玉米/大豆套作体系中大豆茎秆生长过程模拟
2018-03-21杜勇利官思成邓瑜川刘卫国杨文钰四川农业大学农学院农业部西南作物生理生态与耕作重点实验室四川省作物带状复合种植工程技术研究中心四川成都630南充市农业科学院四川南充637000
杜勇利,周 涛,杨 欢,刘 婷,官思成,邓瑜川,刘卫国,杨文钰(.四川农业大学农学院/农业部西南作物生理生态与耕作重点实验室/四川省作物带状复合种植工程技术研究中心,四川 成都 630; .南充市农业科学院,四川 南充 637000)
大豆(Glycinemax)起源于中国,种植历史可以追溯到3 000年以前,除了作为重要的粮油作物外,其生物固氮、土壤磷活化、碳截获等生态意义越来越受到重视。以我国西南地区为例,土壤发育程度浅,有机质、速效氮、磷的含量低。但是受制于人口和粮食生产压力,该地区复种指数高,在旱地生态系统中,间/套作占据主导地位[1]。套作豆科植物相比单作能显著提高产量,主要体现在豆科植物活化土壤中难溶态磷、生物固氮供自身和共生作物吸收利用[2-4]。玉米(Zeamays)-大豆带状复合种植体系以其高产、生态等特点在我国西南地区得到大面积推广[5]。玉米/大豆套作模式不仅能使土壤水分、养分和光资源的供给得到改善,还可以增加这些资源的可利用性,产生互利,从而提高复种系数[6]。间、套作种植模式不仅遵循了时间镶嵌原则、空间扩展原则、种间搭配原则和物能流畅原则,而且还能解决用地和养地的矛盾、复种和季节不够用的矛盾,同时,作物茎秆还可解决发展畜牧业饲料不足的矛盾。
作物形态建成模拟是作物生长模拟中重要的一部分。明确作物外部形态变化规律,定量化研究作物形态建成过程,对指导作物生产具有重要的理论意义和实际应用价值。同时,也是虚拟作物和作物可视化的研究基础[7]。我国西南地区属于寡日照区域,在玉米和大豆共生期,受高位作物玉米遮阴导致苗期大豆受到一定程度的阴蔽胁迫,大豆植株茎长增加,茎粗、光合速率和地上部生物量显著下降,导致大豆易发生倒伏,从而影响大豆产量[8-9]。但是不同耐阴性大豆品种在套作环境中的茎秆生长特性呈现较大差异,强耐阴大豆品种茎秆抗倒伏能力明显强于一般或不耐阴品种[10]。因此,研究套作大豆的茎秆生长特性及不同品种茎秆在特殊光环境中的变化对于保障套作大豆产量和整个套作体系的稳产具有重要意义。
已有研究报道表明,以生理发育时间(physiological development time,PDT)为步长,可以对许多作物的形态生长动态进行模拟[11-13]。如,不同施氮水平和水分条件下水稻(Oryzasativa)的地上部器官形态特征建成[14-15];不同播期条件下大麦(Hordeumvulgare)茎秆、穗的生长动态[16];不同大豆品种茎、叶、荚的形态建成[17]。以生长日(growing degree days,GDD)为步长,Logistic方程可以很好的描述小麦叶片、穗、叶鞘和节间的动态生长变化[18-19]。同样,以扩充的Logistic模型可以对小麦(Triticumaestivum)株高和叶面积变化动态进行模拟[20]。此外,利用系统分析方法和动态建模技术,成功构建小麦叶片茎鞘夹角的动态模拟模型[21]。已有研究从不同角度对不同作物的生长发育规律进行了研究和模拟,通过模型模拟植物的生长动态技术得到广泛应用。虽然对单作大豆的形态建成动态模拟已有研究,但是套作大豆的生长环境与单作有较大差异,所以已有模型不能完全适用于描述套作大豆茎秆的生长动态。
因此,本研究以玉米/大豆带状套作复合种植模式中大豆为研究对象,通过对大豆茎秆生长过程的系统观测与分析,在定量研究的基础上,分别构建套作和单作条件下大豆节间伸长和增粗生长的动态模型,以及大豆茎秆干重变化动态模拟模型,旨为构建大豆在不同种植模式下生长决策支持系统奠定基础,也为实现大豆植株形态的虚拟显示、可视化研究提供关键技术。
