早期胚胎停育患者绒毛组织中hsa-miR- 2276-3p及ITGA9、VEGF mRNA表达变化
2018-03-19张树红覃超喜孙燕
张树红,覃超喜,孙燕
(广西医科大学第一附属医院,南宁530022)
在妇产科领域,胚胎停育发生率约1%[1]。其原因众多,一部分原因归结于胚胎染色体异常、内分泌失调、免疫因素、子宫结构异常、感染、化学制剂和心理因素[2],但仍近50%的患者病因不明。因此,原因不明的胚胎停育是目前研究的热点[3]。成功的妊娠依赖于胎盘良好的发育和生长,在胎盘植入与胎盘形成的过程中,血管生成对于成功妊娠非常关键。有文献报道,胚胎发育过程中,胎盘血管功能不全可能是引起胚胎停育的原因之一[4]。研究发现,miRNA可参与胎盘血管生成的信号通路,影响胎盘血管的生成,介导早期的胚胎发育[5]。hsa-miR-2276-3p作为miRNA的一种,在血管内皮细胞中有特异性高表达。整合素9(ITGA9)作为细胞表面黏附分子介导细胞信号转导,在细胞分化、胚胎发育及细胞黏附等方面发挥重要作用;其配体很多,可与血管内皮生长因子(VEGF)相互作用。本研究检测了胚胎停育患者绒毛组织中miR-2276-3p及ITGA9、VEGF mRNA的表达情况,进而探讨三者的相关性,为胚胎停育发病机制的研究提供新思路。
1 资料与方法
1.1 临床资料 选取2017年11月~2018年2月在广西医科大学第一附属医院计划生育科就诊的胚胎停育和正常早孕行人工流产患者各30例,术中均留取绒毛组织。胚胎停育患者年龄(28.4±3.3)岁,孕龄49~65 d,均经B超确诊;正常早孕者年龄(27.7±3.8)岁,孕龄49~69 d。两者的既往月经均规律,月经周期(28±3)d;近3月未服用激素类药物,且排除生殖道解剖结构异常、内分泌异常、孕期感染、自身免疫性疾病等,无肝炎、结核、性病等传染病史,夫妻双方均无遗传病史。两者年龄及孕龄比较,差异均无统计学意义(P均>0.05)。本研究经医院医学伦理委员会批准,受试者均签署知情同意书。
1.2 绒毛组织中hsa-miR-2276-3p及ITGA9、VEGF mRNA检测方法 采用实时荧光定量PCR法。取100 mg绒毛组织,匀浆研磨,加入细胞裂解液裂解,TRIzol法提取组织总RNA。用NanoDrop2000紫外分光光度计测定RNA浓度和纯度,OD260/OD280均在1.8~2.1。按逆转录试剂盒(TaKaRa公司)说明书进行,将总mRNA逆转录为单链模板链cDNA,以cDNA为模板用PCR反应试剂盒进行PCR。引物序列:hsa-miR-2276-3p上游为5′-TGCGCTCTGCAAGT-GTCAGAGG-3′,下游为5′-GTCGTATCCAGTGCAGG-GTCCGAGGTATTCGCACTGGATACGACCCTCGCCT-3′;ITGA9 mRNA上游为5′-AACCCAGAAGAGGTG-ACGG-3′,下游为5′-GAGGATTCCCACCAACAAAC-3′;VEGF mRNA上游为5′-CAAAGCCAGCACATAGGAGAGAT-3′,下游为5′-TTTTTGCAGGAACATTTACACGTC-3′;GAPDH上游为5′-TCAAGAAGGTGGTGAAGCAGG-3′,下游为5′-TCAAAGGTGGAGGAGTGGGT-3′。PCR反应条件:95 ℃ 30 s,95 ℃ 5 s,60 ℃ 34 s,行 40个循环。以GAPDH作为内参,进行归一化。用2-ΔΔCt计算相对表达量。
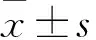
2 结果
2.1 绒毛组织中hsa-miR-2276-3p及ITGA9、VEGF mRNA表达情况比较 胚胎停育患者绒毛组织中hsa-miR-2276-3p相对表达量为2.05±0.52,高于正常早孕者的1.12±0.27(P<0.05);ITGA9、VEGF mRNA相对表达量分别为1.72±0.33、1.77±0.15,均低于正常早孕者的2.34±0.48、2.48±0.19(P均<0.05)。
