董箐库区水环境变化对鱼类分布影响
2018-03-16王从锋秦孝辉
王 瑶 王从锋,2 秦孝辉
(1. 三峡大学 水利与环境学院,湖北 宜昌 443002; 2. 三峡地区地质灾害与生态环境湖北省协同创新中心, 湖北 宜昌 443002; 3. 珠江水利科学研究院,广州 510611)
库区的建立可以通过增加渔业水域面积的方式实现养殖增产,但另一方面又因阻断特殊鱼种洄游制约了鱼类资源多样性的发展,对天然鱼类资源、物种多样性、种质资源产生了严重影响[1].张慧杰[2]等指出水流的流态、速度和水质是影响葛洲坝坝下鱼类分布的重要因素;任玉芹[3]等在研究三峡库区支流鱼类分布的差异性时,认为鱼类的生活习性、地形、水深等对鱼类具有一定影响.在本文所研究的北盘江流域,有学者对光照建坝前后库区内鱼类资源通过传统渔获物调查的方式进行比较研究,其结论是光照电站的建设对库区河段的鱼类资源影响显著[4].随着计算机技术的发展及仪器性能的提高,相比于传统的水声学调查方法,具有速度快、准确性高等优点,能够实现整个水体的观测,提供丰富的鱼类时空分布数据[5].目前,许多国内外学者应用美国BioSonics DT-X科学探鱼仪、图像声呐DIDSON等水声学方法对鱼类资源量进行评估[6-8],但在本文所涉及的北盘江流域,该研究首次采用挪威Simrad EY-60科学探鱼仪,分别在2015年4月、7月、9月、12月对董箐库区鱼类资源进行水声学走航式调查,同步进行水质监测,旨在结合时空尺度上的水质、流速变化特性,研究鱼类资源的时空分布特征以及环境因子改变对鱼类资源分布所造成的差异性影响,为库区鱼类资源的保护提供一定的参考.
1 材料和方法
1.1 研究区域
董箐水库作为北盘江下游最末级电站,是连接北盘江、南盘江和红水河流域鱼类洄游和种群交流的重要纽带,因此其库区鱼类具有较高的研究价值.
水库调节库容1.44亿m3,坝址处多年平均流量398 m3/s,多年年平均径流量为125.55亿m3,多年平均径流深623.3 mm(如图1所示).董箐库区河道地形、水文特性较为特殊,分为干流和支流两大水域,支流属典型的高山峡谷型(“V”型)河道,河道狭窄,流速较大,水力特性较为复杂.而其支流打帮河大部分河道与“U”型相似,江面相对宽阔,流速相对较小,水力特性较为简单.故文章重点研究干流水域的水文、水质、鱼类资源情况.

图1 董箐库区监测采样走航示意图
1.2 研究方法
检测路线采用“之”字形走航式方法,航速保持在6~8 km/h范围之内.探测过程中将EY60固定于船右舷,淹没深度0.5 m.该研究所采用的EY60回声探测仪换能器工作频率为120 kHz,其波束形式为分裂波束,波束扩散角为7°,采样间隔时间为64 μs.在航线的每个拐点处同步采集水质参数,包括水温、pH、DO、流速及水深,监测仪器采用哈希公司生产的HydrolabDS5多参数水质分析仪测定,每1 m水深读取一次数据,数据导出到Excel中处理.流速运用挪威Nortek公司生产的VectrinoPlus声学多普勒点式流速仪测定.由于研究区域水深过大(大部分区域水深在50 m左右),数据线较短,采用hydrolabDS5离线测量模式,以人工拉索绳的方式将多参水质分析仪探头按0.1 m/s的速率放入水中,索绳长30 m.计算出每个区域的水质参数及流速平均值,利用Excel进行初步统计.