1 材料与方法
1.1 供试材料
3个供试大豆品种株型间差异较大,分别为“南豆032-4”(无限型,弱耐阴性,四川省南充市农业科学院育种材料),株高120 cm左右,茎秆较细;“九月黄”(有限型,中度耐阴性,四川省阆中市地方品种),株高75 cm左右,茎秆粗细中等;“南豆12”(亚有限型,强耐阴性,四川省南充市农业科学院选育,西南地区套作大豆主推品种),株高90 cm左右,茎秆较粗。玉米选用半紧凑型品种“川单418”(四川农业大学玉米所选育)。
1.2 试验设计
试验于2012年和2013年在四川农业大学雅安校区教学科研园区(29°98′ N,103°00′ E)进行,属亚热带湿润季风气候区。年均温16.2 ℃,年日照时数1 005~1 200 h,年降水量1 250~1 750 mm;土壤类型为紫色土,质地为重壤土。pH 7.4,有机质含量为27.86 g·kg-1,全氮含量为1.79 g·kg-1,全磷含量为0.89 g·kg-1,全钾含量为29.76 g·kg-1,速效氮含量为115 mg·kg-1,速效磷含量为37 mg·kg-1,速效钾含量为136 mg·kg-1。田间采用随机区组排列,每个区组内包含3个玉米/大豆套作处理(3个大豆品种)和3个大豆单作处理,小区面积24 m2,重复3次。套作模式采用宽窄行种植,玉米窄行行距40 cm,宽行行距160 cm,穴距35 cm,每穴2株,分别于当年4月初播种玉米。大豆于玉米宽行间播种两行,窄行行距为50 cm,穴距10 cm,穴留2株,玉米/大豆间距55 cm,分别于2012年6月22日和2013年6月23日播种。单作模式中,大豆行距50 cm,穴距10 cm,每穴定苗1株。氮(N)、磷(P2O5)、钾(K2O)用量分别为195、75和105 kg·hm-2;玉米于每年4月初挖窝点播,窝深7 cm左右,宽15 cm左右,将30%的氮和全部的磷、钾肥施于窝的一侧,另一侧放种子避免种子与肥料接触,而后覆土,种子覆土深度不超过5 cm;再于玉米拔节期和大喇叭口期分别追施30%、40%的氮,均采用兑清水冲施于株旁的方式。大豆不施肥, 田间杂草病虫害管理方式同当地农户高产田。
1.3 观测项目及建模方法
由于大豆第1节和第2节的叶片分别为子叶、真叶,从第3节间才开始长出复叶。因前两个节均不会开花结荚,且节间长度较短,同时品种间差异不大,所以本研究中所有指标的模拟值和观测值均从第3节间开始。从大豆第3节始,每小区选长势一致的大豆植株挂牌标记,测定以下指标。
1.3.1茎秆生长的观测 每隔3 d定点定株测量已标记大豆茎秆第3节及其以上各节的节间长度和粗度。节间长度使用米尺测量,粗度使用游标卡尺测量,其测量部位为节间中部。
1.3.2茎干重测定 自播种大豆后7 d (即套作和单作大豆生育期都为VE)开始调查取样。每小区选取生长一致的300株挂牌标记。每隔7 d取样,直至大豆收获。每次选取10株从茎秆第1节剪断,单株按叶片、茎分解植株,然后分别装入牛皮纸袋中, 置于烘箱中105 ℃杀青30 min,再于75 ℃下烘干至恒重,最后称重,计算干重。
1.3.3数据处理及建模 本研究以生理发育日(PDT)为步长,运用Richards方程构建大豆茎长、茎粗生长的动态模型,获取方程特征参数α、β、γ、δ,参数α为增长量的极限值、β为初始值参数、γ为增长速率参数、δ为曲线形状参数。方程模拟图形是以α为渐近线的S形曲线。以作物生长发育实际天数为步长,运用Logistic方程构建大豆茎干重积累的动态模型,获取方程特征参数α、β、γ,参数α为增长量的极限值,β、γ均为方程特征参数。利用 Curve Expert 1.3 软件对单作和套作大豆的生长动态进行拟合,探明生理发育时间与大豆茎秆生长的关系,建立大豆生长动态模型所需的量化关系。大豆各发育阶段的生理发育时间的计算参照陈传梅等使用的方法[22]。并利用SPSS 17.0 软件分析这些特征参数。2012年试验数据用于模型构建和参数调试,2013年数据用于模型检验。采用根均方差RMSE来对模型进行检验,统计分析模拟值与观测值之间的符合情况。RMSE值越小,说明模拟值与观测值的一致性越好,模型的模拟结果越精确、可靠。RMSE的计算公式如下:
