北京城市公园环境下自生植物物种组成及多样性时空特征
2018-03-08李晓鹏关军洪吴思佳
李晓鹏,董 丽,关军洪,赵 凡,吴思佳
北京林业大学,园林学院, 国家花卉工程技术研究中心,城乡生态环境北京实验室, 北京 100083
长期以来城市绿地中的植物景观是被大面积的人工草坪和栽培群落所主导,这种整齐华丽的景观不仅需要消耗大量的人力、物力及不可再生资源,更造成了本土植物群落种间关系失衡、生物多样性急剧下降、各城市景观同质化严重等一系列问题,且生态服务价值不高[1-3]。与此同时,那些能在城市中自发生长繁衍的植被则被冠以“杂草”之名,几乎在园林绿地中没有容身之处。随着生态环境的持续恶化,人类生态意识逐渐提高,开始越来越多地关注起“杂草”这一植物群体。20世纪70年代,spontaneous vegetation最早被国外生态学者所使用,泛指自然定居生长的植物群体[4- 6]。本世纪初,这类城市中无需过多养护管理、可自播繁衍、野趣美感十足的自生植物逐渐引起了欧美景观设计师的关注,既而在风景园林领域开展了相关的理论和实践研究,认为这类植物在构建可持续、低维护园林植物景观中具有重要地位[7-9],并且相比城市栽培群落可更好地发挥生态效益,如作为动物栖息地、吸附棕地土壤重金属等[10- 12]。同时,国外研究者对其在城市环境中的分布特征、自生植物群落组合设计与筛选[13- 14]等方面也进行了初步的研究,自生植物逐渐成为园林植物景观规划设计中一类不容忽视、十分重要的组成部分。正是基于此,本研究亦采用自生植物这一名称来指代这类未经人工栽培而在城市环境中自发定居生长的植物群体。
在国内,虽有学者已指出“杂草”这一名称的产生带有强烈的主观色彩,并非科学上的分类[15],但目前研究者所使用的名称仍以“杂草”或野生植物为多。在生态领域对其进行的研究主要以城市建成区为基底,研究侧重该类植物在城市化影响下的物种组成、多样性和群落分类[16- 22]。而在园林行业,对这类植物进行的研究多以城市绿地中的杂草防治为目的,仅有少量学者针对其景观表现及园林应用潜力进行了初步探讨[23- 24]。此外,由于自生植物生长周期较短,以往研究由于目的不同,未重视其丰富的时间变化特点和时空分布格局这些可反映多样性维持机制且具有景观意义的特征。城市绿地功能多样,生境特殊且外来影响因子复杂多变,这些都必将会影响自生植物的分布,但目前自生植物在城市绿地中的物种组成、种间关系、多样性及维持机制尚不明确,相关研究少有报道。公园绿地作为城市绿地系统中规模较大、功能最为综合的绿地类型,也最迫切需要营造可持续、低维护的植物景观,同时其多样的绿地环境和空间类型也必然为城市自生植物繁衍提供最佳场所,而明确自生植物在其间的时空分布特征、群落动态和景观特征是对其进行合理规划和应用的重要前提。北京奥林匹克森林公园作为北京城区最大的城市公园,建成8年以来已成为北京市建成区最为重要的“绿肺”之一,并为许多动植物提供了丰富的栖息地。由此,本研究以北京奥林匹克森林公园为例,首先针对自生植物在整个生长季时间和空间上的物种组成及多样性特征进行分析,其次以物种优势度为依据进行不同季节和生境下自生植物群落聚类,最后对影响其时空特征的因素、启示及景观效果进行了讨论,以期为低维护自生植物群落的营建与应用提供指导。
1 研究地区与研究方法
1.1 研究地区概况
北京奥林匹克森林公园位于北京中轴线的北端,地理坐标为40°00′N,116°22′E,占地680hm2,被北五环划分为南园和北园两部分。其所在地区北京市为典型的暖温带半湿润大陆性季风气候,西北部被太行山、燕山环抱,中南部是由潮白河和永定河冲积而成的大平原,总面积1.68万km2,平原海拔高度20—60m,山地海拔1000—1500m,优越的地理环境孕育了丰富的动植物资源,但同时城市建成区的高速发展也在挑战着生态环境的极限。
1.2 样点及样方设置
通过网格系统取样法,利用Google Earth和GPS仪沿着正南正北方向设置200m×200m的网格对公园区域进行布点,去掉落在水面的网格点,部分样点依据实际情况进行微调,最终选择了123个调研样点(图1)。以每个调研样点为中心设置1个20m×20m的样方,并记录其中的乔木和灌木种以及株数、株高、冠幅和胸径。采取平均布样法在每个样方的四角和中心设置5个1m×1m的小样方进行自生植物的调查,记录其中出现的物种名、株数和盖度,以此作为每个样方的综合数据。全园小样方总计615个,所有小样方于2015年的3月末、4月末、6月初、7月末、8月末、10月初、11月中共调研7次。按照每个样方在公园中所处的位置和所包含的景观元素划分为路边、林地、公园边缘、园路与水体之间、水边、园路与广场之间、园路与建筑之间、园路与公园边缘之间和铺装广场9个生境类型。
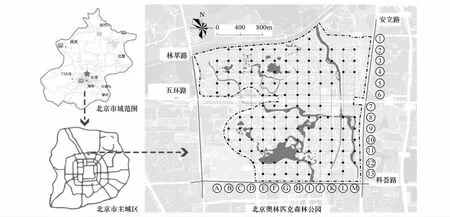
