不同基因型高粱的磷效率和磷素转运特性研究
2018-03-19武爱莲王劲松南江宽董二伟焦晓燕平俊爱
刘 鹏,武爱莲,王劲松,南江宽,董二伟,焦晓燕,平俊爱
(1.山西大学生物工程学院,山西太原 030006;2.山西省农业科学院农业环境与资源研究所,山西太原 030031;3.山西省农业科学院高粱研究所,山西晋中 030600)
磷是植物生长所必需的大量元素之一,植物中的磷通过直接或间接参与糖类、脂肪以及蛋白质代谢等生理生化反应,从而影响着植物的生长发育[1]。土壤中磷元素很丰富,但由于磷易于被土壤吸附、固定的原因,导致出现土壤磷素的“遗传学缺乏”现象[2-3]。施用磷肥可有效缓解作物缺磷现象[4],但是由于磷肥难以被吸收,水稻、小麦与玉米等粮食作物投入磷肥的当年使用率只有10%~20%[5],导致磷素的浪费以及造成潜在的环境污染。我国79%的土地为中低产田[6],筛选具有高效吸收和利用磷素遗传潜力的作物品种,利用作物固有的生物学特性,挖掘作物自身高效吸收利用磷素的潜力,对实现作物高产、资源高效具有重要意义。
研究表明,磷效率是磷素吸收、同化和转运等多个生理过程综合作用的结果[4,7]。大麦[8]、马铃薯[9]、谷子[10]等作物在缺磷时可以感受外界胁迫,通过生理调节以及形态学改变以增强在低磷胁迫条件下的生存能力,其主要目的是在磷素有限的情况下吸收足够的磷(磷素吸收效率)以及高效利用已吸收的磷素从而形成更多的产量(磷素利用效率)[1]。此外,禾谷类作物开花后营养器官中的磷素逐渐向籽粒中转运,对籽粒磷素累积以及产量形成具有重要作用[11]。当作物处于磷胁迫时,根系所吸收的磷素远不能满足开花后籽粒的磷素需求,作物体内磷的再利用和转运就显得更为重要[12]。
高粱(Sorghum bicolor(L.)Moench)是世界第五大粮食作物[13],在饲料、能源、酿造以及食品等领域具有重要利用价值[14]。尽管高粱具有较强的耐瘠薄特性,适宜于种植在肥力较低的土壤上[15],但缺磷仍然是限制高粱产量的关键因素之一[13]。目前,有关高粱对低磷胁迫的反应以及耐低磷特性的研究滞后于其他粮食作物。
本研究通过盆栽试验,以4个耐瘠性不同的高粱基因型为材料,研究了不同基因型高粱磷素转运特性以及磷效率相关指标之间的关系,旨在为耐低磷型高粱品种的选育提供理论和实践依据。
1 材料和方法
1.1 试验材料
供试高粱品种为冀蚜2号,TX7000B,SX44B和TX378,由山西省农业科学院高粱研究所提供。其中,冀蚜2号和TX7000B为低磷敏感型材料,SX44B和TX378为耐低磷型材料。
1.2 试验设计
