外源硅对盐胁迫下黄芩幼苗生理生化特性的影响
2018-03-16华智锐李小玲
华智锐,李小玲
(商洛学院 生物医药与食品工程学院,陕西 商洛 726000)
黄芩(ScutellariabaicalensisGeorgi)属于唇形科黄芩属多年生草本植物,以根入药,具有抗菌、消炎、调节血脂及抗艾滋病病毒等功效,为大宗常用中药材之一[1]。近几年来,黄芩在中药生产、黄酮化合物提取等方面具有较高的应用价值,市场需求量巨大[2]。但由于对野生黄芩资源的过度采挖,目前黄芩药材生产主要来源仍然依靠人工的大田栽培和引种驯化,而日益加重的土壤盐渍化现象给黄芩大田生产构成了严重威胁。
硅是地球表面的第二大元素,对大多数高等植物的生长有益[3],有促进植物(尤其是单子叶植物)的生长和增加生物学产量的功能。硅对植物生长发育的影响有直接和间接两种途径,直接途径是指硅参与植物生理生化代谢过程,间接途径则通过改善土壤氮、钾等元素的利用效率而促进植物生长[4]。添加硅可以提高盐胁迫下玉米幼苗地上部和根系干重,增加其根冠比[3]。研究表明,硅(K2SiO3)能促进植株的生长,还能改善植株对矿物质的营养吸收[5]。另外,硅在提高植物对生物及非生物胁迫抗性方面也具有重要作用[5]。Liang(1998)[6]研究发现,加入适宜浓度的外源硅可减轻盐胁迫下大麦叶绿体的结构损伤,有利于保护叶绿体膜的完整性。在盐胁迫条件下适宜浓度的外源硅会进一步促进小麦幼苗的生长[3]。外源硅不仅能提高干旱胁迫下番茄根系线粒体活性氧清除酶的活性,减轻活性氧的积累和膜脂过氧化程度,还有利于维持根系细胞及重要细胞器结构的完整性,进而增强植株的抗旱性[6]。硅处理后大麦幼苗根系的CAT、SOD、POD、GSH活性提高,丙二醛含量降低,从而使大麦盐胁迫的过氧化伤害程度降低。硅能增强盐胁迫下玉米硝酸还原酶的活性,促进蛋白质的合成,提高叶片的光合速率[3],增强叶绿体清除活性氧的能力,从而缓解盐胁迫对叶绿体膜的伤害[4]。
1 材料与方法
1.1 试验材料
选取一年生商洛黄芩幼苗进行盆栽试验,待植株在盆中培养时间达到3个月时,进行处理并测定相关指标。
1.2 试验方法
1.2.1 材料预培养 预处理:将所购买的黄芩幼苗移栽至直径40 cm、高55 cm左右的花盆中进行预培养,在此期间定时进行浇水和松土以保持土壤湿润。3个月后选取生长一致的黄芩幼苗用于试验。
