高原鼠兔干扰对高山嵩草繁殖分配的影响
2018-03-21张红艳庞晓攀金少红郭正刚草地农业生态系统国家重点实验室兰州大学草地农业科技学院甘肃兰州730020草业科学国家级实验教学示范中心兰州大学甘肃兰州730020
张红艳,王 倩,于 成,庞晓攀,金少红,郭正刚(1.草地农业生态系统国家重点实验室,兰州大学草地农业科技学院,甘肃 兰州 730020;2.草业科学国家级实验教学示范中心(兰州大学),甘肃 兰州 730020)
高山嵩草(Kobresiapygmaea)草甸是青藏高原高寒草甸的主要类型,具有维持高原特色畜牧业生产、生物多样性保育、水源涵养及水土流失控制等重要功能[1-2]。高山嵩草是高山嵩草草甸的优势种,也是优质牧草[3-4],更是高山嵩草草甸植物群落的维稳者。高山嵩草的繁殖更新是高山嵩草草甸持久存在的基础,因此研究高山嵩草的繁殖特征对维系高山嵩草草甸的健康具有重要意义。高山嵩草的繁殖主要以营养繁殖为主[5],但往往受外界干扰而改变。外界干扰影响高山嵩草的繁殖特征,往往影响高山嵩草在植物群落内的更新能力,进而影响高山嵩草草甸的功能。
高原鼠兔是青藏高原特有的、常见的、营社会生活的小型草食哺乳动物。高山嵩草草甸相对低矮,视野开阔,往往成为高原鼠兔的主要栖息地[6]。高原鼠兔往往通过直接和间接的方式对高山嵩草草甸产生广泛的影响[7],一方面通过少量采食影响高山嵩草生长,另一方面通过改变高山嵩草生存的环境和资源状况影响高山嵩草的繁殖特征,从而影响高山嵩草的更新能力和草甸功能的发挥。已有研究表明,高原鼠兔干扰明显影响了高山嵩草草甸内达乌里秦艽(Gentianadahurica)的繁殖特征[8],较低干扰强度促进达乌里秦艽的同化物质向同化器官分配,较高干扰强度则促进达乌里秦艽的同化物质向生殖器官分配,然而达乌里秦艽的繁殖主要以有性繁殖为主,但高原鼠兔干扰对营养繁殖为主的植物繁殖特征是否具有同样的影响,尚需科学试验提供证据。
植物繁殖分配是植物繁殖特征的核心内容,是植物适应变化生境的主要特征之一[9-10]。植物繁殖分配主要包括两个方面的内容,首先是同化物质在地上和地下间的分配倾向,其次是同化物质在营养和生殖之间的分配倾向,以此实现植物种群在生存与生殖之间的权衡[8]。已有研究表明,狼毒(Stellerachamaejasme)种群随着分布海拔高度的增加,往往通过调整其地上地下生物量的分配适应环境变化[11];放牧干扰迫使冷蒿(Artemisiafrigida)种群调整其在营养和生殖生长之间的生物量分配倾向适应环境变化[12]。高原鼠兔干扰必然引起高山嵩草草甸内各种自然资源的重新配置,而资源的重新配置通常会促使植物个体将所获得的资源进行优化分配,以增加其适合度来维持生长繁殖[13-14]。因此,研究高原鼠兔干扰对高山嵩草繁殖分配的影响,是揭示高山嵩草繁殖特征响应高原鼠兔干扰的主要内容,更是明确高原鼠兔干扰是否影响无性繁殖植物的繁殖特征的需求。本研究通过调查高原鼠兔干扰和未干扰区,以及干扰区内不同干扰强度下高山嵩草同化物质的繁殖分配比例,在一定程度上阐明高山嵩草繁殖分配对高原鼠兔干扰的响应,回答高原鼠兔干扰是否影响营养繁殖植物的繁殖特征的科学问题,为揭示高原鼠兔干扰对高山嵩草生长的影响提供基础资料。
1 研究地区和方法
1.1 研究地区概况
研究地区位于青藏高原东部,行政隶属于甘肃省玛曲县境内,平均海拔4 053 m,属高寒大陆性气候。地理坐标为100°40′-102°29′ E,33°06′-34°30′ N,年均气温1.2 ℃,月平均温度变化较大,从1月份的-10 ℃到7月份的11.7 ℃。年相对无霜期仅19 d,无绝对无霜期。年均降水量为564 mm,年蒸发量为1 000~1 500 mm,蒸发量远大于降水量。植被类型主要是高寒草甸,其中高山嵩草草甸是主要草甸类型,该草甸类型以高山嵩草为优势种,以达乌里秦艽、小花草玉梅(Anemonerivularisvar.flore-minore)、垂穗披碱草(Elymusnutans)、羊茅(Festucaovina)、矮藨草(Scirpuspumilus)、米口袋(Gueldenstaedtiaverna)等为主要伴生种。
1.2 研究方法