式中:OBSi为观测值,SIMi为模拟值,n为样本容量。优化不同大豆品种在不同栽培模式下,建立具有生物学意义的动态共性模拟方程,同时使用Excel分析软件进行回归分析。
2 结果与分析
2.1 品种遗传参数的确定
3个供试大豆品种的茎秆节间潜在长度、茎秆潜在粗度和茎秆潜在干重如表1所列。3个供试大豆品种的遗传参数,品种间和不同种植模式下都表现出了差异。且在套作种植模式下品种间的节间潜在长度、潜在粗度和茎的潜在干重均表现出显著差异性;单作种植模式下,品种间的遗传参数也表现出差异,说明不同种植模式会改变品种的遗传参数。
2.2 模型构建
2.2.1大豆茎秆节间伸长的动态模型 试验结果表明,在套作模式下大豆各节间从开始伸长到定长需要16.53个生理日,各节间开始伸长的时间间隔为2.8个生理日。大豆第3节间开始伸长的生理发育时间为11.21个生理日。在单作模式下大豆各节间从开始伸长到定长需要14.88个生理日,各节间开始伸长的时间间隔为1.9个生理日。大豆第3节间开始伸长的生理发育时间为8.95个生理日。各节间伸长的动态方程分别如下:


式中:Ln(PDT)为大豆在某一生理发育时刻的第n节的节间长度(cm);Lmaxn为大豆第n节节间潜在长度(cm)。α、β、γ、δ均为模型参数(表2)。可以看出,套作和单作栽培模式下,α、β、γ、δ值变异均大。但是,3个大豆品种在套作和单作栽培模式下的方程,其相关系数均在0.99以上。α为每个品种潜在极限值,β值和γ值在栽培模式和不同大豆品种间变异均较大。δ值在单作栽培模式下变异较小,套作模式下变异较大。因此,需要通过对套作和单作栽培模式下的大豆分别建立共性模拟方程。
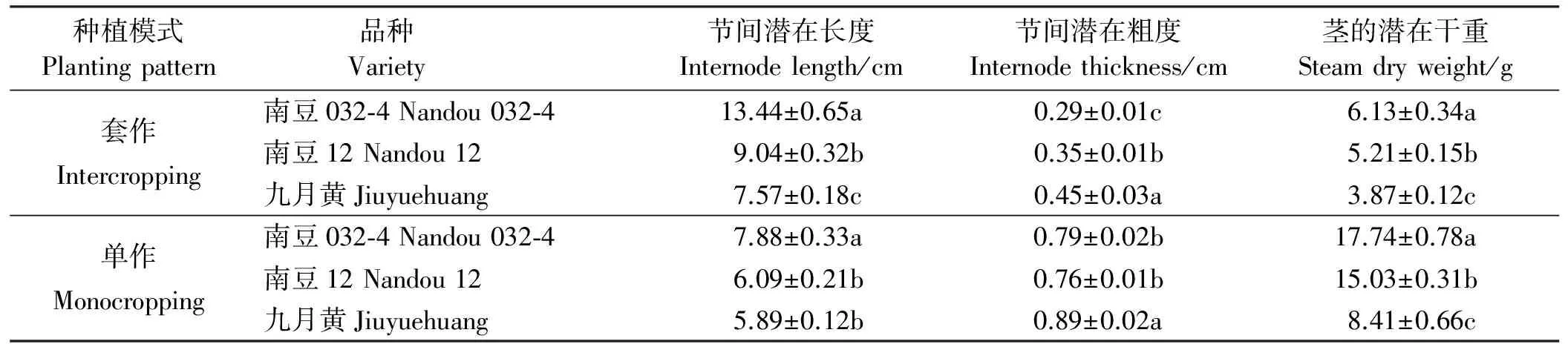
表1 单作、套作下的大豆节间潜在长度、潜在粗度和茎的潜在干重Table 1 Genetic coefficients of the three varieties of soybean
同列不同字母表示同处理不同品种间差异显著(P<0.05)。
Different lowercase letters indicate significant difference for the same planting pattern among different varietiesy at the 0.05 level.
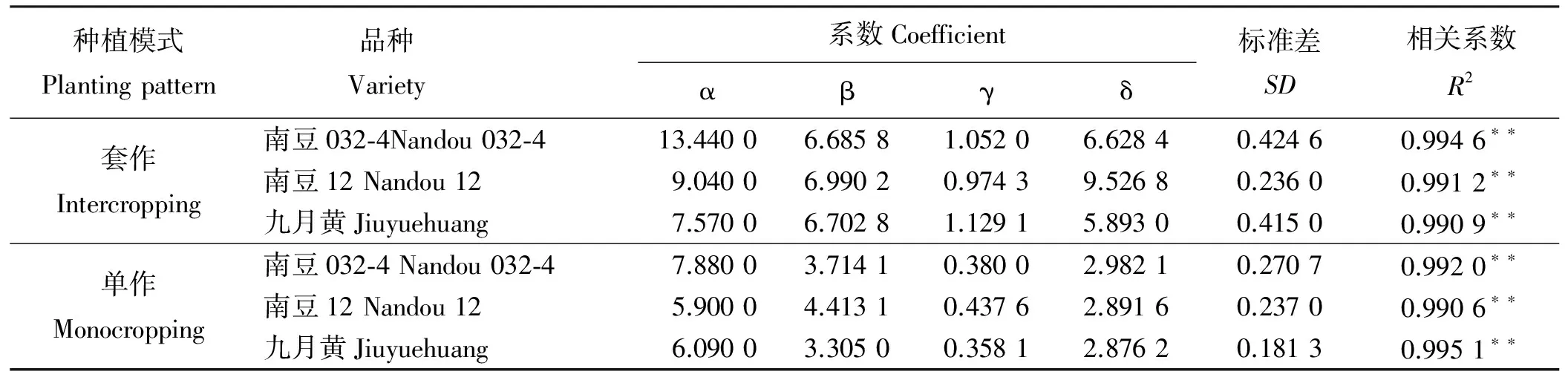
表2 单作、套作下不同大豆品种相对节间长度的方程系数Table 2 Coefficients of normalized internode lengths calculated using the dynamic equations in three varieties of soybean in intercropping and monocropping practices
**表示P<0.01;α为相对最大生长量;β为参数初值;γ为生长速率参数;δ为形状参数。下同。
**,P<0.01; α, denoted relative maximum increment; β, denoted initial value of parameter; γ,denoted growth rate parameters;δ, denoted shape parameter; similarly for the following tables.