1.2.2 试验设计 本试验共设6组,两个对照组和4个试验组:CK1(蒸馏水对照);CK2(0.5%盐对照);Si0,0.01 g/L K2SiO3+0.5% NaCl;Si1,0.025 g/L K2SiO3+0.5% NaCl;Si2,0.05 g/L K2SiO3+0.5% NaCl;Si3,0.1 g/L K2SiO3+0.5% NaCl。对上述6组试验用不同浓度外源硅采用灌根方式处理8 d(每2 d处理一次),然后选取长势良好的植株进行试验。摘取适宜的叶片,洗净,用蒸馏水冲洗,取出主叶脉后,剪碎,用于测定幼苗保护酶(SOD、POD、CAT)活性、丙二醛(MDA)和脯氨酸的含量等生理指标。
1.2.3 生理指标的测定
1.2.3.1 过氧化物酶(POD)活性的测定 酶液的提取:称取0.5 g黄芩叶片于研钵中,加入2 mL磷酸缓冲液,研成匀浆,将其转入离心管中,于8000 r/min下离心10 min;取上清液转入25 mL 容量瓶,沉淀用5 mL磷酸缓冲液再提一次,上清液转入25 mL容量瓶定容。
POD的测定:加入20 μL酶液、3 mL反应液于比色皿中,在470 nm下每隔1 min读1次数,共读3次,以每分钟吸光度变化值计算POD活性:POD活性[(△A470/(min·g )]=△A470×V/Va/W=△A470×5/0.02/0.5=△A470×500 。式中△A470为反应时间内OD变化值;V为提取酶液总体积(mL);W为黄芩叶片鲜重(g);Va为测定时取用酶液体积(mL)。
1.2.3.2 过氧化氢酶(CAT)活性的测定 酶液提取:称取黄芩叶片0.5 g置于研钵中,加入2~3 mL 4 ℃下预冷的磷酸缓冲液和少许石英砂,研磨成匀浆,然后转入25 mL容量瓶中,用缓冲液冲洗研钵后合并冲洗液,并定容至刻度,混合均匀,将容量瓶置5 ℃冰箱中静止10 min,取上清液于4000 r/min下离心15 min,5 ℃下保存备用。
CAT的测定:0.1 mL酶液+2.5 mL反应液,在240 nm下比色,每隔1 min读数1次,共读3次。
班主任工作是平凡而繁琐的工作,我们只有从点滴做起,从小事做起,因势利导,做到润物细无声。浇灌出一朵朵希望之花,让这些美丽的花朵,盛开在祖国肥沃的土地上。