1.2.1试验设计 高原鼠兔营社会性生活,具有迁移特征,繁殖周期大约为3周,每胎产仔2~7只[15],采用直接密度估测其干扰程度十分困难,因此经常采用有效洞穴密度测度其干扰程度[7]。2016年7月底在玛曲县阿孜站围栏的冷季放牧草地(夏季不放牧)内,选定地势相对平坦且一致的高山嵩草草甸为取样区,在取样区内,若在草地内同时发现高原鼠兔洞口和高原鼠兔,则将其设定为干扰区,若草地内没有高原鼠兔洞口(或很少,指1 hm2内洞口密度不超过1个)和没有发现高原鼠兔,则设定为非干扰区,干扰区与非干扰区草甸类型和地形地貌保持一致[16]。每随机选择一个干扰区样地,则在最近距离处选择一个非干扰区,总计选择了10个干扰区样地和10个非干扰区样地,样地面积为25 m×25 m,样地间隔大于25 m,然后用木桩四角标记,干扰区样地和非干扰区样地整体在研究区内呈现为交错分布状态。在干扰区内,采用连续3 d堵洞法测定样地内的有效洞穴密度[17-19],取3 d测的有效洞口数的平均值作为该观测样地的高原鼠兔干扰水平,其分别为304、384、432、512、576、752、864、928、1 040和1 216个·hm-2。然后在20个样地内,均采用“W”型布设5个1 m×1 m的样方,样方间隔8 m。每个样方内,采取挖根法采集全部高山嵩草的完整植株,装入信封并置于冷藏盒中带回实验室。实验室内,将每个样方内所收获的全部植株从茎基部将根系剪下,地上部分分离成生殖枝、营养枝与剩余构件三部分分别装入信封袋,地下部分根系放入0.5 mm筛子,用流水冲洗干净并装入信封袋,所有样品均置于60 ℃烘箱中烘至恒重后分别称量其干重。地上生物量为生殖枝、营养枝及剩余植株构件生物量的总和,总生物量是地上和地下生物量的和,生殖枝与营养枝比是指生殖枝与营养枝生物量比值,根冠比指地下生物量和地上生物量比值。
1.2.2数据分析 高原鼠兔对各指标的影响是否存在显著差异采用独立样本的T检验,若差异不显著,则不进行深入分析;若差异显著,则对植物不同部分生物量和干扰强度间的关系采用R-3.3.2-windows进行相关性检验。
2 结果
2.1 高原鼠兔干扰对高山嵩草总生物量、地上生物量及地下生物量的影响
高原鼠兔干扰虽然明显影响了高山嵩草总生物量、地上生物量和地下生物量,但对3种生物量的影响结果不一致(图1)。高原鼠兔干扰显著提高了高山嵩草总生物量和地下生物量(P<0.05),但降低了地上生物量(P<0.05)。高山嵩草地上生物量与总生物量相关性不显著(P>0.05),但地下生物量与总生物量极显著正相关(P<0.01)(表1)。高原鼠兔干扰区内,高山嵩草总生物量、地上生物量和地下生物量随高原鼠兔干扰程度增加表现出不一样的变化趋势(图2),具体体现为高山嵩草地下生物量随干扰梯度增加没有明显的变化规律,而总生物量和地上生物量均随干扰梯度增加表现为单峰曲线。

表1 高原鼠兔干扰与未干扰下高山嵩草地上生物量、地下生物量与总生物量的相关性分析Table 1 Correlation coefficient between total biomass, aboveground biomass and underground biomass of Kobresia pygmaea

图1 高原鼠兔干扰对高山嵩草总生物量、地上生物量及地下生物量的影响Fig. 1 Effects of plateau pika disturbance on total biomass, aboveground biomass and underground biomass of Kobresia pygmaea

图2 干扰区不同干扰水平下高山嵩草总生物量、地上生物量及地下生物量的变化Fig. 2 The total biomass, aboveground biomass and underground biomass of Kobresia pygmaea under different disturbance levels of plateau pika
图中灰色部分代表95%的置信区间。
An adjusted local smoothed regression line (black) is shown with its 95% confidence interval in gray.