1.3 数据处理计算
在水声学方法中,目标强度是根据回波积分值对鱼类资源的个体组成及资源密度评估的重要参数,该研究采用Foote[9]提出的有鳔鱼类鱼体体长与目标强度的经验公式TS=20logL-71.9对流域内鱼类资源生长结构进行估算,其中TS为鱼体目标强度;L为鱼体体长.当鱼类个体分散密度较低时,Misund[10]指出通过水声学仪器直接计数的方法进行密度估算.水质数据由于得到的各监测点水深数据随机性较大,根据监测数据计算出每个区域的水质参数及流速平均值.将四次监测的水质参数进行统计,分析年度的水质参数的差异性,P<0.05时差异具有统计学意义.为了分析引起鱼类密度变化的决定性因素,本文划分了水质参数及流速的区间范围,将不同环境参数值(水质、流速、时间)分别作为固定因素,将鱼类密度作为因变量,运用SPSS19软件,通过单因素方差(ANOVA)方法来分析.
2 结果分析
2.1 不同季节鱼类目标强度分布
鱼类目标强度百分比分布正态性检验结果表明,4个季节的目标强度分布呈偏态分布,春、夏、秋、冬四季的目标强度平均值分别为-47.84±3.27 dB、-52.94±2.85 dB、-53.47±2.07 dB、-51.36±2.17 dB,对4个季节采用单因素方差分析结果显示,不同季节的TS百分比分布呈显著性差异(F=25.64,P=0.024).多重方差分析显示,春季与夏季、秋季差异较显著,且春季目标强度显著大于夏季、秋季;夏、秋、冬3个季节无显著性差异(P值均大于0.05).
如图2所示,春季鱼类目标强度百分比值随体长的增加,先增加后减小的趋势,其中目标强度为-55~-40 dB的鱼体所占百分比为82.5%,对应的体长范围为7~39.36 cm;夏季鱼类目标强度百分比值随体长呈锯齿状变化,且-52~-49 dB体长的百分比显著小于附近体长范围的百分比值.与春季相比,目标强度大于-40 dB以上鱼类体长所占比例有所增加;秋季鱼类目标强度为-58~-52 dB所占百分比为49.3%,占到所有体长的一半,其对应的体长为4.95~9.89 cm,秋季的鱼类体长百分比随体长的增加,基本呈减小的趋势,且鱼类体长在-46~-30 dB之间相对春、夏季向高体长值迁移;冬季鱼类体长分布相对“矮胖”.随季节的变化,董箐库区春、冬两个季节体长分布较为符合鱼类自然增长规律.春季到夏季之间鱼类以成熟个体开始产卵、孵化,到进入秋季时大多数受精卵长成幼小育苗,且每年的3月、9月初在董箐库区进行保护鱼种的鱼苗放流,数量均为万尾以上,秋季鱼类主要以幼小鱼体为主.

图2 不同季节鱼类目标强度百分比分布
2.2 鱼类资源密度分布
在2015年间共4次的走航式水声学探测,由于走航探测的连续性,在分析计算鱼类密度时,将走航式探测的数据以约0.5 nm(约900 m)的基本采样距离单元进行分割,而在分析区域时将整个库区分成9个子区域(Ⅰ~Ⅸ),故所有区域平均鱼类密度均以平均值±标准差表示(见表1).

表1 2015年董箐库区鱼类平均密度分布(单位:ind/1 000 m3)
干流、近坝区以及支流的水下地形、流速流态和水质特征的差异决定了鱼类密度在时空分布上的差异性.表1给出了整个董箐库区鱼类密度检测结果.从数据看出干流区域(Ⅰ~Ⅳ分区)的相比于其他区域具有更为丰富的鱼类密度.对年度内不同季节的鱼类资源进行K-S检验,检验发现4月份鱼类密度非正态分布,故对鱼类密度数据进行对数转换,正态检验所得sig.均大于0.05,接受原假设.由ANOVA方差分析结果得出:显著性sig.为0.033,因此不同季节的鱼类密度季节差异性具有统计学意义.
2.3 同步水环境因子监测
2.3.1 流速监测结果分析
通过分析监测点流速值沿断面变化情况发现,从测点D01到D11,流速的年度变化维持在0~1.2 m/s范围之内(如图3所示).