结果计算:CAT活性[△A240/(min·g)]=△A240×V/Va/W=△A240×5/0.05/0.5=△A240×200。△A240为反应时间内OD变化值。
1.2.3.3 超氧化物歧化酶(SOD)活性的测定 酶液提取:称取黄芩叶片0.5 g置于预冷的研钵中,加1 mL预冷的磷酸缓冲液研磨成浆,加缓冲液使终体积为5 mL,取2 mL于1000 r/min下离心20 min,上清液即为SOD粗提液。
SOD的测定:取型号相同的试管,吸取20 mL酶液,加入3 mL反应液,4000 lx照光30 min,同时取4支试管,3支作对照,1支做空白(不加酶液,以缓冲液代替);空白管放置暗处,对照(CK)与酶液同置于4000 lx条件下光照30 min,避光保存,以空白调零,于560 nm条件下比色。
将NBT光还原50%定义为一个SOD活性单位。SOD总活性(吸光度/g)=(ACK-AE)×V/(W×0.5×ACK) 。
ACK为照光对照管反应液的吸光度;AE为样品管反应混合液的吸光度;V为提取酶液总体积(mL);W为黄芩叶片鲜重(g)。
SOD比活性(酶单位/mg)=SOD总活性/蛋白质浓度。
1.2.3.4 丙二醛(MDA)含量的测定 MDA提取:称取黄芩叶片0.5 g置于研钵中,加入2 mL 10%三氯乙酸(TCA)和少量的石英砂,研磨至匀浆,再加入8 mL 10% TCA进一步研磨,将其转入离心管,于4000 r/min下离心10 min,除去沉淀,上清液即为样品提取液。
显色反应及测定:取提取液2 mL(对照加2 mL蒸馏水),加入2 mL 0.6% TBA(硫代巴比妥酸)液,转入试管中(试管加塞)沸水浴15 min,迅速冷却后,在4000 r/min下离心2 min。取上清液分别测定532 nm、600 nm和450 nm条件下的吸光值。
结果计算:MDA(μmol/g)=[6.45×(D532-D600)-0.56×D450]×0.015/W或[6.45×(D532-D600)-0.56×D450]×0.03/W。式中D450、D532、D600分别为在450、532、600 nm波长下测得的吸光度值;W为黄芩叶片鲜重(g)。