2.2 高原鼠兔干扰对高山嵩草根冠比的影响
高原鼠兔干扰显著增加了高山嵩草的根冠比(P<0.05)(图3)。高原鼠兔干扰区内,高山嵩草的根冠比随高原鼠兔干扰程度增加依然没有明显的变化规律(图4)。
2.3 高原鼠兔干扰对高山嵩草生殖枝与营养枝比的影响
高原鼠兔干扰显著增加了生殖枝与营养枝比(P<0.05)(图5)。高原鼠兔干扰区内,高山嵩草生殖枝与营养枝比随干扰程度增加而表现为先降低后增加的态势(图6)。

图3 高原鼠兔干扰对高山嵩草根冠比的影响Fig. 3 Effect of plateau pika disturbance on root-shoot ratio of Kobresia pygmaea

图4 高原鼠兔干扰区不同干扰水平下高山嵩草根冠比的变化Fig. 4 Root-shoot ratio of Kobresia pygmaea under different disturbance levels of plateau pika
3 讨论
繁殖分配指植物分配给繁殖活动的各种资源,是植物生长、生殖、贮藏或防卫等主要功能间资源需求权衡的结果[20]。高原鼠兔干扰行为一定程度上改变了植物利用生境内资源的能力,因此,植物繁殖分配必将对各种资源做出重新配置[8],优化各主要功能间的需求,将资源配置于植物最需求的功能。植物生物量积累或分配是各种资源在植物体内积累或分配结果的具体量化反映,也是生态系统最基本的数量特征,更是植物适应环境条件能力和生长发育规律的综合反映[21]。本研究表明,高原鼠兔干扰明显降低了高山嵩草种群的地上生物量,这一方面可能与高原鼠兔干扰打破了原有植物群落间的物种密切关系,未干扰区高山嵩草为优势种,具有较强的竞争优势,而干扰区内高原鼠兔干扰形成了星罗棋布的裸斑,这些裸斑增加了地表蒸发,迫使草甸生境旱化,而旱生化环境释放了原来处于竞争弱势的中旱生植物如达乌里秦艽和莓叶委陵菜(Potentillafragarioides)的生长潜势,这客观上给高山嵩草的生长形成竞争压力,一定程度上抑制了高山嵩草的长势,从而降低了其地上生物量;另一方面可能是由于高原鼠兔对高山嵩草的采食与践踏等直接干扰活动,造成其地上生物量明显降低。虽然高原鼠兔干扰降低了高山嵩草的地上生物量,但其却显著增加了地下生物量,这归功于高原鼠兔干扰改变了植物利用土壤碳、氮能力和土壤物理性质,一方面,高原鼠兔干扰所形成的裸斑会产生微斑块效应,而这种微斑块效应往往促进植物对土壤碳、氮的利用率[22],另一方面,高原鼠兔干扰行为增加了土壤孔隙度[23],两个方面综合的结果导致植物根系累积营养物质的能力增加,且有利于植物根系生长,从而提高了地下生物量。地下生物量在高原鼠兔干扰下与总生物量呈正相关关系,这在一定程度上说明了根系在总生物量的构成中贡献较大,因此,高山嵩草地下生物量的增加导致了总生物量的增加。