图3 监测点流速变化
因为董箐库区属日调节电站,受上游马马崖电站日调节来水影响,除汛期以外,整个库区的流速分布无明显的季节特征.对4、7、9、12月份的流速进行正态性检验,结果显示流速年度变化上的差异性并不显著(Kruskal-Wallis检验结果的sig.值大于0.05),且两两之间并无显著差异,这与库区内流速的年际变化实际情况相一致.空间分布上,水体流速受地形及河道影响,具有较为明显的差异性.
2.3.2 温度、DO、pH、浊度分析
为分析各参数指标在不同季节的差异性,对不同季节的温度、DO、pH、浊度进方差分析及Kruskal-Wallis检验.结果显示除溶氧服从正态分布(sig.值为0.144、0.200*、0.152、0.200*),另外3个参数只有部分季节服从正态分布.对上述3个因子(温度除外,温度受季节影响差异性显著)分别进行相应的统计:从不同季度间溶氧值的LSD多重分析结果发现四个季节溶氧值存在显著性变化(P<0.05),其中4月与12月、7月与9月之间差异最为显著.对pH、浊度分别进行Kruskal-Wallis检验,检验结果sig.值均为小于0.05,故拒绝原假设,认为pH和浊度在季节差异性上变化显著.根据spss分析结果初步看出,在流域范围上季节变化除了影响温度和降雨条件,同时也影响了水体的水质状况,包括pH、浊度和pH值等,所以这些水质参数的改变对鱼类资源的影响程度需要进一步的分析验证.
2.3.3 水质因子与鱼类资源的相关性分析
初步分析发现影响鱼类资源时空分布的水质因子具有复杂性,因此对其进行因素整合分析,并结合鱼类密度与多项水质参数进行相关性分析.该研究鉴于各因子在数据上的叠加性,运用Bartlett球型和KMO检验法检验各因子的相关关系,其中KMO的值为0.416,显著性水平为0.000,说明这些指标之间至少有两个是相关的,进行主成分分析最终计算得主成分函数如下:
Y1=0.58×水温+0.34×溶氧+0.62×浊度+0.40×pH
Y2=-0.38×水温+0.64×溶氧
Y3=-0.07×水温-0.05×溶氧-0.31×浊度+0.61×pH
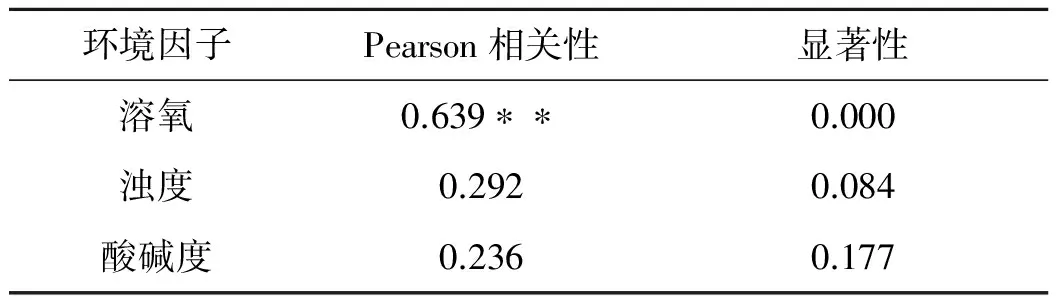
表2 主成分因子与鱼类密度的相关性
**.置信度(双侧)为0.01时,显著性是相关的
从主成分表达式可以看出,在第一主成分Y1上浊度具有较高的载荷,第二主成分Y2上溶氧载荷较高第三主成分,Y3则主要依赖于pH值.综合来看,溶氧、浊度以及pH值是影响鱼类资源分布的主要原因.表2给出了经过Pearson相关性分析的结果,发现董箐库区鱼类资源密度与水体中溶氧值显著相关,与浊度和pH呈微弱相关.