2.2 胚胎停育患者绒毛组织中hsa-miR-2276-3p与ITGA9、VEGF mRNA表达的相关性 在胚胎停育患者绒毛组织中,has-miR-2276-3p与ITGA9、VEGF mRNA的表达均呈负相关(r分别为-0.679、-0.528,P均<0.05)。
3 讨论
目前,胚胎停育的发病机制不明,导致临床治疗困难。越来越多的证据表明,胎盘血管发育不良和内皮功能障碍可能导致胚胎停育的发生[6]。近年来研究发现,一些特异性表达在血管内皮细胞中的miRNA与胚胎的血管生成有密切关系,通过影响胎盘血管形成、细胞迁移和细胞侵袭、表观调控特别是DNA甲基化,影响早期胎盘发育和功能表达,在滋养细胞的增殖和凋亡中发挥重要作用[7],增加了胚胎停育的发生风险。
研究发现,hsa-miR-2276-3p特异性高表达于内皮细胞上,调控内皮细胞对VEGF的反应[8]。hsa-miR-2276-3p在国内外研究较少,仅有较少文献报道其在乳腺癌[9,10]、结肠癌[11]组织中表达下调,参与其肿瘤细胞的血管增生、侵袭与转移。因此,我们就has-miR-2276-3p是否也参与胚胎绒毛血管的生成进而调控胚胎的发育进行研究。前期研究发现,has-miR-2276-3p在胚胎绒毛组织中的血管内皮细胞中有特异性表达;本研究发现,hsa-miR-2276-3p在胚胎停育组绒毛组织中的表达量高于正常早孕者。这表明has-miR-2276-3p的上调可能与胚胎发育中血管生成减少有关,进而引起胚胎停育发生。我们推测,hsa-miR-2276-3p可能通过调控其他基因的表达影响绒毛滋养细胞成血管能力,包括增殖、迁移和成小管能力,作用于孕早期胚胎的发育。
整合素是细胞表面重要的黏附因子,可通过与配体的结合介导细胞之间以及细胞、胞外基质之间的信号转导,在细胞分化、胚胎发育及细胞黏附等方面发挥非常重要的作用[12]。整合素种类有很多,在机体内各种细胞内广泛表达,ITGA9是整合素α9β1重要组成成分。整合素α9β1的配体很多,可与神经生长因子、血管细胞黏附分子1、肌腱蛋白C、VEGF等相互作用。现已证实,整合素α9β1在细胞黏附和迁移、肺发育、淋巴管瓣和静脉瓣膜发育及伤口愈合过程中具有重要功能。整合素α9β1在不同的信号转导通路中调控细胞的增殖和分化,以及抗血管生成,起着不可或缺的作用[13]。目前,有关ITGA9的研究较少,其主要参与多种肿瘤细胞的生长、血管形成[14,15],可能是新的抗肿瘤、抗血管生成的靶点[16]。最近有研究发现,ITGA9突变可能严重影响胚胎发育[17,18]。本研究显示,ITGA9 mRNA在胚胎停育患者绒毛组织中的表达量低于正常早孕者。这提示ITGA9作为信号传导分子,其下调与胚胎停育发生有关。
VEGF是一类能够与肝素结合的糖蛋白,能够促使血管内皮细胞的分裂、增殖,促进新生血管的形成,并且能增强血管通透性[19]。在胚胎发育过程中,VEGF主要分布在胎盘的细胞滋养层细胞、合体滋养层细胞和绒毛间质细胞中。这提示在胎盘形成及植入过程中,VEGF具有促进胎盘绒毛血管网形成的作用。如果VEGF表达失衡,将会导致胎盘绒毛组织血管生成减少,胎盘血管生成障碍,胎儿供血不足,不能满足胎儿生长发育所需的足够氧气和营养物质,使胎儿的生长发育受限,最终引起胚胎停育的发生[20]。本研究显示,VEGF mRNA在胚胎停育患者绒毛组织中的表达量低于正常早孕者,提示我们VEGF表达减少与胚胎停育有关。
我们前期通过基因芯片测序发现,ITAG9是hsa-miR-2276-3p的一个靶基因;本次研究发现,胚胎停育患者绒毛组织中hsa-miR-2276-3p与ITGA9、VEGF mRNA呈负相关。因此,我们推测hsa-miR-2276-3p与ITGA9可能存在靶向关系。本研究提示,hsa-miR-2276-3p与ITGA9在胚胎停育的发生过程中有负向调节作用,ITGA9作为中间信号分子,可作用于与其配体VEGF。因此,我们推测hsa-miR-2276-3p在胚胎停育患者绒毛组织中的上调可能通过抑制ITGA9的表达进而下调VEGF的表达,影响血管新生,最终导致胚胎停育。因此,hsa-miR-2276-3p/ITGA9/VEGF可能是胚胎停育的一条新的信号通路。本研究具有很强的创新性及研究价值,可为胚胎停育发生提供新的理论基础。