盆栽试验于2017年5—9月在山西省农业科学院连栋温室内进行。供试土壤质地为黏壤土,土壤养分含量为:有机质15.2g/kg,全氮含量0.65 g/kg,硝态氮5.7mg/kg,有效磷6.43mg/kg,速效钾105mg/kg,pH值 8.47,EC值 162 μS/cm。试验共设置低磷(LP)和正常供磷(HP)2个处理,每个处理各设6次重复。低磷处理P2O5用量为每千克风干土0.015 g,正常供磷处理为0.15 g;2个处理N和K2O用量分别为每千克风干土0.24,0.15 g。肥料和风干土以每盆为单元充分混匀后装盆,每个塑料盆共装9 kg。试验于5月6日播种,播前充分灌水,每盆种20粒种子以保证出苗效果。3叶期时每盆留苗5株,4叶期时每盆留苗3株直至收获。9月8日结束,出苗至收获一共122 d。
1.3 测定项目及方法
分别在开花期以及收获期以每盆为单元进行采样。花期植株样分为叶片和茎秆2个部位,成熟期植株样分为叶片、茎秆、穗芯以及籽粒4个部位。将植株样取下后按部位分开,去离子水冲洗干净后于105℃下杀青30 min,之后于65℃烘至恒质量,称质量后粉碎进行全磷测定。将植株样用体积比为1∶3的浓HClO4和浓HNO3消煮后,用钒钼黄比色法测定全磷含量[14]。
收获指数=籽粒产量/收获期地上部生物量;植株磷累积量=某一部位全磷浓度×该部位生物量;磷收获指数=籽粒磷累积量/植株磷累积量;磷效率=籽粒产量/施磷量;磷素吸收效率=植株磷累积量/施磷量;磷素利用效率=籽粒产量/植株磷累积量;磷转运率=(开花期茎叶等器官磷累积量-收获期对应各器官磷累积量)/开花期茎叶等器官磷累积量。
1.4 数据统计分析
采用Microsoft Office 2007和SPSS进行数据分析,表中数据均为3个重复的平均数±标准误。
2 结果与分析
2.1 不同基因型高粱在不同磷肥处理下的生物量和产量构成要素
与正常供磷相比,低磷处理下高粱花期整体延后,冀蚜2号,TX7000B,SX44B和TX378分别推迟2,6,1,2 d。从表1可以看出,施磷量对开花期高粱地上部生物量没有显著影响(P>0.05),对收获期冀蚜2号、TX7000B和TX378地上部生物量影响显著(P<0.05)。低磷处理下,冀蚜2号、TX7000B地上部生物量较正常供磷处理分别降低31.63%和15.07%,TX378降低13.66%,低磷敏感型高粱降幅大于耐低磷型高粱。
低磷处理显著降低了低磷敏感型高粱的籽粒产量(P<0.05),低磷处理下冀蚜2号和TX7000B的籽粒产量较正常供磷处理分别下降30.63%和26.88%,耐低磷型高粱SX44B籽粒产量虽然也有所下降,但是降幅仅为12.38%,说明缺磷对耐低磷型高粱的影响要小于低磷敏感型高粱。由表1可知,低磷处理下低磷敏感型高粱产量降低原因不同,冀蚜2号主要是由于籽粒数量减少,TX7000B则是籽粒数量减少和千粒质量变小共同导致的,缺磷抑制了TX7000B籽粒形成以及籽粒碳水化合物的填充。
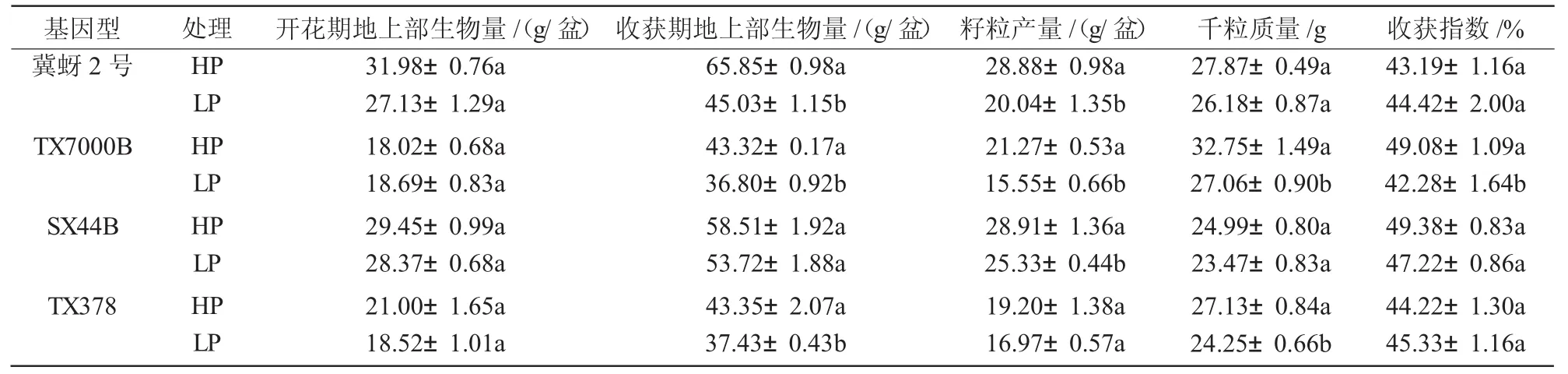
表1 不同磷处理对高粱生物量、产量构成要素的影响
2.2 不同基因型高粱的磷素累积量及磷效率