套作模式下,随着大豆节间数的增加,其各节节间潜在长度Lmaxn逐渐增长,均在第7节间达到最大值,然后又逐渐下降(图1)。因此,在套作栽培模式下对南豆032-4、南豆12和九月黄的前7节茎秆节间长与PDT进行共性化处理,得到如下方程:
单作模式下,随着大豆节间数的增加,其各节节间潜在长度(Lmaxn)逐渐增长,不同品种大豆均在第12节间达到最大值,然后又逐渐下降(图1)。因此,在单作栽培模式下对南豆032-4、南豆12和九月黄的前12节茎秆节间长与PDT进行共性化处理,得到如下方程:
2.2.2节间增粗的动态模型 大豆节间增粗是从慢到快再到慢的过程。在套作模式下,大豆各节间从开始增粗到粗度固定需要14.88个生理日,各节间开始增粗的时间间隔为1.9个生理日。在单作模式下,大豆各节间从开始增粗到粗度固定需要16.53个生理日,各节间开始增粗的时间间隔为2.8个生理日。各节间增粗的动态方程如下:
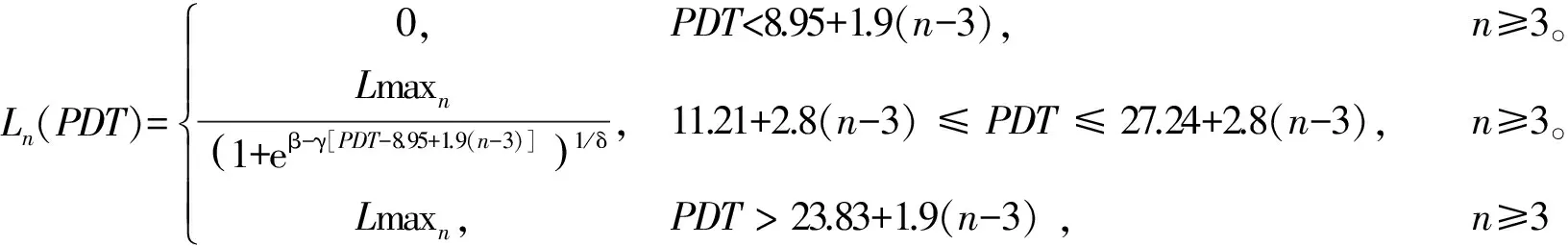

方程式中:Ln(PDT)为大豆在某一生理发育时刻的第n节的节间粗度(cm);PDT为生理发育时间,8.95为在套作中大豆第3节间开始增粗的生理发育时间,11.21为在单作中大豆第3节间开始增粗的生理发育时间。Lmaxn为大豆第n节节间的潜在粗度,是品种遗传参数。
可以看出(表3),3个大豆品种在不同栽培模式下的方程,其相关系数均在0.94以上,α为潜在极限值;β值和γ值在栽培模式和不同大豆品种间变异均较大。δ值在单作和套作栽培模式下变异均较大。

图1 单作、套作下各节间潜在长度随节间数的动态变化Fig. 1 Changes in internode length with respect to the number of internodes

表3 单作、套作下不同大豆品种相对节间粗度的方程系数Table 3 Coefficients of normalized internode thickness calculated using the dynamic equations in three varieties of soybean in intercropping and monocropping cultivation practices
套作模式下,随着大豆节间数的增加,其各节节间潜在粗度Lmaxn逐渐增加,在第5节间达到最大值,然后又逐渐下降(图2)。因此,在套作栽培模式下对南豆032-4、南豆12和九月黄的前5节茎秆节间长与PDT进行共性化处理,得到如下方程:
单作模式,随着大豆节间数的增加,其各节节间潜在粗度INLmaxn逐渐增加,在第7节间达到最大值,然后又逐渐下降(图2)。因此,在单作栽培模式下对南豆032-4、南豆12和九月黄的前7节茎秆节间长与PDT进行共性化处理,得到如下方程:
2.2.3大豆干物质量动态变化模型 大豆地上生物量干质量呈“缓慢生长-快速生长-缓慢生长”的生长过程。开始生长时较为缓慢,随着时间的推移(或温度的升高),在某一段时间内增长速度很快,当达到某一值后,增长速度又趋于缓慢,直至最后停止生长。这一特点符合 Logistic生长曲线。利用 Curve Expert 1.3软件对单作和套作大豆茎干重和生长发育实际天数之间的关系进行拟合,得到最优化的Logistic方程y=α/(1+βe-γx)],其中,y为干重,x为实际生长天数,获得方程特征参数α、β、γ(表4)。

图2 单作、套作下各节间潜在粗度随节间数的动态变化Fig. 2 Changes in internode thickness with respect to the number of internodes
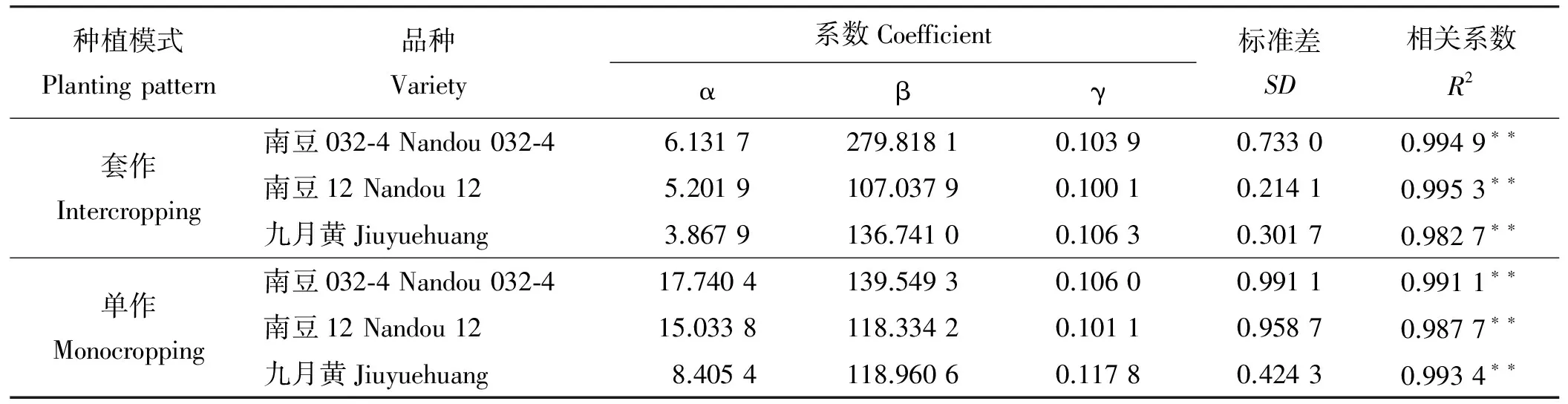