1.2.3.5 脯氨酸含量的测定 提取脯氨酸:分别称取胚轴,每种处理各取3份,每份0.3 g,剪碎,加入适量80%乙醇、少量石英砂,于研钵中研磨成匀浆,将匀浆液全部转移至25 mL刻度试管中后用80%乙醇冲洗研钵,将洗液移入相应的试管中,最后用80%乙醇定容至刻度,混匀,80 ℃水浴中提取20 min。
脯氨酸的测定:称取黄芩叶片0.3 g,加入5 mL磺基水杨酸,加盖,沸水浴10 min,过滤,吸取滤液2 mL(同时作空白,吸取2 mL蒸馏水)、2 mL冰醋酸和3 mL酸性茚三酮,沸水浴40 min,冷却,再加入5 mL甲苯,充分振荡,静止分层,取上层甲苯溶液于比色皿中,于520 nm下比色。
结果计算:脯氨酸含量(μg/g)=C×V/Va/W。其中,C为脯氨酸浓度;V为提取酶液总体积(mL);W为黄芩叶片鲜重(g);Va为测定时取用酶液体积(mL)。
1.2.4 数据处理方法 所有处理每次测定均重复3次,数据取3次测定的平均值,采用Excel 2007进行数据统计,用SPASS 17.0软件进行数据处理和统计分析。
2 结果与分析
2.1 外源硅对盐胁迫下黄芩幼苗中过氧化物酶(POD)活性的影响
由图1可知,在0.5% NaCl处理下POD活性比CK1明显下降;在0.5% NaCl胁迫下随着外源硅浓度的升高POD的活性呈先升后降的趋势,其中Si0、Si1、Si2、Si3的活性与对照(CK2)相比分别提高了33.33%、100%、120%、86.67%。Si2处理的POD活性达到最佳,是CK2的2.2倍,差异达极显著水平(P<0.01)。而Si1、Si2、Si3的活性与对照(CK1)相比分别提高了20%、32%、12%;在Si2下POD活性达到最佳,是CK1的1.32倍,差异达显著水平(P<0.05)。由此可知,与对照相比,Si2处理(0.05 g/L K2SiO3)能有效提高POD的活性,缓解盐胁迫对黄芩幼苗的伤害。
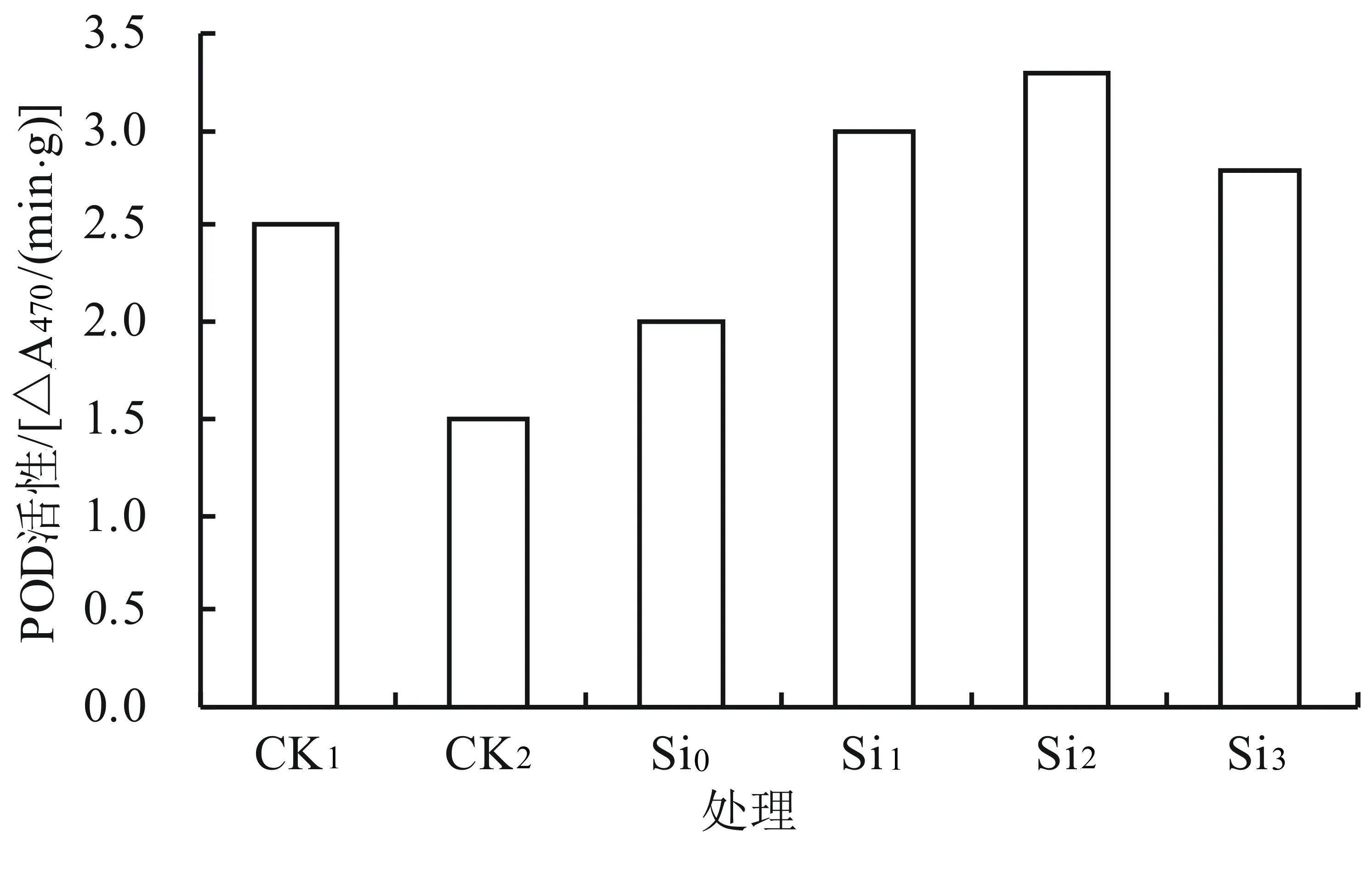
图1 外源硅对盐胁迫下黄芩幼苗中POD活性的影响
2.2 外源硅对盐胁迫下黄芩幼苗中过氧化氢酶(CAT)活性的影响