低磷处理显著降低了各基因型高粱的收获期植株磷累积量、籽粒磷累积量(P<0.05)(表2)。低磷敏感型高粱冀蚜2号、TX7000B低磷处理植株磷累积量仅为正常供磷处理的37.67%和52.20%,籽粒磷累积量分别为38.26%和47.01%,而耐低磷型高粱SX44B和TX378低磷处理下的植株磷累积量和籽粒磷累积量分别为正常供磷处理的67.99%,64.44%和83.84%,72.59%。同时,低磷胁迫也显著提高了耐低磷型高粱SX44B和TX378的磷收获指数,低磷敏感型高粱收获指数则没有影响或者显著降低(表 2)。
各基因型高粱磷效率2个磷处理间差异均显著,缺磷会显著提高高粱磷效率(P<0.05)(表2),正常供磷条件下,磷效率为32.57~49.05 g/g,低磷处理下高粱磷肥偏生产力提高至263.77~429.73 g/g。冀蚜2号和TX7000B低磷处理下的磷效率分别是正常供磷处理的 6.94,7.31倍,而SX44B和TX378分别是8.76,8.84倍。缺磷也会显著提高高粱的磷素吸收效率(P<0.05),低磷处理下的冀蚜2号和TX7000B的磷素吸收效率分别是正常供磷处理的3.77,5.22倍,SX44B和TX378分别是正常供磷处理的6.80,6.44倍。正常供磷和低磷处理下的相对磷效率和相对磷素吸收效率与高粱的耐低磷能力表现出较好的一致性。表2也反映出低磷处理显著提高了各基因型高粱的磷利用效率(P<0.05),耐低磷基因型SX44B和TX378的磷素利用效率的增加幅度低于低磷敏感基因型冀蚜2号和TX7000B。

表2 不同磷处理对高粱磷累积量、磷效率的影响
2.3 缺磷对不同基因型高粱植株磷浓度的影响
从表3可以看出,不同基因型高粱叶片和茎秆的磷分配有着明显差异(P<0.05)。在开花期,低磷敏感型高粱和耐低磷型高粱叶片和茎秆中磷的分配没有明显规律,冀蚜2号和SX44B表现出叶片磷浓度高于茎秆,而TX7000B和TX378茎秆磷浓度高于叶片,说明高粱叶片和茎秆磷分配与高粱是否耐低磷没有直接关系。在收获期,低磷处理显著降低了高粱叶片磷浓度,降低幅度和高粱是否耐低磷无关。低磷处理也显著降低了收获期茎秆中的磷浓度,相较于正常供磷处理,低磷处理下的冀蚜2号和TX7000B的茎秆磷浓度分别降低39.23%和25.63%,SX44B和 TX378分别降低 5.32%和51.05%,耐低磷型高粱茎秆磷浓度降低幅度大于低磷敏感型高粱。各基因型高粱均表现为籽粒磷浓度最高,耐低磷型高粱SX44B和TX378缺磷处理下的籽粒磷浓度较正常供磷处理分别降低4.30%和17.83%,低磷敏感型高粱冀蚜2号和TX7000B则分别降低39.37%和35.74%。

表3 不同磷处理对高粱各部位磷浓度的影响 g/kg
2.4 不同基因型高粱在不同磷肥处理下的磷素积累和转运