表4 单作、套作下不同大豆品种相对茎秆干重的方程系数Table 4 Coefficients of normalized dry weights of the stems calculated by dynamic equations in three soybean varieties in intercropping and monocropping cultivation practices
在不同栽培模式下对南豆032-4、南豆12和九月黄的茎秆干重与实际生长天数进行共性化处理。
套作种植模式下的方程:
式中:Wn(PDT)为套作大豆在某一实际生长时刻的茎干重(g);n为生长实际时间(d);通过动态观测套作大豆结果得出,Wmax77为大豆在第77天的时候大豆茎潜在干重达到最大(图3)。
单作种植模式下的方程:
式中:Wn(PDT)为单作大豆在某一实际生长时刻的茎干重(g);n为生长实际时间(d);通过动态观测单作大豆得出,Wmax75为大豆在第75天的时候大豆茎潜在干重达到最大(图3)。

图3 大豆茎潜在干重随生长时间的动态变化Fig. 3 Dynamic changes of potential stem dry weight of intercropping and monocropping soybean
2.3 模型的检验
通过2012-2013年的试验数据对套作和单作大豆茎秆动态模型进行检验。不同栽培模式下不同品种的大豆茎秆模拟值的RMSE结果(表5)表明,套作大豆节间长度、粗度、干重的模拟误差分别为0.24~0.33 cm、0.007~0.030 cm、0.21~0.37 g。单作大豆节间长度、粗度、干重的的模拟误差分别为0.11~0.17 cm、0.021~0.027 cm、0.46~0.54 g。说明套作模式下的模型和单作模式下的模型模拟值与观测值都表现出很好的一致性,模型的模拟结果精确、可靠。
采用RMSE分别对不同栽培模式下不同大豆节间长度、节间粗度、茎干重模型进行检验,对其模拟值与观测值分别做1∶1关系图(图4、5、6)。可以看出,不同栽培模式下不同大豆品种模拟值与实测值较接近,在套作模式下节间长度、节间粗度、茎干重模拟的准确度R2均在0.93以上,在单作模式下节间长度、节间粗度、茎干重模拟的准确度R2均在0.98以上。

表5 模型检验RMSE值Table 5 The root mean square errors of the validated model

图4 大豆不同节间长度的预测误差Fig. 4 Prediction errors for internode length in the different varieties of soybean
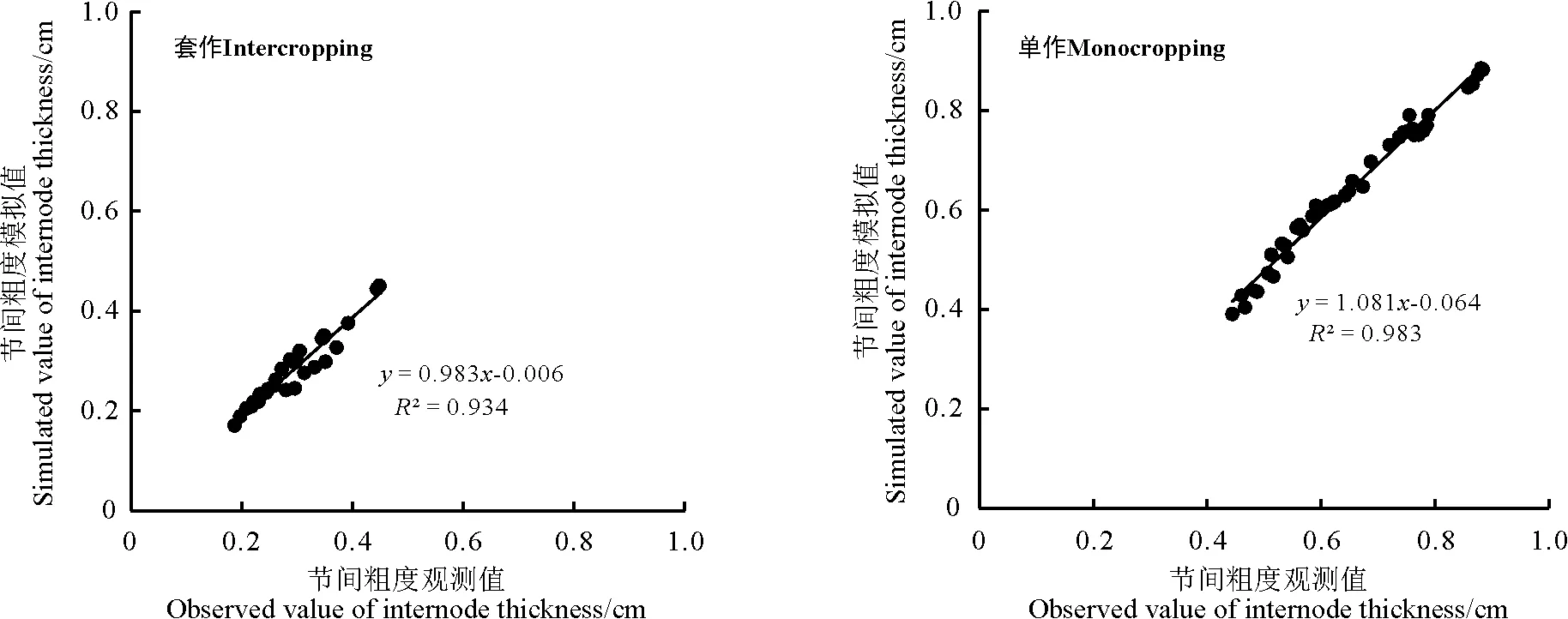
图5 大豆不同节间粗度的预测误差Fig. 5 Prediction errors for internode thickness in different varieties of soybean
3 讨论
3.1 不同大豆品种茎秆生长动态模型的建立
作物生长模型可以很好地描述作物生长发育的特征,如在水稻[23]、玉米[24]、大豆[25]等作物上均有相关研究报道。但是,对大豆生长模型的描述多侧重于单作大豆的全生育期,而对套作大豆模型的研究鲜有报道。单作模式下,大豆节间较短、较粗,保持较好的形态特征;与单作相比,套作大豆受高秆作物玉米遮阴的影响,光照不足,呈避阴反应,茎秆伸长变细;同一品种大豆在不同栽培模式下形态有差异明显。因此本研究分别对不同栽培模式大豆茎秆生长的动态模拟,通过量化各节间同步生长以及与生理发育时间的关系,以固定的节间潜在长度和潜在粗度来推算模型参数,以确定模型参数。以生理发育时间(PDT)为步长,建立了符合大豆茎秆和生理发育时间关系的Richards方程y=α/(1+βe-γx)1/δ,该方程能很好地模拟大豆茎秆随生理发育时间变化的动态特征,其方程参数有很好的生物学意义。