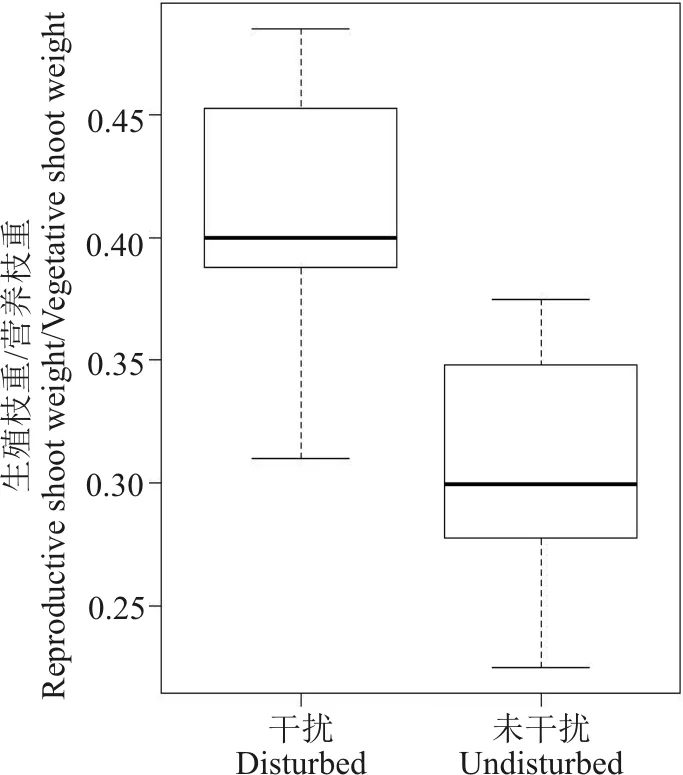
图5 高原鼠兔干扰对高山嵩草生殖枝重/营养枝重的影响Fig. 5 Effect of plateau pika disturbance on reproductive shoot-vegetative shoot biomass ratio of Kobresia pygmaea

图6 高原鼠兔干扰区不同干扰水平下高山嵩草生殖枝重/营养枝重的变化Fig. 6 The reproductive shoot-vegetative shoot biomass ratio of Kobresia pygmaea under different disturbance levels of plateau pika
图中灰色部分代表95%的置信区间。
An adjusted local smoothed regression line (black) is shown with its 95% confidence interval in gray.
根冠比是植物体生物量在地上和地下分配的直接体现[24],它的变化在一定程度上代表了根茎密丛型植物对环境的适应性应答[18]。高原鼠兔干扰显著增加了高山嵩草种群的根冠比,这与放牧干扰对冷蒿种群的根冠比影响一致[25],这可能有两个原因:首先高原鼠兔干扰后草甸生境异质化程度增加[26],高山嵩草的茎叶不能得到充分生长,此时高山嵩草种群将同化物质更多地投入到根系生长,增加高山嵩草的营养繁殖能力,旨在维持种群的稳定性和持续更新能力,这是高山嵩草长期适应高原鼠兔干扰的自然结果。其次,高原鼠兔的采食行为往往会诱发植物种群的规避性应答,由于植物体不能主动避免被采食,只能被动的通过调整自身物质分配,将更多资源转移至根系[27],以维持种群的生存力。
植物个体既要在生境中增强其适应性,保证亲代的存活,又要最大可能地产生种子延续后代,以确保完成其生活史,就会在营养生长和有性繁殖之间进行权衡[20]。而生殖枝和营养枝作为植物有性繁殖和营养生长能力的载体,在一定程度上能够代表植物对有性繁殖和营养生长的投入[28],二者比例的变化能够体现植物繁殖分配策略。高原鼠兔干扰明显增加了高山嵩草的生殖枝与营养枝比,这是由于高原鼠兔主要采食高山嵩草的营养器官,采食作用诱导植物的超补偿反应[29]会增加对生殖生长的投入,提高种群适应干扰环境的能力[30],一方面是由于植物种子存活的环境要求比根茎存活的环境要求相对较低,更有利于长期保持繁殖能力;另一方面,高原鼠兔干扰会导致生境发生不可预测性的变化,高山嵩草为保证其基因延续和存活无忧,将资源主要分配给生殖生长。