开花前期,作物以营养生长为中心,开花后转为以生殖生长为主,叶片和茎秆在开花前后会经历“源库”的转变[16-17]。低磷胁迫显著降低了各高粱基因型的花前植株磷素累积量(P<0.05)。开花后,高粱磷素“源库”关系发生转变,叶片和茎秆转变为磷素“汇”开始为籽粒提供大量的磷素。收获期各高粱基因型叶片和茎秆的磷素累积量较开花期都显著下降(P<0.05)(表4)。正常供磷处理下,叶片的磷素转运率为22.95%~76.08%,低磷处理下磷素转运率为54.20%~89.11%。茎秆在低磷处理下的磷素转运率为28.15%~68.52%,也高于正常供磷处理下的23.27%~55.12%,反映出缺磷会加剧叶片和茎秆磷素的转运。低磷处理下的低磷敏感型高粱冀蚜2号茎秆磷转运率较正常供磷处理增加20.93%,TX7000B茎秆磷转运率较正常供磷处理降低0.65%;耐低磷型高粱SX44B和TX378茎秆磷转运率分别增加92.12%和68.00%,均高于低磷敏感型高粱。
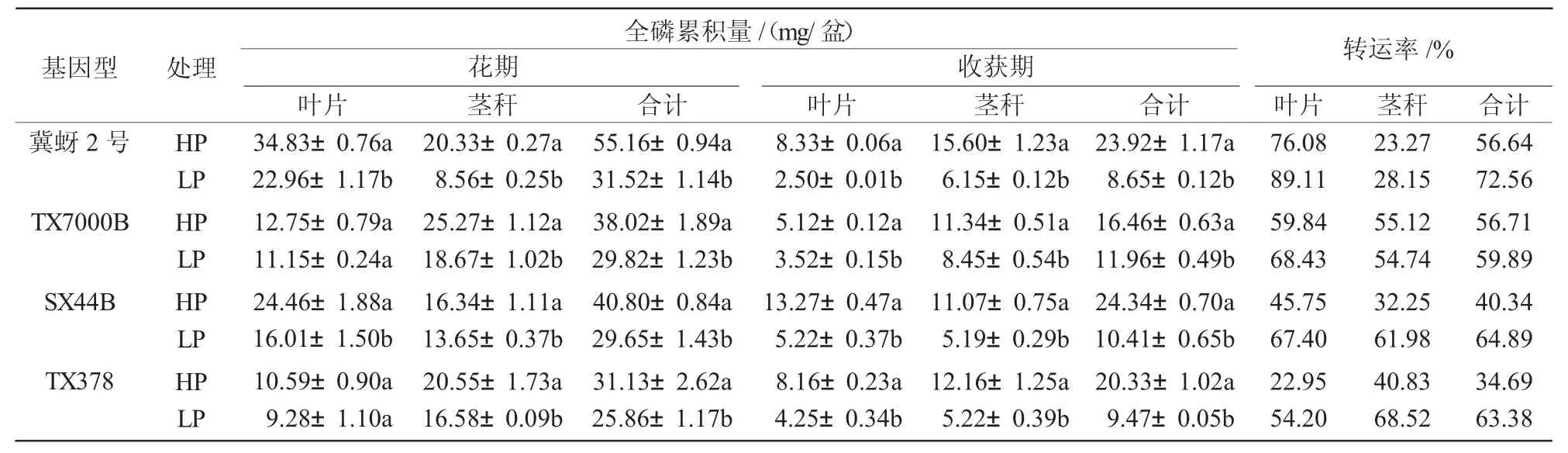
表4 不同磷处理对高粱各部位磷累积量和转运率的影响
3 讨论
磷是有机化合物(核酸与蛋白质、脂类、ATP等)的重要成分,磷本身也是代谢过程中的调节剂,缺磷将严重影响作物的生理以及产量性状[7,13-14]。不同作物或者是同一作物不同品种对磷素的吸收、利用以及转运存在明显的差异[9-10]。本研究表明,低磷胁迫会降低高粱的收获期地上部生物量和籽粒产量,但是耐低磷型高粱SX44B和TX378受缺磷的影响要小于低磷敏感型高粱冀蚜2号和TX7000B。
植物体内的磷素转运能力很强,易于被代谢活性高的器官再利用[12,18]。研究发现,玉米[3]、小麦[5]的耐低磷特性和磷素的再分配以及转运能力有关。本试验表明,低磷处理增加了高粱叶片磷素转运率,增加了耐低磷型高粱的茎秆磷转运率,对低磷敏感型高粱茎秆磷素转运影响很小。在开花后,营养器官中的磷素向生殖器官转运,对作物磷素累积、利用以及产量的形成具有重要意义[5,7,12],小麦、玉米等作物的籽粒中1/2以上的磷素均来自于营养器官的转运[19-20]。本试验中,在正常供磷处理下,高粱籽粒中的磷素有20.68%~46.02%来自营养器官的转运;低磷处理下,磷素转运增加,来自营养器官转运的磷素比例提高至43.21%~87.23%,低磷敏感型高粱籽粒中来自营养器官转运的磷素比例在2个处理下都高于耐低磷型高粱,这可能和耐低磷型高粱开花后磷素吸收能力更强,其营养器官磷素转运压力相对较小有关。低磷处理下,耐低磷型高粱的叶片磷素转运增加,但是茎秆的磷素转运率也同时增加。茎秆中磷素的再利用一定程度上缓解了叶片的磷素转运压力,这有助于维持叶片正常的生理功能,从而保证碳水化合物的合成以及光合产物的运输。
磷效率为籽粒产量与供磷量的比值[1,9-10],根据磷效率的定义,在同一供磷水平下,可以认为介质供磷量相同,即磷效率的大小可以用来表征籽粒产量[21]。从矿质营养学的角度看,作物磷高效是通过磷吸收、利用效率来实现的[4]。低磷胁迫下,植物会有2种应对策略:提高磷素吸收效率(PUpE)或者提高磷素利用效率(PUtE)[22-23]。对于磷效率而言,PUpE和PUtE哪个因素的作用更加重要尚无统一报道。