3 讨 论
3.1 影响董箐库区鱼类资源分布水质因素
影响鱼类资源在时间尺度上分布的差异性主要体现在水温、浊度、pH、溶氧等因子随季节性变化呈现出的变化,由于监测条件限制,该研究只对本次监测所得的温度、溶氧、pH、浊度以及流速进行分析研究.通过主成分分析及Pearson相关性分析提取了影响鱼类分布的3个主成分,分别为溶氧、浊度和pH.许多研究指出温度是影响鱼类分布的重要参量,但在该研究中,通过提取主成分,发现温度与溶氧、浊度及pH具有一定的相关性,因此不考虑温度变量.通过分析将溶氧确定为影响库区鱼类资源分布的主要因子,在Y2主成分上具有0.64的高载荷系数,说明溶氧作为影响鱼类资源分布的主要的水质因子是被接受的.
在董箐库区所进行的4次监测中,4月、7月以及9月份的水体溶氧值要明显高于冬季12月份,从生物代谢角度而言,溶氧通过鱼鳃经血液循环运送到鱼的各个组织,保证鱼类的正常生理活动[11].Wilhelm Filho[12]指出高溶解氧水平可提高鱼类的生长速度、调节鱼类的生理平衡,而减少溶解氧含量能够限制鱼的摄食活动,或者改变动物体对食物的消化和吸收.因此,鱼类生活的水体,溶解氧必须满足呼吸的需要[13].杨凯等通过测定不同溶氧水平下的黄颡鱼稚鱼摄食、生长及呼吸代谢过程,发现溶解氧的差异能够直接影响鱼体的摄食活动和营养物质的利用效率[14].在流域内,不同的鱼类对水中溶氧的需求量不同,鱼类在水中缺氧时,就向溶氧丰富水域游去.故笔者认为,造成鱼类密度季节差异性的原因为春夏秋三季水体中较高的溶氧值为董箐库区的鱼类资源提供了良好的生长繁殖条件,旺盛的生命活动使得鱼类密度显著上升,所以导致溶氧成为制约董箐库区鱼类资源时间序列上的分布差异性的关键因素.
浊度和pH分别作为反映水中悬浮物质多少和水体酸碱程度的量化指标,对鱼体的生境选择也有着相应关系.由于夏季汛期由于上游来水量增大,干支流来水夹杂的泥沙和微小颗粒直接导致董箐库区的浊度升高,其中对鱼类生存最直接的影响是对水体光线的阻碍作用,浊度较大的水体有助于幼鱼躲避敌害,因此幼鱼多偏向于栖息在浊度较大的水体.刘淑德等人[15]的研究还指出浊度对于鱼类还存在着间接影响,从其生活环境方面来讲,浊度越大致使水体的透明度越差,不利于浮游植物的生长繁殖,从而影响鱼类资源的分布.除浊度影响外,鱼类生活的水体pH需保持在一定的范围之内,超出特定范围会引起鱼类死亡.弱酸性水体中,鱼体的载氧能力要明显下降,导致生存能力下降.整个董箐库区的水体pH介于7.7~8.4之间,常年呈弱碱性,较适合鱼类的水体栖息选择,故与鱼类资源密度呈弱相关性特征.
3.2 流速对鱼类资源的影响
该研究按走航路线所获取的鱼类资源密度数据显示,董箐库区干流的鱼类资源密度值最大,发现干流Ⅰ~Ⅳ分区的流速与近坝区和支流的流速具有显著性的差异,决定了干流区域的鱼类密度大于近坝区及支流,得出董箐库区鱼类资源对流速具有较为明显的选择性的结论,且鱼类资源大多集中在流态稳定,水面较为开阔的流速较大水体(如干流Ⅰ、Ⅱ分区).究其原因可能位于河口断面变化明显的地方,该位置相对于其余各个分区,水流流态复杂多变,其丰富的水下地形和流态为不同生态习性(喜急流、缓流的鱼类为主)的鱼类提供了栖息地、索饵场甚至产卵场,说明了水下地形对鱼类栖息有着重要影响,水体环境中鱼类密度的分布与水文条件密切相关[16].生活在水中的鱼类对于流速的变化具有敏感的反应机能,趋流性的鱼类主要依靠流速的大小判断其行进路线[17],同时流速的大小也影响鱼类在行进过程中的耗能方式.许多研究表明流速对鱼类的生境选择有着重要的影响:不同流速下的鱼类聚集和栖息行为具有显著的差异性,熊锋[18]等通过对葛洲坝1号船闸运行时的流速变化与鱼类行为进行监测,亦发现鱼类密度与水流流速的相关关系.从整个董箐库区而言,因其丰富的水地形,导致不同区域的水动力特性差异显著,相关资料显示该库区存在喜急流、喜流水以及静缓均有的生态群落结构,因此流速的空间差异性对于维持生物尤其是鱼类资源的多样性有着积极的作用.