地上生物量及生殖枝与营养枝比不仅与高原鼠兔干扰有关,而且与干扰强度有关。其中地上生物量随着高原鼠兔干扰强度增加表现为先增加后降低态势,而生殖枝与营养枝比表现为先降低后增加态势,这说明上述指标响应高原鼠兔干扰时均存在合适阈值。当高原鼠兔干扰强度从低增加至合适阈值时,优势种高山嵩草占据有利竞争地位,能够利用较多的资源保持其生长优势[31-32],故地上生物量增加。但当高原鼠兔干扰强度超过合适阈值时,单位面积内的裸斑数随之增加,生境逐渐旱化,这使得高山嵩草与某些杂草之间存在较大的生态位宽度重叠[31],高山嵩草种群生长受到一定的限制,随着干扰强度增加,抑制效应愈加明显,表现为地上生物量逐渐降低。生殖枝与营养枝比随高原鼠兔干扰强度增加表现为先降低后增加态势,这是由于优势种高山嵩草在高原鼠兔干扰强度从低增加至合适阈值时,种群不断加大其在群落中的资源竞争能力,优势度逐渐增加[31,33],此过程中高山嵩草通过旺盛的营养生长占据较大的生境面积加速种群扩散,至合适阈值时,营养生长达到最高。然而,当高原鼠兔干扰强度超过合适阈值时,高山嵩草将大部分资源分配给生殖生长,一方面是因为过度干扰活动使得生境内可利用资源逐渐减少,土壤水分减少,较低的土壤水分影响了根茎芽的形成,进而导致较低的营养分配,而高山嵩草为保证其基因延续,对生殖生长投入更多资源;另一方面由于高山嵩草种群的优势地位下降,草地毒杂草急剧增加,它们释放的化学物质致使草食动物不愿接近采食,反而对高山嵩草生殖枝的产生起到了某种保护作用[5,34],从而表现为生殖枝与营养枝比增加。这与退化程度越大越不利于高山嵩草营养生长的研究结果一致[5]。
综合上述,高原鼠兔干扰虽然对无性繁殖高山嵩草和有性繁殖达乌里秦艽均产生一定的影响,但对两种植物的影响是不一样的,达乌里秦艽随干扰强度增加生长潜势越大[8],而高山嵩草生长潜势只有干扰强度在合理阈值时才得到释放。
4 结论
高原鼠兔干扰显著增加高山嵩草种群总生物量、地下生物量,生殖枝和营养枝比和根冠比,说明高山嵩草通过调整繁殖分配而适应高原鼠兔干扰后的生境,要么提高种群扩张力,要么提高种群生存力。高山嵩草在不同干扰强度下总生物量、地上生物量和生殖枝与营养枝比表现出的差异显示了其对异质环境较强的适应性,但三者均存在一个合理的干扰阈值,干扰强度在此阈值以下时,有利于高山嵩草生长,其总生物量与地上生物量达到最大值,主要以营养生长为主,而干扰强度超过阈值时,高山嵩草生长受到抑制,其总生物量与地上生物量逐渐下降,主要以生殖生长为主,这说明适宜的高原鼠兔干扰能够增强以营养繁殖为主的高山嵩草的资源获取能力,而当高原鼠兔过度干扰时,其资源竞争能力下降。
References:
[1] 王启基,李世雄,王文颖,景增春.江河源区高山嵩草(Kobresiapygmaea)草甸植物和土壤碳、氮储量对覆被变化的响应.生态学报,2008,28(3):885-894.