当生理发育时间趋于无穷大时,不同栽培模式、品种的相对最大茎长、茎粗均趋于α,α值即为各品种的遗传参数,由品种自身决定;β值(初值参数)在栽培模式间变化幅度较大,在品种间变化幅度较小;γ值(生长速率参数)在栽培模式和品种间变化幅度均较大;δ值(形状参数)在不同栽培模式下变异很大。通常某一大豆品种对生理发育时间的需求是一定的, 同一品种在单作和套作环境下表现出不同的形态特征。因此通过调试方程中的β、γ、δ值,对Richards方程模型进行拟合,检验结果表明,在单作栽培模式下,节间长度、节间粗度观测值和模拟值的拟合较好(R2均大于0.983)。同样,在套作栽培模式下,节间长度、节间粗度观测值和模拟值的拟合较好(R2均大于0.934)。说明分别在不同品种及在单作、套作栽培模式下所建立的模型方程都表现出很好的拟合性,符合茎秆生长的生物学规律,具有一定的生物学意义。

图6 大豆不同生理发育时期茎干重的预测误差Fig. 6 Prediction errors of the dry weight of soybean stems over the physiological growth time
3.2 不同大豆品种茎秆干重动态模型的建立
大豆茎秆干重积累速率随着生长而变化,呈先上升后下降趋势,可分为渐增期、快增期和缓增期。前人的研究主要集中在大豆单作栽培模式下进行的模拟,根据Logistic方程分析不同耕作方式对夏大豆干物质积累与分配规律的影响[26];根据Logistic方程建立大豆干物质累积曲线,从而预报大豆产量[27]。然而,在带状套作复合种植模式下对大豆茎秆干重的模拟鲜见报道。本研究结果表明,不同的栽培模式,3个大豆品种的全生育期的茎秆干物质积累不同,套作大豆茎秆干重显著(P<0.05)低于单作大豆,由于光照不足亦导致大豆光合作用减弱,最终使大豆茎秆干物质积累降低[9]。因此分别对单作和套作栽培模式下进行模拟,以生长发育的实际天数为时间步长,通过Logistic方程的拟合,其在单作、套作栽培模式下的拟合效果较好(R2均大于0.979)。说明在单作、套作栽培模式下所建立的模型方程能很好的预测不同大豆品种生长发育实际天数与茎干重,能较好地拟合茎秆生长动态,符合茎秆生长的生物学规律,具有一定的生物学意义,将经验性和机理性有机地结合起来,具有较好的描述性和预测性。
4 结论
本研究采用单作、套作栽培模式,3种大豆在不同栽培模式下茎秆形态建成具有较大差异的材料,使模拟研究的可靠性更高。在系统观察的基础上,通过对这3种不同生长习性的大豆生长进行动态模拟,量化了各节间同步生长以及与生理发育时间的关系,根据观测结果,以固定的潜在节间长度和直径来推算其他模型参数。但本研究中由于仅考虑了栽培模式和品种对大豆茎秆生长的影响,并未深入研究养分、水分等外界因子的影响,所以需要进一步考虑温度、光照时间、养分等条件对大豆器官形态建成影响。今后将继续调试和优化模型,考虑水肥热等因素对大豆生育进程影响,使其在更多品种和更大地域范围内接受检验。
References:
[1] 杨文钰,杨峰,雍太文.我国间套作大豆研究方向和发展对策研讨会纪要.大豆科技,2011,27(1):35-36.
Yang W Y,Yang F,Yong T W.Symposium of research and development countermeasure of interplanting soybean in China.Soybean of Science and Technology,2011,27(1):35-36.(in Chinese)
[2] 雍太文,刘小明,刘文钰,苏本营,宋春,杨峰,王小春,杨文钰.减量施氮对玉米-大豆套作体系中作物产量及养分吸收利用的影响.应用生态学报,2014,25(2):474-482.