由图2可知,在0.5% NaCl处理下CAT的活性比CK1明显下降;在0.5% NaCl胁迫下随着外源硅浓度的升高CAT的活性呈先升后降的趋势,其中Si0、Si1、Si2、Si3的活性与对照(CK2)相比分别提高了11.1%、52.3%、60.4%、11.5%;在Si2下CAT活性达到最佳,分别是CK1、CK2的1.08、1.60倍,差异均达显著水平(P<0.05);而Si1、Si2处理的CAT活性与对照(CK1)相比分别提高了2.7%、8.3%。由此可知,与对照相比,Si2处理(0.05 g/L K2SiO3)能有效提高CAT的活性,缓解盐胁迫对黄芩幼苗的伤害。
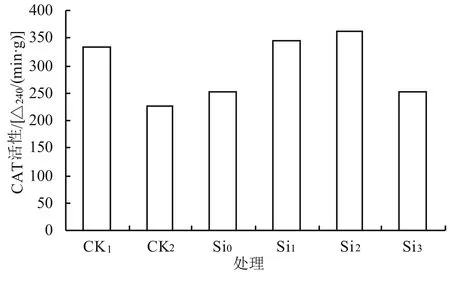
图2 外源硅对盐胁迫下黄芩幼苗中CAT活性的影响
2.3 外源硅对盐胁迫下黄芩幼苗中过氧化物歧化酶(SOD)活性的影响
由图3可知,0.5% NaCl处理的SOD活性比CK1有所下降;在 0.5% NaCl胁迫下随着外源硅浓度升高SOD的活性呈先升后降的趋势,其中Si0、Si1、Si2、Si3的活性与对照(CK2)相比分别提高了25.2%、39.4%、6.7%、1.8%;Si1处理的SOD活性达到最佳,分别是CK1、CK2的1.19、1.39倍,差异均达显著水平(P<0.05),而Si0、Si1的SOD活性与对照(CK1)相比分别提高了7.0%、19.2%。由此可知,与对照相比,Si1处理(0.025 g/L K2SiO3)能有效提高SOD的活性,缓解盐胁迫对黄芩幼苗的伤害。
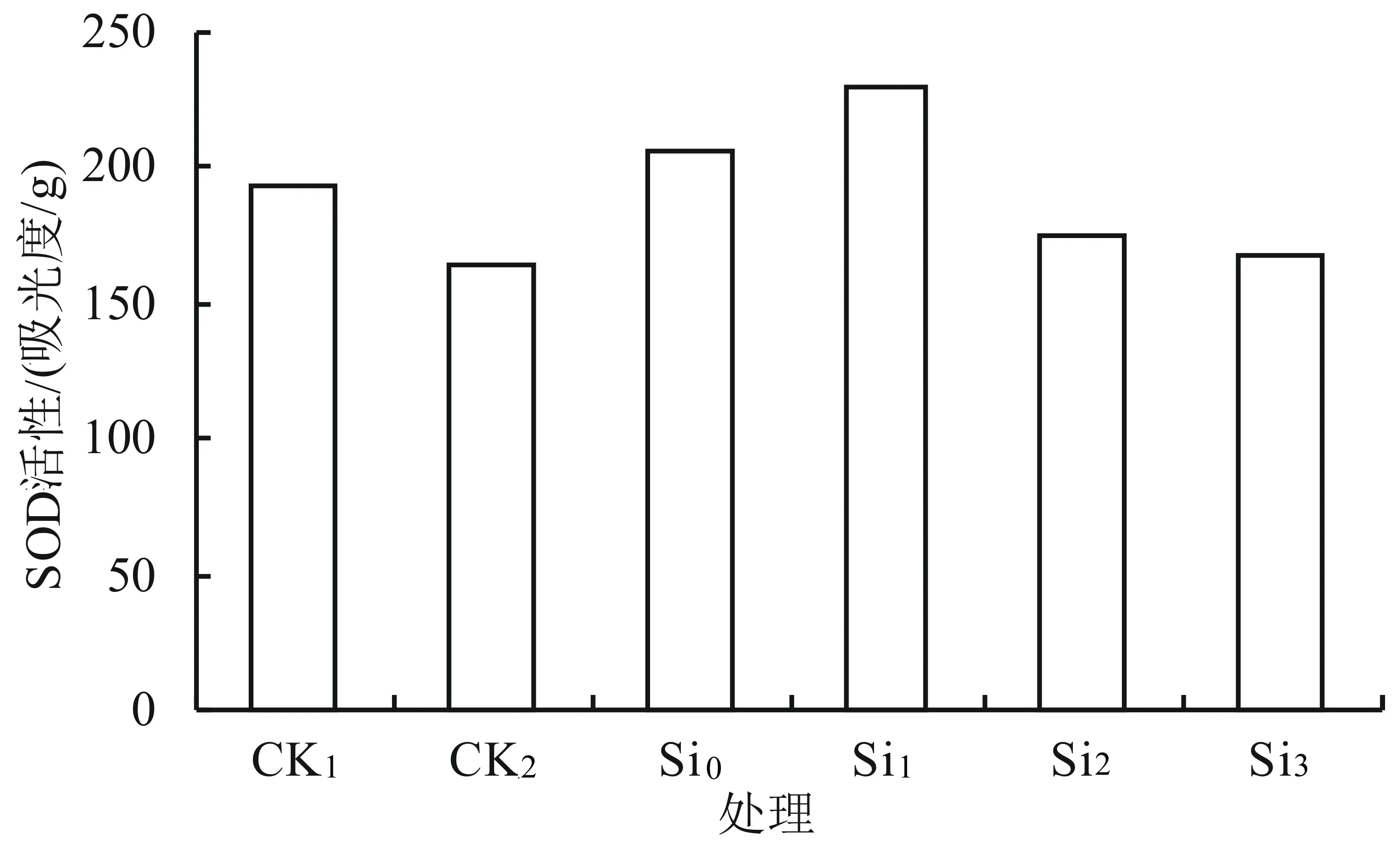
图3 外源硅对盐胁迫下黄芩幼苗中SOD活性的影响
2.4 外源硅对盐胁迫下黄芩幼苗中丙二醛(MDA)含量的影响
由图4可知,0.5% NaCl处理的MDA含量比CK1明显上升,增加了71.3%;不同浓度的外源硅在0.5% NaCl胁迫下对商洛黄芩幼苗叶片中MDA含量的影响存在不同差异,随着外源硅浓度的增加MDA含量基本上呈先增后降的趋势,在Si2时达到最高,与CK1相比增加了48.3%。各浓度硅处理中MDA含量均显著低于CK2(P<0.05),Si0、Si3处理中MDA含量均显著低于CK1,而Si1、Si2处理中的与CK1相比分别增加了9.2%、48.3%。由此可见,外源硅可缓解盐胁迫对黄芩的伤害,降低细胞膜伤害程度。