Wang Q J,Li SX,Wang W Y,Jing Z C.The despondences of carbon and nitrogen reserves in plants and soils to vegetations cover change onKobresiapygmaeameadow of Yellow River and Yangtze River source region.Acta Ecologica Sinica,2008,28(3):885-894.(in Chinese)
[2] Wu R X,Chai Q,Zhang J Q,Zhong M Y,Liu Y H,Wei X T,Pan D,Shao X Q.Impacts of burrows and mounds formed by plateau rodents on plant species diversity on the Qinghai-Tibetan Plateau.Rangeland Journal,2015,37(1):117-123.
[3] 李雄,胡向阳,杨永平.青藏高原重要牧草高山嵩草的研究现状.草业与畜牧,2013(1):30-39.
Li X,Hu X Y,Yang Y P.Research of important forageKobresiapygmaeain Qinghai-Tibet Plateau.Prataculture and Animal Husbandry,2013(1):30-39.(in Chinese)
[4] 周兴民,王启基,师生波.中国嵩草草甸.北京:科学出版社,2001.
Zhou X M,Wang Q J,Shi S B.AlpineKobresiaMeadow in China.Beijing:Science Press,2001.(in Chinese)
[5] 杨元武,李希来.不同退化程度高寒草甸高山嵩草的构件变化.西北植物学报,2011,31(1):167-171.
Yang Y W,Li X L.Modular variety ofKobresiapygmaeaon alpine meadow with different degraded levels.Acta Botanica Boreali-Occidentalia Sinica,2011,31(1):167-171.(in Chinese)
[6] 庞晓攀,王倩,贾婷婷,李倩倩,郭正刚.高原鼠兔有效洞口数密度对高山嵩草草甸植物种间联结性的影响.草业学报,2015,24(5):224-230.
Pang X P,Wang Q,Jia T T,Li Q Q,Guo Z G.Effect of burrow entrance densities of plateau pika (Ochotonacurzoniae) on interspecific association inKobresiapygmaeameadow.Acta Prataculturae Sinica,2015,24(5):224-230.(in Chinese)
[7] Pang X P,Guo Z G.Plateau pika disturbances alter plant productivity and soil nutrients in alpine meadows of the Qinghai-Tibetan Plateau.The Rangeland Journal,2017,39(2):133-144.
[8] 王倩,贾婷婷,庞晓攀,金少红,陈传武,郭正刚.高原鼠兔干扰对达乌里秦艽繁殖特征的影响.草业科学,2015,32(5):796-801.
Wang Q,Jia T T,Pang X P,Jin S H,Chen C W,Guo Z G.Effects of disturbance of plateau pika (Ochotonacurzoniae) on reproductive characteristics ofGentianadahuricain the Qinghai-Tibetan Plateau.Pratacultural Science,2015,32(5):796-801.(in Chinese)
[9] Suter M.Reproductive allocation ofCarexflavareacts differently to competition and resources in a designed plant mixture of five species.Plant Ecology,2009,201(2):481-489.
[10] 牛克昌,赵志刚,罗燕江,杜国祯.施肥对高寒草甸植物群落组分种繁殖分配的影响.植物生态学报,2006,30(5):817-826.
Niu K C,Zhao Z G,Luo Y J,Du G Z.Fertilization effects on species reproductive allocation in an alpine meadow plant community.Journal of Plant Ecology,2006,30(5):817-826.(in Chinese)
[11] 张茜,赵成章,马小丽,侯兆疆,李钰.高寒草地狼毒种群繁殖分配对海拔的响应.生态学杂志,2013,32(2):247-252.
Zhang Q,Zhan C Z,Ma X L,Hou Z J,Li Y.Response of reproductive allocation ofStellerachamaejasmepopulation in alpine grassland to altitude.Chinese Journal of Ecology,2013,32(2):247-252.(in Chinese)
[12] 杜利霞,李青丰,董宽虎.放牧强度对短花针茅草原冷蒿繁殖特性的影响.草地学报,2007,15(4):367-370.