Yong T W,Liu X M,Liu W Y,Su B Y,Song C,Yang F,Wang X C,Yang W Y.Effects of reduced N application rate on yield and nutrient uptake and utilization in maize-soybean relay strip intercropping system.Chinese Journal of Applied Ecology,2014,25(2):474-482. (in Chinese)
[3] Li L,Li S M,Sun J H,Zhou L L,Bao X G,Zhang H G,Zhang F S.Diversity enhances agricultural productivity via rhizosphere phosphorus-deficient soils.Proceedings of the National Academy of Science of the United States of America,2007,104:11192-11196.
[4] 关佳威,姬明飞,王志强,樊哲翻,冉金枝,邓建明.不同种植密度条件下单混种作物的生长曲线.草业科学,2015,32(8):1243-1251.
Guan J W,Ji M F,Wang Z Q,Fan Z F,Ran J Z,Deng J M.The research of growth curve for monoculure and mixture planting crops grown under different density conditions.Pratacultural Science,2015,32(8):1243-1251.(in Chinese)
[5] 杨文钰,雍太文,任万军,樊高琼,牟锦毅,卢学兰.发展套作大豆,振兴大豆产业.大豆科学,2008(1):1-7.
Yang W Y,Yong T W,Ren W J,Fan G Q,Mu J Y,Lu X L.Develop relay planting soybean,revitalize soybean industry.Soybean of Science and Technology,2008,27(1):1-7.(in Chinese)
[6] 祁军,郑伟,张鲜花,唐高溶,王祥,朱进忠.不同豆禾混播模式的草地生产性能.草业科学,2016,33(1):116-128.
Qi J,Zheng W,Zhang X H,Tang G R,Wang X,Zhu J Z.Determination and comparison of the production performance of pastures among different spatial structure of legume-grass mixtures.Pratacultural Science,2016,33(1):116-128.(in Chinese)
[7] 徐寿军,李志刚,杨恒山,陈传梅,赵达,郭艳锋.大豆茎秆、叶片及豆荚生长的动态模拟.农业工程学报,2013,29(20):151-159.
Xu S J,Li Z G,Yang H S,Chen C M,Zhao D,Guo Y F.Dynamic simulation of stem,leaf and pod development on soybean.Transactions of the Chinese Society of Agricultural Engineering(Transactions of the CSAE),2013,29(20):151-159.(in Chinese)
[8] 邹俊林,刘卫国,袁晋,蒋涛,叶素琴,邓榆川,杨晨雨,罗玲,杨文钰.套作大豆苗期茎秆木质素合成与抗倒性的关系.作物学报, 2015,41(7):1098-1104.
Zou J L,Liu W G,Yuan J,Jiang T,Ye S Q,Deng Y C,Yang C Y,Luo L,Yang W Y.Relationship between lignin synthesis and lodging resistance at seedlings stage in soybean intercropping system.Acta Agronomica Sinica,2015,41(7):1098-1104. (in Chinese)
[9] 刘卫国,蒋涛,佘跃辉,杨峰,杨文钰.大豆苗期茎秆对荫蔽胁迫响应的生理机制初探.中国油料作物学报,2011,33:141-146.
Liu W G,Jian T,She Y H,Yang F,Yang W Y.Preliminary study on physiological response mechanism of soybean (Glycinemax) stem to shade stress at seedling stage.Chinese Journal of Oil Crop Sciences,2011,33(2):141-146.(in Chinese)
[10] 刘婷,刘卫国,任梦露,杜勇利,邓榆川,邹俊林,方萍,杨文钰.遮荫程度对不同耐荫性大豆品种光合及抗倒程度的影响.中国农业科学,2016,49(8):1466-1475.
Liu T,Liu W G,Ren M L,Du Y L,Deng Y C,Zou J L,Fang P,Yang W Y.Effects of shade degrees on photosynthesis and lodgingresistance degree of different shade tolerance soybean.Scientia Agricultura Sinica,2016,49(8):1466-1475.(in Chinese)
[11] Johnen T,Boettcher U,Kage H.A variable thermal time of the double ridge to flag leaf emergence phase improves the predictive quality of a CERES-Wheat type phenology model.Computers & Electronics in Agriculture,2012,89(5):62-69.
[12] Watanabe T,Hanan J S,Room P M.Rice morphogenesis and plant architecture:Measurement,specification and the reconstruction of structural development by 3D architectural modeling.Annals of Botany,2005,95(7):1131-1143.
[13] Hanan J S,Heam A B.Linking physiological and architectural models of cotton.Agricultural Systems,2003,75(1):47-77.
[14] 常丽英,顾东祥,张文宇,杨杰,曹卫星,朱艳.水稻叶片伸长过程的模拟模型.作物学报,2008,34(2):311-317.
Chang L Y,Gu D X,Zhang W Y,Yang J,Cao W X,Zhu Y.A simulation model of leaf elongation process in rice.Acta Agronomica Sinica,2008,34(2):311-317.(in Chinese)
[15] 石春林, 朱艳, 曹卫星. 水稻叶曲线特征的机理模型.作物学报,2006,32(5):656-660.
Shi C L,Zhu Y,Cao W X.A quantitative analysis on leaf curvature characteristics in rice.Acta Agronomica Sinica,2006,32(5):656-660.(in Chinese)
[16] 徐寿军,顾小莉,许如根,杨建香,席文娟,庄恒扬.大麦穗和茎秆生长的动态模拟.麦类作物学报,2007,27(2):282-287.