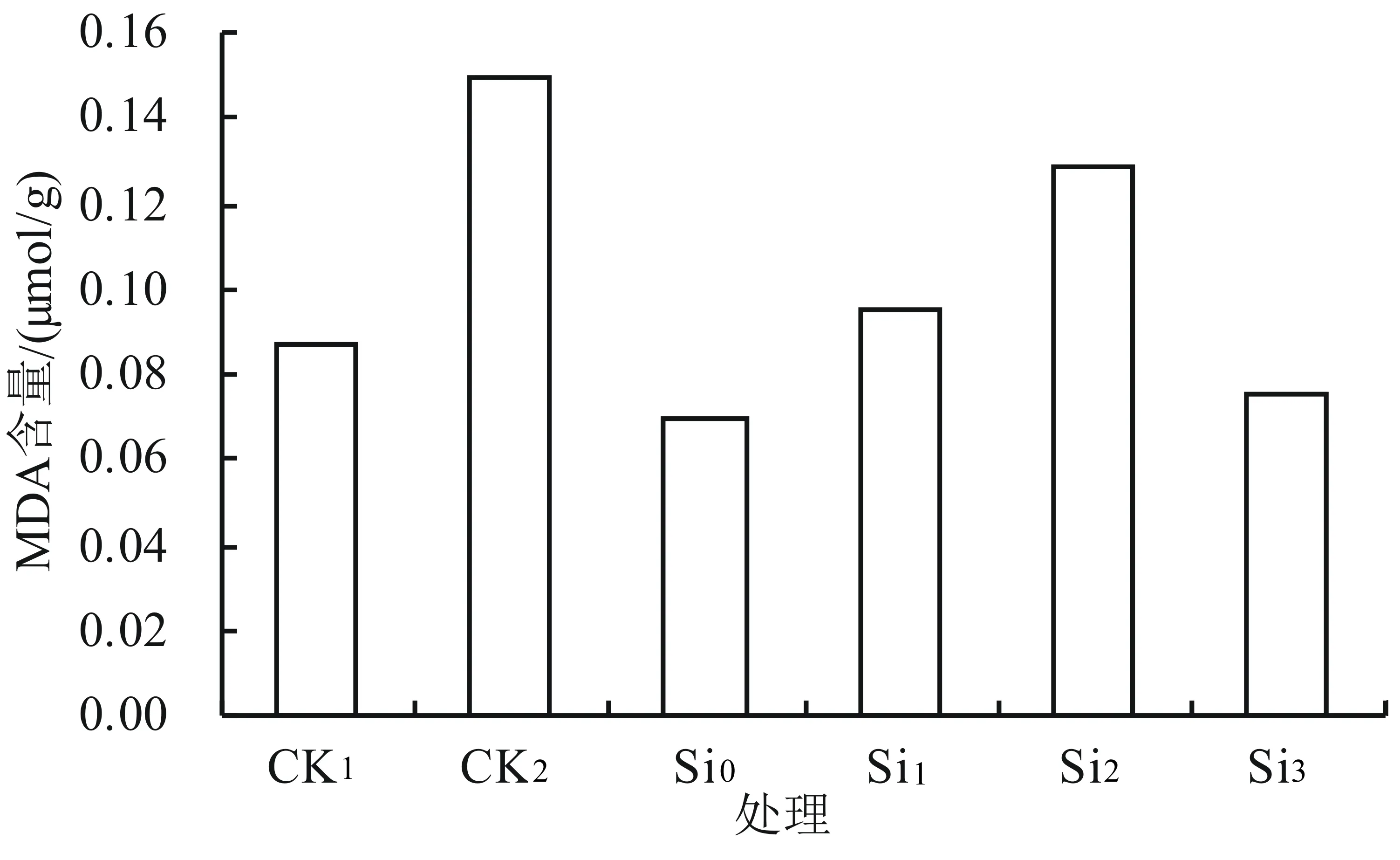
图4 外源硅对盐胁迫下黄芩幼苗中丙二醛含量的影响
2.5 外源硅对盐胁迫下黄芩幼苗中脯氨酸含量的影响
由图5可知,盐胁迫处理(CK2)能够增加黄芩叶片中的脯氨酸含量,与对照(CK1)相比增加了71.9%;不同浓度的外源硅在0.5% NaCl胁迫下对黄芩幼苗叶片中脯氨酸含量的影响存在不同差异,随着外源硅浓度的升高,黄芩叶片中的脯氨酸含量呈先增后降的趋势;在Si1时达到最高,是CK1的1.5倍;各浓度硅处理中脯氨酸含量均显著低于CK2(P<0.05);与对照CK1相比,Si0、Si1、Si2、Si3处理中脯氨酸含量分别增加了40.6%、50.0%、28.1%、6.3%。由此可见,外源硅可降低盐胁迫下脯氨酸的积累量。
3 讨论
SOD是植物处于逆境条件下细胞内清除活性氧系统中的重要抗氧化酶,对维持植物体内的活性氧代谢的平衡和保护膜结构起着重要作用,能提高植物组织的抗氧化能力[8];其与超氧化物阴离子自由基(O2-)发生歧化反应,生成O2和H2O2,生成的H2O2可以被过氧化氢酶(CAT)分解成O2和H2O,可以避免H2O2积累对细胞的氧化破坏作用。在本研究的盐胁迫条件下,不同浓度外源硅明显提高了黄芩幼苗中SOD、POD、CAT的活性,这与沙滩黄芩[9]、黄瓜[10]、甘草[11]的研究结果相一致。本试验中,在0.5% NaCl胁迫下,用不同浓度外源硅处理商洛盆栽黄芩幼苗,发现黄芩幼苗中SOD、POD、CAT活性缓解趋势相似,其中SOD、POD、CAT活性均高于盐对照组(CK2)。由此说明不同浓度外源硅具有提高盐胁迫下黄芩幼苗中SOD、POD、CAT活性的作用,其中POD、CAT活性在Si2处理时表现最佳,SOD活性在Si1时效果最明显。
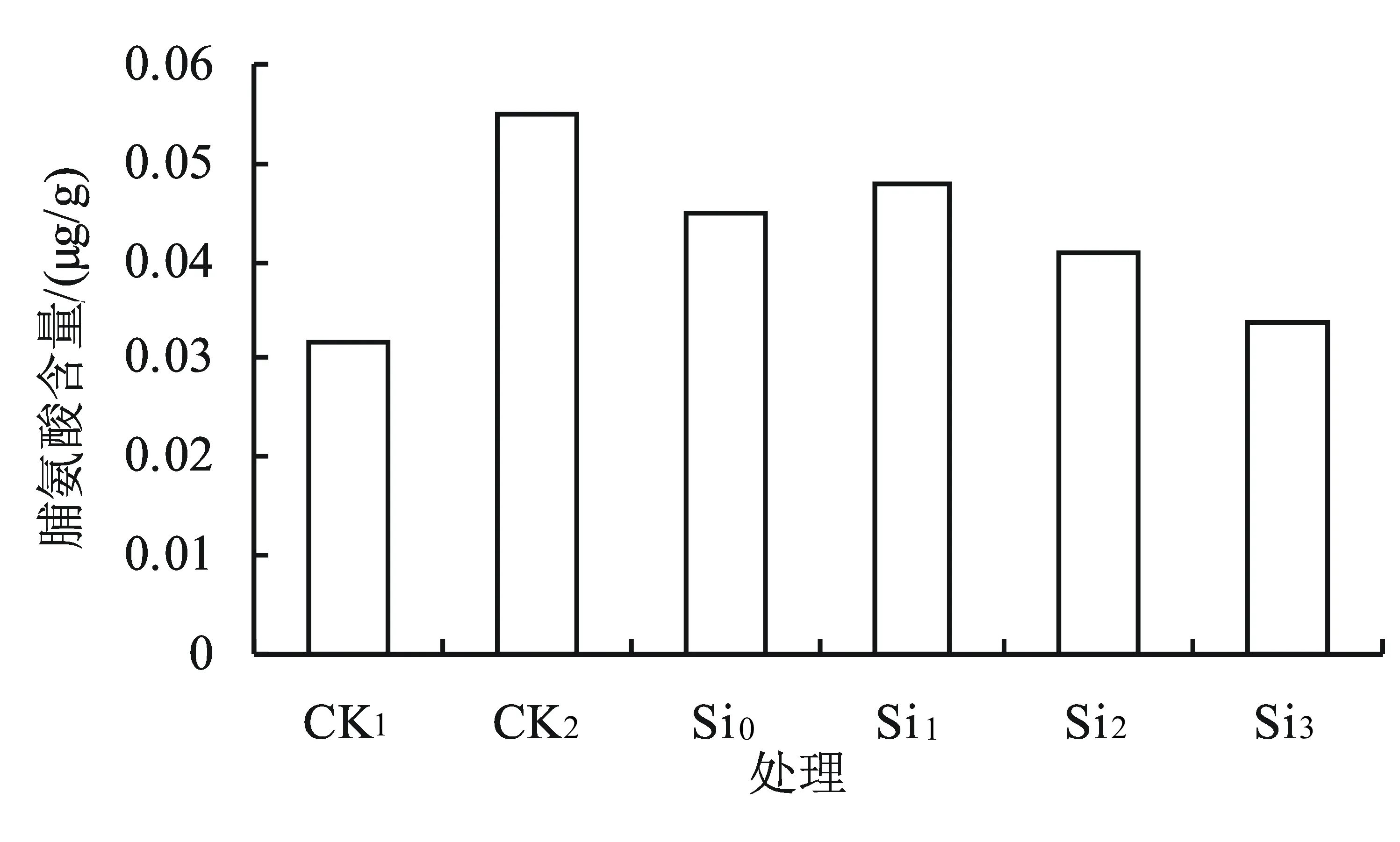
图5外源硅对盐胁迫下黄芩幼苗中脯氨酸含量的影响
盐胁迫会严重危害植物的细胞膜结构和组成。由于盐胁迫中离子的胁迫作用改变植物的细胞膜功能,质膜受到伤害,细胞内电解质外渗,随着逆境加剧膜破坏的程度增加[7]。在盐胁迫下,植物最先接触到盐分并且受到伤害的是细胞质膜,并导致膜的透性增大和膜脂过氧化加剧,而膜脂过氧化产物对植物的防御体系破坏又再次加重了膜脂的过氧化作用程度。已有研究表明,加外源硅处理可显著降低植物叶片电解质外渗率和过氧化物产物丙二醛(MDA)的含量[12-13]。