Du L X,Li Q F,Dong K H.Effects of grazing intensity on reproductive characteristics ofArtemisiafrigidaWilld. inStipabrevifloraGriseb. steppe.Acta Agrestia Sinica,2007,15(4):367-370.(in Chinese)
[13] 陈尚,马艳,李自珍,王刚.克隆植物种子繁殖和营养繁殖的适合度分析和度量.生态学报,1999,19(2):287-290.
Chen S,Ma Y,Li Z Z,Wang G.Analysis and measurement of the fitness of seed reproduction and vegetative reproduction of clonal plant.Acta Ecologica Sinica,1999,19(2):287-290.(in Chinese)
[14] 汤俊兵,肖燕,安树青.根茎克隆植物生态学研究进展.生态学报,2010,30(11):3028-3036.
Tang J B,Xiao Y,An S Q.Advance of studies on rhizomatous clonal plants ecology.Acta Ecologica Sinica,2010,30(11):3028-3036.(in Chinese)
[15] Smith A T.The World of Pikas.//Alves P C,Ferrand N,Hackländer K.Lagomorph Biology.Heidelberg,Berlin:Springer,2008:89-102.
[16] 金少红,刘彤,庞晓攀,于成,郭正刚.高原鼠兔干扰对青海湖流域高山嵩草草甸植物多样性及地上生物量的影响.草业学报,2017,26(5):29-39.
Jin S H,Liu T,Pang X P,Yu C,Guo Z G.Effects of plateau pika (Ochotonacurzoniae) disturbances on plant species diversity and aboveground plant biomass in aKobresiapygmaeameadow in the Qinghai Lake Region.Acta Prataculturae Sinica,2017,26(5):29-39.(in Chinese)
[17] Yu C,Pang X P,Wang Q,Jin S H,Shu C C,Guo Z G.Soil nutrient changes induced by the presence and intensity of plateau pika (Ochotonacurzoniae) disturbances in the Qinghai-Tibet Plateau,China.Ecological Engineering,2017,106:1-9.
[18] Sun F D,Chen W Y,Liu L,Liu W,Cai Y M,Smith P.Effects of plateau pika activities on seasonal plant biomass and soil properties in the alpine meadow ecosystems of the Tibetan Plateau.Grassland Science,2015,61(4):195-203.
[19] Guo Z G,Zhou X R,Hou Y.Effect of available burrow densities of plateau pika(Ochotonacurzoniae) on soil physicochemical property of the bare land and vegetation land in the Qinghai-Tibetan Plateau.Acta Ecologica Sinica,2012,32(2):104-110.
[20] 张林静,石云霞,潘晓玲.草本植物繁殖分配与海拔高度的相关分析.西北大学学报(自然科学版),2007,37(1):77-80.
Zhang L J,Shi Y X,Pan X L.Analysis of correlativity between reproductive allocation and altitude in plants.Journal of Northwest University(Natural Science Edition),2007,37(1):77-80.(in Chinese)
[21] 樊星,蔡捡,刘金平,伍德,宗人旭.草地植物群落组成对鹅绒委陵菜无性系拓展能力及生物量配置的影响.草业科学,2017,34(1):119-128.
Fan X,Cai J,Liu J P,Wu D,Zong R X.Effects of grassland plant communities on ramet expansion ability and biomass allocation ofPotentillaanserina.Pratacultural Science,2017,34(1):119-128.(in Chinese)
[22] 于成,贾婷婷,庞晓攀,郭正刚.高原鼠兔干扰强度对高寒草甸土壤碳氮分布的影响.土壤学报,2016,53(3):768-778.
Yu C,Jia T T,Pang X P,Guo Z G.Effects of plateau pika (OchotonaCurzoniae) disturbing soil carbon and nitrogen distribution in alpine meadow.Acta Pedologica Sinica,2016,53(3):768-778.(in Chinese)
[23] Canals R M,Herman D J,Firestone M K.How disturbance by fossorial mammals alters N cycling in a California annual grassland.Ecology,2008,84(4):875-881.
[24] 龙毅,孟凡栋,王常顺,白玲,钟扬,汪诗平.高寒草甸主要植物地上地下生物量分布及退化对根冠比和根系表面积的影响.广西植物,2015(4):532-538.