WANG等[24]研究表明,作物的PUpE和PUtE对磷效率的相对贡献率和土壤的磷肥供应能力有关。对玉米[3,20]的研究表明,在高磷条件下,作物磷效率与磷素吸收效率以及利用效率都呈正相关。对谷子[10]和马铃薯[9]的研究表明,低磷条件下和高磷条件下磷素吸收和利用效率对磷效率的贡献不同。本试验中,在低磷胁迫条件下,耐低磷型高粱SX44B和TX378的相对磷吸收效率高于磷敏感型材料冀蚜2号和TX7000B,表明高粱的耐低磷特性和缺磷条件下高粱的磷素吸收能力密切相关。高粱在正常供磷条件下,籽粒产量和磷素吸收效率(r=0.943;P<0.01) 以及磷素利用效率(r=0.911;P<0.01)呈极显著正相关。而在低磷胁迫时,高粱的籽粒产量仍然和磷素吸收效率(r=0.814;P<0.01)呈极显著正相关,和磷素利用效率(r=-0.400;P>0.05)呈负相关,且并不显著。可见,当磷肥供应充足时,高粱的籽粒产量由磷素吸收和磷素利用效率共同决定,而在磷肥短缺条件下,高的磷素吸收效率对产量更加重要。
低磷胁迫下,作物会通过改变根系形态或者通过根系分泌有机酸、质子、酸性磷酸酶等分泌物,提高根系磷酸活性酶来适应低磷胁迫,导致根际环境酸化,提高环境磷的有效性以缓解磷胁迫压力[1,25-27]。本试验仅仅探讨了耐低磷型高粱的磷素分配以及转运特征,发现高粱耐低磷特性与磷素吸收能力密切相关,至于高粱在低磷胁迫条件下是通过何种方式提高磷素吸收能力仍需要进一步研究。
4 结论
低磷处理会降低高粱的收获期地上部生物量、籽粒产量和植株磷累积量,耐低磷型高粱在低磷胁迫下有着更高的相对地上部生物量、产量和植株磷累积量;低磷处理增加了高粱叶片磷素转运率,增加了耐低磷型高粱的茎秆磷转运率,对低磷敏感型高粱茎秆磷素转运影响很小;低磷处理下,高粱磷效率、磷素吸收效率均显著增加,耐低磷型高粱的增幅大于低磷敏感型高粱;正常供磷时,高粱磷型效率由磷吸收效率和磷利用效率共同决定,而在低磷胁迫下,高粱磷效率主要由磷吸收效率决定。
[1]CLEMENS C M,VAN DE WIEL,OLGA E,et al.Improving phosphorus use efficiencyin agriculture:opportunities for breeding[J].E-uphytica,2016,207(1):1-22.
[2]张锡洲,阳显斌,李廷轩,等.不同磷效率小麦对磷的吸收及根际土壤磷组分特征差异 [J].中国农业科学,2012,45(15):3083-3092.
[3]张丽梅,贺立源,李建生,等.不同耐低磷基因型玉米磷营养特性研究[J].中国农业科学,2005,38(1):110-115.
[4]唐珧,李丽君,白光洁,等.山西南部设施蔬菜施肥及土壤氮磷累积现状调查分析[J].山西农业科学,2017,45(5):773-776.
[5]阳显斌,张锡洲,李廷轩,等.小麦磷素利用效率的品种差异[J].应用生态学报,2012,23(1):60-66.