丙二醛是膜脂过氧化作用的产物之一,它能够与膜蛋白发生交联作用,使膜的透性增大;又可以与细胞内的各种成分发生反应,使膜系统中的多种酶生理功能严重受到损害。因此,可以用MDA的含量来代表植物膜脂过氧化作用的强弱,反映植物受伤害的程度和植物对逆境的反应[9,14],所以在盐胁迫下MDA含量的高低可反映黄芩幼苗叶片中生物膜受伤害的程度及其耐盐性的强弱。在0.5% NaCl胁迫下,MDA的含量增加,经过不同浓度的外源硅处理后的黄芩幼苗中MDA含量均低于对照组(CK2),说明不同浓度的外源硅可以缓解膜脂过氧化的过程,降低MDA的含量。研究表明,随着外源硅浓度的增加MDA的含量基本上呈先增后降的趋势,各浓度硅处理中MDA含量均显著低于CK2(P<0.05),Si0、Si3中MDA含量均显著低于CK1(P<0.05),由此说明适宜浓度的外源硅可缓解盐胁迫对黄芩幼苗的伤害,且当外源硅达到一定浓度时还可降低细胞膜伤害程度。
脯氨酸是一种重要的渗透调节物质, 在盐胁迫下植物体内大量积累脯氨酸, 可以避免细胞内外渗透势失衡而造成植物细胞的生理性缺水,有助于增加细胞的持水力,从而保护原生质和质膜的完整性[10]。在逆境胁迫条件下,与不耐盐植物相比,耐盐植物体内积累了更多的脯氨酸,能提高植物的耐盐性。本试验中,黄芩幼苗受到盐胁迫后,脯氨酸含量显著增大,各浓度硅处理中脯氨酸含量均显著低于CK2(P<0.05),与对照CK1相比Si0、Si1、Si2、Si3处理中脯氨酸含量分别增加了40.6%、50.0%、28.1%、6.3%,且小于CK2,由此可见,不同浓度的外源硅可降低盐胁迫下脯氨酸的积累量,这可能是硅缓解黄芩幼苗盐分伤害的一种生理响应机制。
本试验结果表明,用不同浓度外源硅处理盐胁迫下商洛盆栽黄芩幼苗,可使SOD、POD、CAT活性增加,提高植物细胞内抗氧化酶活性,增强植物的抗逆性;同时可降低盐胁迫下丙二醛(MDA)和脯氨酸的积累量,缓解盐胁迫对细胞膜的伤害程度。本试验结果为黄芩的耐盐性研究提供了参考,也为利用外源硅作为化控措施以缓解黄芩盐害提供了理论依据。但本研究主要是通过盆栽试验完成了相关数据采集,在黄芩的大田栽培试验真实环境中相关生理指标和响应机制的研究还有待进一步进行。