Long Y,Meng F D,Wang C S,Bai L,Zhong Y,Wang S P.Above- and below-ground biomass distribution of main alpine meadow plants and impact of degradation on root/shoot ratio and root area.Guihaia,2015(4):532-538.(in Chinese)
[25] 王静,杨持,王铁娟.放牧退化群落中冷蒿种群生物量资源分配的变化.应用生态学报,2005,16(12):2316-2320.
Wang J,Yang C,Wang T J.Changes of biomass allocation ofArtemisiafrigidapopulation in grazing induced retrogressive communities.Chinese Journal of Applied Ecology,2005,16(12):2316-2320.(in Chinese)
[26] Bagchi S,Namgail T,Ritchie M E.Small mammalian herbivores as mediators of plant community dynamics in the high-altitude arid rangelands of Trans-Himalaya.Biological Conservation,2006,127(4):438-442.
[27] 卫智军,闫瑞瑞,运向军,褚文彬,杨静.放牧制度下荒漠草原主要植物生物量及能量分配研究.中国沙漠,2011,31(5):1124-1130.
Wei Z J,Yan R R,Yun X J,Chu W B,Yang J.Study on biomass and energy allocation of major plant species in desert steppe under different grazing systems.Journal of Desert Research,2011,31(5):1124-1130.(in Chinese)
[28] 李金花,潘浩文,王刚.草地植物种群繁殖对策研究.西北植物学报,2004,24(2):352-355.
Li J H,Pan H W,Wang G.Reproductive strategy of grassland plant.Acta Botanica Boreali-Occidentalia Sinica,2004,24(2):352-355.(in Chinese)
[29] Olejniczak P.Overcompensation in response to simulated herbivory in the perennial herbSedummaximum.Plant Ecology,2011,212(11):1927-1935.
[30] Vanderweide B L,Hartnett D C.Belowground bud bank response to grazing under severe,short-term drought.Oecologia,2015,178(3):795-806.
[31] 贾婷婷,毛亮,郭正刚.高原鼠兔有效洞穴密度对青藏高原高寒草甸群落植物生态位的影响.生态学报,2014,34(4):1-9.
Jia T T,Mao L,Guo Z G.Effect of available burrow densities of plateau pika (Ochotonacurzoniae) on plant niche of alpine meadow communities in the Qinghai-Tibet Plateau.Acta Ecologica Sinica,2014,34(4):1-9.(in Chinese)
[32] 庞晓攀,贾婷婷,李倩倩,雒明伟,肖玉,赵旭,郭正刚.高原鼠兔有效洞穴密度对高山嵩草群落及其主要种群空间分布特征的影响.生态学报,2015,35(3):873-884.
Pang X P,Jia T T,Li Q Q,Luo M W,Xiao Y,Zhao X,Guo Z G.Effect of available burrow densities of plateau pika (Ochotonacurzoniae) on characteristics and distribution pattern ofKobresiapygmaeacommunity.Acta Ecologica Sinica,2015,35(3):873-884.(in Chinese)
[33] 王莹,庞晓攀,肖玉,贾婷婷,王倩,于成,郭正刚.高原鼠兔干扰对高寒草甸植物多样性与土壤养分间关系的影响.生态学报,2016,36(17):5485-5496.
Wang Y,Pang X P,Xiao Y,Jia T T,Wang Q,Yu C,Guo Z G.The effect of plateau pika (Ochotonacurzoniae) disturbance on the relationship between plant diversity and soil nutrients of alpine meadow.Acta Ecologica Sinica,2016,36(17):5485-5496.(in Chinese)
[34] 冶娜娜,张灵菲,卫万荣,王菊霞,张小刚,麻安卫,江小雷.啮齿动物应对植物次生代谢物的行为策略.草业科学,2015,32(2):281-286.
Ye N N,Zhang L F,Wei W R,Wang J X,Zhang X G,Ma A W,Jiang X L.The behavior strategy of herbivorous rodents against plant secondary metabolites.Pratacultural Science,2015,32(2):281-286.(in Chinese)