[6]YAN H,JI Y,LIU J,et al.Potential promoted productivity and spatial patterns of medium-and low-yield cropland land in China[J].Journal ofGeographical Sciences,2016,26(3):259-271.
[7]于天一,孙学武,王才斌,等.不同基因型花生磷素转运特性及磷效率研究[J].核农学报,2015,29(9):1813-1820.
[8]YANG T,HE J B,ZENG Y,et al.Differentiation of barley genotypes with high phosphorus efficiencyunder lowphosphorus stress[J].A-gricultural Science and Technology,2013,14(11):1615-1619.
[9]SANDAÑA P.Phosphorus uptake and utilization efficiency in response to potato genotype and phosphorus availability[J].European Journal ofAgronomy,2016,76:95-106.
[10]GEMENETDC,HASH CT,SANOGOMD,et al.Phosphorus uptake and utilization efficiency in West African pearl millet inbred lines[J].Field Crops Research,2015,171:54-66.
[11]GEMENET D C,LEISER W L,BEGGI F,et al.Overcoming phos-phorus deficiency in west African pearl millet and sorghum production systems:promising options for crop improvement[J].Frontiers in Plant Science,2016,7:1389.
[12]樊明寿,陈刚,孙国荣.低磷胁迫下玉米根中磷的运转与再利用[J].作物学报,2006,32(6):946-948.
[13] LEISER W L,RATTUNDE H F,PIEPHO H P,et al.Getting the most out ofsorghumlow-input field trials in west Africa usingspatial adjustment[J].Journal of Agronomy&Crop Science,2012,198(5):349-359.
[14]王劲松,焦晓燕,丁玉川,等.粒用高粱养分吸收、产量及品质对氮磷钾营养的响应[J].作物学报,2015,41(8):1269-1278.
[15]FARRÉ I,FACI J M.Comparative response of maize (Zea mays L.)and sorghum(Sorghum bicolor L.Moench)todeficit irrigation in a Mediterranean environment[J].Agricultural Water Management,2006,83(1):135-143.
[16]MASONI A,ERCOLI L,MARIOTTI M,et al.Post-anthesis accumulation and remobilization of dry matter,nitrogen and phosphorus in durumwheat as affected bysoil type[J].European Journal ofA-gronomy,2007,26(3):179-186.
[17] VAN OOSTEROM E J,CHAPMAN S C,BORRELL A K,et al.Functional dynamics of the nitrogen balance of sorghum.II.Grain fillingperiod[J].Field Crops Research,2009,115(1):29-38.
[18]张丽梅,郭再华,张琳,等.缺磷对不同耐低磷玉米基因型酸性磷酸酶活性的影响 [J].植物营养与肥料学报,2015,21(4):898-910.
[19]党红凯,李瑞奇,孙亚辉,等.超高产栽培条件下冬小麦对锰的吸收、积累和分配 [J].植物营养与肥料学报,2010,16(3):575-583.
[20]王艳,孙杰,王荣萍,等.玉米自交系吸收利用磷素的差异及其相关性[J].应用与环境生物学报,2003,9(5):479-481.
[21]戢林,李廷轩,张锡洲,等.氮高效利用基因型水稻根系形态和活力特征[J].中国农业科学,2012,45(23):4770-4781.
[22] VANCE C P,UHDESTONE C,ALLAN D L.Phosphorus acquistion and use:critical adaptations by plants for securing a nonrenewable resource[J].NewPhytologist,2003,157(3):423-447.
[23]RICHARDSON A E,LYNCH J P,RYAN P R,et al.Plant and microbial strategies to improve the phosphorus efficiency of agriculture[J].Plant&Soil,2011,349(1/2):121-156.
[24]WANG X R,SHEN J B,LIAO H.Acquisition or utilization,which is more critical for enhancing phosphorus efficiency in modern crops?[J].Plant Science,2010,179(4):302-306.
[25]丁广大,陈水森,石磊,等.植物耐低磷胁迫的遗传调控机理研究进展[J].植物营养与肥料学报,2013,19(3):733-744.
[26]邱双,刘利军,闫双堆,等.低磷胁迫下谷子的磷吸收利用效率[J].山西农业科学,2016,44(6):786-789.
[27]魏猛,张爱君,李洪民,等.长期施肥条件下黄潮土有效磷对磷盈亏的响应[J].华北农学报,2015,30(6):226-232.