[1] 孙方行,孙明高,夏阳,等.NaCl胁迫对柴荆幼苗保护酶系统的影响[J].中南林学院学报,2005,25(6):34-37.
[2] 王玉萍,王映霞,白向利,等.硅对NaCl胁迫下甜瓜种子萌发及幼苗生长的影响[J].甘肃农业大学学报,2015,24(5):108-116.
[3] 孟长军,邹志荣.外源ALA对樱桃番茄幼苗盐伤害的缓解效应[J].江苏农业学报,2011,27(2):378-381.
[4] 齐曼.尤努斯,李秀霞,等.盐胁迫对大果沙枣膜脂过氧化和保护酶活性的影响[J].干旱区研究,2005,22(4):503-507.
[5] 阎秀峰,李晶,祖元刚.干旱胁迫对红松幼苗保护酶活性及脂质过氧化作用的影响[J].生态学报,1999,19(6):850-854.
[6] 石庆华,林嘉鹏,姚正培,等.NaCl胁迫对小麦生理生化特征的影响[J].新疆农业科学,2010,47(7):1479-1484.
[7] 郭云平,王秀峰,李岩.NaCl胁迫对野生和栽培西瓜幼苗生理特性的影响[J].山东农业科学,2016,48(2):45-48.
[8] 江绪文,李贺勤,王建华,等.盐胁迫下黄芩种子萌发及幼苗对外源抗坏血酸的生理响应[J].植物生理学报,2015,51(2):166-170.
[9] 王胜,丁雪梅,时彦平,等.盐胁迫对沙滩黄芩生长及其生理特性的影响[J].山东林业科技,2015,25(5):35-37.
[10] 张平艳,高荣广,杨凤娟,等.硅对连作黄瓜幼苗光合特性和抗氧化酶活性的影响[J].应用生态学报,2014,25(6):1733-1738.
[11] 崔佳佳,张新慧,李月彤,等.外源Si对盐胁迫下甘草幼苗形态及生理指标的影响[J].草业学报,2015,24(10):214-220.
[12] 陈罡,樊平声,冯伟民,等.外源硅对盐胁迫下黄瓜幼苗生长和光合荧光特性的影响[J].江苏农业学报,2014,30(6):1402-1409.
[13] 沈慧,米永伟,王龙强.外源硅对盐胁迫下黑果枸杞幼苗生理特性的影响[J].草地学报,2015,20(3):43-45.
[14] 宫德衬.小麦抗旱性生理指标研究[D].烟台:烟台大学,2014:1-62.