Xu S J,Gu X L,Xu R G,Yang J X,Xi W J,Zhuang H Y.Modeling spike and internode growth dynamics in barley.Journal of Triticeae Crops,2007,27(2):282-287.(in Chinese)
[17] 徐寿军,李志刚,杨恒山,陈传梅,赵达,郭艳锋.大豆茎秆、叶片及豆荚生长的动态模拟.农业工程学报,2013,29(20):151-159.
Xu S J,Li Z G,Yang H S,Chen C M,Zhao D,Guo Y F.Dynamic simulation of stem,leaf and pod development on soybean.Transactions of the Chinese Society of Agricultural Engineering(Transactions of the CSAE),2013,29(20):151-159.(in Chinese)
[18] 陈国庆,朱艳,曹卫星.冬小麦叶片生长特征的动态模拟.作物学报,2005,31(11):1524-1527.
Chen G Q,Zhu Y,Cao W X.Modeling leaf growth dynamics in winter wheat.Acta Agronomica Sinica,2005,31(11):1524-1527.(in Chinese)
[19] 谭子辉,朱艳,姚霞,田永超,刘小军,曹卫星.冬小麦麦穗生长过程的模拟研究.麦类作物学报,2006,26(4):93-97.
Tan Z H,Zhu Y,Yao X,Tian Y C,Liu X J,Cao W X.Modeling spike growth dynamics in winter wheat.Journal of Triticeae Crops,2006,26(4):93-97.(in Chinese)
[20] 王声锋,段爱旺,徐建新.冬小麦株高和叶面积指数变化动态分析及模拟模型.灌溉排水学报, 2010,29(4):97-100.
Wang S F,Duan A W,Xu J X.Dynamic changes and simulation model of plant height and leaf area index of winter wheat.Journal of Irrigation and Drainage,2010,29(4):97-100.(in Chinese)
[21] 张文宇,汤亮,朱相成,杨月,曹卫星,朱艳.基于过程的小麦茎鞘夹角动态模拟.应用生态学报,2011,22(7):1765-1770.
Zhang W Y,Tang L,Zhu X C,Yang Y,Cao W X,Zhu Y.Dynamic simulation of wheat stem sheath angle based on process.Chinese Journal of Applied Ecology,201l,22(7):1765-1770.(in Chinese)
[22] 陈传梅,赵达,李志刚,徐寿军,杨恒山,郭艳锋.大豆花芽分化和物候期的机理模型.中国油料作物学报,2012,34(5):502-507.
Chen C M,Zhao D,Li Z G,Xu S J,Yang H S,Guo Y F.Mechanism model for flower bud differentiation and phenophase of soybean.Chinese Journal of Oil Crop Sciences,2012,34(5):502-507.(in Chinese)
[23] 韦还和,孟天瑶,李超,张洪程,史天宇,马荣荣,王晓燕,杨筠文,戴其根,霍中洋,许轲,魏海燕,郭保卫.甬优籼粳杂交稻花后干物质积累模型与特征分析.作物学报,2016,42(2):265-277.
Wei H H,Meng T Y,Li C,Zhang H C,Shi T Y,Ma R R,Wang X Y,Yang J W,Dai Q G,Huo Z Y,Xu K,Wei H Y,Guo B W.Dynamic model and its characteristics analysis for dry matter production after heading of Indica/Japonica hybrid rice of yongyou series.Acta Agronomica Sinica,2016,42(2):265-277.(in Chinese)
[24] 李向岭,赵明,李从锋,葛均筑,侯海鹏,李琦,侯立白.播期和密度对玉米干物质积累动态的影响及其模型的建立.作物学报,2010,36(12):2143-2153.
Li X L,Zhao M,Li C F,Ge J Z,Hou H P,Li Q,Hou L B.Effect of sowing-date and planting density on dry matter accumulation dynamic and establishment of its simulated model in maize.Acta Agronomica Sinica,2010,36(12):2143-2153.(in Chinese)
[25] 杨萍,李杰,剡斌,牛俊义.胡麻/大豆间作体系下施氮对胡麻干物质积累和产量的影响.中国油料作物学报,2015(4):489-497.
Yang P,Li J,Yan B,Liu J Y.Effects of applied nitrogen on dry matter accumulation and oil flax yield in flax/soybean intercropping system.Chinese Journal of Oil Crop Sciences,2015(4):489-497.(in Chinese)
[26] 唐江华,苏丽丽,罗家祥,李亚杰,徐文修,彭姜龙.不同耕作方式对夏大豆干物质积累及转运特性的影响.核农学报,2015(10):2026-2032.
Tang J H,Su L L,Luo J X,Li Y J,Xu W X,Peng J L.Effects of different tillage methods on dry matter accumulation and transfer characteristics of summer soybean.Acta Agriculturae Nucleatae Sinica,2015(10):2026-2032.(in Chinese)
[27] 姜丽霞,那济海,朱海霞,韩俊杰,纪仰慧,闫平,王萍,王晨轶,李秀芬.基于Logistic方程的大豆产量预报方法.大豆科学,2008(3):414-419.
Jiang L X,Na J H,Zhu H X,Han J J,Ji Y H,Yai P,Wang P,Wang C Y,Li X F.A method to forecast yield of soybena which based on Logistic equation.Soybean of Science and Technology,2008(3):414-419.(in Chinese)