由于条件限制,本研究只对董箐库区鱼类资源与部分水环境因子进行了相关性的研究,未深入的讨论通过改善水环境对鱼类资源进行保护和恢复.在后期研究中,应进一步结合水库生态调度理论,科学的提出鱼类资源保护和水生态修复的具体工程措施.
[1] 汤娇雯,张 富,陈兆波.我国鱼类生物多样性保护策略[J].淡水渔业,2009,39(4):75-79.
[2] 张慧杰,杨德国,危起伟,等.葛洲坝至古老背江段鱼类的水声学调查[J].长江流域资源与环境,2007,16(1):86-91.
[3] 任玉芹,陈大庆,刘绍平,等.三峡库区澎溪河鱼类时空分布特征的水声学研究[J].生态学报,2012,32(6):1734-1744.
[4] 周 路,张竹青,林艳红,等.梯级开发对北盘江中下游鱼类资源的影响[J].贵州农业科学,2011,39(10):134-137.
[5] 刘慧杰,王从锋,刘德富,等.双频识别声呐在鱼类资源调查中的应用进展[J].三峡大学学报(自然科学版),2015,37(3):28-34.
[6] 张 赞,蔺丹清,汤 勇,等.基于声学测量方法的大伙房水库鱼类资源季节变动特征[J].南方水产科学,2014,10(6):12-19.
[7] 孙明波,谷孝鸿,曾庆飞,等.基于水声学方法的太湖鱼类空间分布和资源量评估[J]. 湖泊科学, 2013,25(1): 99-107.
[8] 周家飞,王从锋,刘德富,等.葛洲坝下游近坝区水域鱼类资源声学调查与评估[J]. 长江流域资源与环境, 2014,23(11): 1551-1557.
[9] Foote K G. Fish Target Strengths for Use in Echo Integrator Surveys[J]. Journal of the Acoustical Society of America, 1987, 82(3): 981-987.
[10] Misund O A. Underwater Acoustics in Marine Fisheries and Fisheries Research[J]. Reviews in Fish Biology and Fisheries, 1997, 7(1): 1-34.
[11] 王得祥. 水流紊动对鱼类影响实验研究[D].南京:河海大学, 2007.
[12] Filho D W, Torres M A, Zaniboni-Filho E, et al. Effect of Different Oxygen Tensions on Weight Gain, Feed Conversion, and Antioxidant Status in Piapara, Leporinus Elongatus (Valenciennes, 1847)[J]. Aquaculture, 2005, 244(1): 349-357.
[13] 王 珂.三峡库区鱼类时空分布特征及与相关因子关系分析[D].北京:中国水利水电科学研究院, 2013.
[14] 杨 凯,樊启学,张 磊,等.溶氧水平对黄颡鱼稚鱼摄食、生长及呼吸代谢的影响[J]. 淡水渔业, 2010,40(2): 24-29.
[15] 刘淑德.长江口及其邻近海域鱼类浮游生物群落结构特征研究[D].青岛:中国科学院研究生院(海洋研究所), 2009.
[16] Imre I, Dlg M L R.Phenotypic Plasticity in Brook Charr: Changes in Caudal fin Induced by Water Flow[J]. Journal of Fish Biology, 2002, 61(5): 1171-1181.
[17] 袁 喜,李丽萍,涂志英,等.鱼类生理和生态行为对河流生态因子响应研究进展[J]. 长江流域资源与环境, 2012(S1): 24-29.
[18] 熊 锋,王从锋,刘德富,等.葛洲坝1号船闸启闭闸门对近闸区域鱼类活动规律的影响[J]. 水生态学杂志, 2014,35(5): 8-14.
