yield of different oat varieties in saline-alkali land.Journal of Triticeae Crops,2016,36(11):1510-1516.(in Chinese)
[30] 刘建新,王金成,王瑞娟,刘秀丽.混合盐碱胁迫对燕麦幼苗矿质离子吸收和光合特性的影响.干旱地区农业研究,2017,35(1):178-184,239.
Liu J X,Wang J C,Wang R J,Liu X L.Effect of complex saline-alkali stress on the mineral ions absorption and photosynthetic characteristics of oat seedlings.Agricultural Research in the Arid Areas,2017,35(1):178-184,239.(in Chinese)
[31] 刘建新,王金成,王瑞娟,贾海燕.燕麦幼苗对氯化钠和氯化钾胁迫的生理响应差异.水土保持学报,2014,34(5):74-75.
Liu J X,Wang J C,Wang R J,Jia H Y.Differences in physiological responses ofAvenanudaseedlings to NaCl and KCl stress.Bulletin of Soil and Water Conservation,2014,34(5):74-75.(in Chinese)
[32] 刘拥海,俞乐,王若仲,彭新湘,萧浪涛.抗坏血酸对植物生长发育的作用及其缺失突变体的研究进展.植物生理学报,2011,47(9):847-854.
Liu Y H,Yu L,Wang R Z,Peng X X,Xiao L T.Role of ascorbic acid in plant growth and development and its deficient mutants in higher plants.Plant Physiology Journal,2011,47(9):847-854.(in Chinese)
[33] 张启雷,刘强,高辉,陆俐娜,彭长连.内源抗坏血酸对水稻种子萌发及幼苗生长的影响.热带亚热带植物学报,2016,24(3):273-279.
Zhang Q L,Liu Q,Gao H,Lu L N,Peng C L.Effect of endogenous ascorbic acid on seed germination and seedling growth of rice.Journal of Tropical and Subtropical Botany,2016,24(3):273-279.(in Chinese)
[34] Gallie D R.The role of L-ascorbic acid recycling in responding to environmental stress and in promoting plant growth.Journal of Experiment Botany,2013,64(2):433-443.
[35] 刘艳军,范晶,韩学珅,李斌,张义.外源维生素对高NaCl胁迫下紫花苜蓿种子萌发及恢复性的影响.中国农学通报,2015,31(26):12-27.
Liu Y J,Fan J,Han X K,Li B,Zhang Y.Effect of exogenous vitamins on seed germination and recovery ofMedicagosativaL. under high NaCl stress.Chinese Agricultaural Science Bulletin,2015,31(26):12-27.(in Chinese)
[36] 范美华,张义鑫,石戈,崔大练,李鹏.外源抗坏血酸对油菜种子在海水胁迫下萌发生长的影响.中国油料作物学报,2009,31(1):34-38.
Fan M H,Zhang Y X,Shi Y,Cui D L,Li P.Effects of exogenous ascorbic acid on seed germination and growth ofBrassicanapusunder seawater stress.Chinese Journal of Oil Crop Science,2009,31(1):34-38.(in Chinese)
[37] 江绪文,李贺勤,王建华.盐胁迫下黄芩种子萌发及幼苗对外源抗坏血酸的生理响应.植物生理学报,2015,51(2):166-170.
Jiang X W,Li H Q,Wang J H.Physiological response ofScutellariabaicalensisseed germination and seedling to exogenous ascorbic acid under salt stress.Plant Physiology Journal,2015,51(2):166-170.(in Chinese)
[38] 石永春,杨永银,薛瑞丽,刘巧真.植物中抗坏血酸的生物学功能研究进展.植物生理学报,2015,51(1):1-8.
Shi Y C,Yang Y Y,Xue R L,Liu Q Z.Research advance of biological function of ascorbic acid in plants.Plant Physiology Journal,2015,51(1):1-8.(in Chinese)