图1 研究地及样点设置Fig.1 Study area and sampling design A—M为样点的横向编号,1—13为样点的纵向编号
1.3 数据计算及统计分析
1)频度(Frequency)[25]:
F= (Si/N)×100%
式中,Si为物种i出现的样方数,N为总样方数,即123。
2)简化优势度(Simplified dominance):
Sdi=(Ci×100+Ai)/2
式中,Sdi为物种i的简化优势度,Ci为样方内物种i的盖度即物种i地上部分垂直投影面积占地面积的比率;Ai为物种i在样方中的个体数。
3)Shannon-Wiener多样性指数[25]:
H=-∑PilnPi
式中,Pi为物种i的简化优势度比例。
样方整理及数据计算运用R语言的vegan包和spaa包[26]进行,物种数、Shannon-Wiener多样性指数的均值比较采取Duncan检验法(α=0.05)运用R语言的agricolae包进行;群落聚类利用SPSS 19.0采用组间pearson相关系数法;统计图由Excel和R语言绘制。对植物物种信息的整理参考《北京植物志》和《中国植物志》。其中,乡土植物(native species)为自然分布于北京地区的物种,外来植物(alien species)指由人类有意或无意引进的北京范围以外的物种,分为国外外来和国内外来植物,通过查询植物志和历史文献确定[27-30]。入侵植物为在北京地区存在大量个体及有大范围扩散潜力的有入侵认定的物种,参考相关文献确定[29-30]。
2 结果与分析
2.1 奥林匹克森林公园自生植物整体概况
全园共调查到自生植物128种,隶属于32科、98属(附表1)。其中,一、二年生植物74种(57.82%)、多年生植物51种(39.84%)、木本植物3种(2.34%)。在物种来源方面,乡土植物98种(76.56%),如蒲公英(Taraxacummongolicum)、二月蓝(Orychophragmusviolaceus)、车前(Plantagoasiatica)等,国内外来植物6种(4.69%),如灰绿藜(Chenopodiumglaucum)、秃疮花(Dicranostigmaleptopodum)等,国外外来植物24种(18.75%),如小蓬草(Erigeroncanadensis)、钻形紫菀(Astersubulatus)等;入侵植物包括牛膝菊(Galinsogaparviflora)、小蓬草、意大利苍耳(Xanthiumitalicum)、钻形紫菀等16种(12.50%)。各科所含物种数差异明显,菊科(Asteraceae)植物物种数最多,共计26种;其次是禾本科(Gramineae)12种,蝶形花科(Papilionaceae)11种。全园物种频度排名前十的依次是:狗尾草(Setariaviridis)(96.75%)、蒲公英(86.18%)、抱茎苦荬菜(Ixerissonchifolium)(86.18%)、灰菜(Chenopodiumalbum)(79.67%)、早开堇菜(Violaprionantha)(74.80%)、车前(73.17%)、牛筋草(Eleusineindica)(70.73%)、狗牙根(Cynodondactylon)(69.11%)、二月蓝(55.28%)、夏至草(Lagopsissupina)(54.47%)。
2.2 自生植物物种组成的时空特征
从3月末到11月中,记录到的物种数呈单峰型变化。3月末物种数仅为48种,8月末物种数量达到最高峰,为102种。11月自生植物逐渐进入休眠期,物种数降到49种。各生境下多年生草本物种数全年较为稳定,一年生草本物种数变化则呈单峰型,并于8月末超过了多年生草本,达到全年最高(44种)后开始逐渐减少(图2)。全园各生境类型样方数量排序为:路边(43)、林地(40)、公园边缘(16)、园路与水体之间(9)、水边(6)、园路与广场之间(4)、园路与建筑之间(3)、园路与公园边缘之间(1)和铺装广场(1),整个生长季各生境所含物种数依次是109、106、91、90、87、57、60、33、14。物种数随着生境数量的减少表现出递减的趋势,路边的物种数在所有生境中最多,8月末记录到81种,其中外来植物22种。样方数大于3的生境类型外来植物所占比例见图3。整体上外来植物物种数与物种总数的时空变化基本保持一致,而各生境中外来植物物种数所占比例也符合这一特点,其值在8.33%—27.16%之间变动,并且于8月末和10月初达到最高,此时各生境外来物种数比例平均值分别为23.56%和23.97%。
科组成在时间上的变化特征为:菊科和禾本科植物物种数在整个生长季显著高于其他科,分别在6月初和7月末达到全年最高(24种和12种);其次是蝶形花科和十字花科(Brassicaceae),前者从4月末到10月初基本保持平稳(9—11种),而后者在4月末物种数达到最高(9种)。各生境中主要科所含属数和物种数见表1。可以看出,菊科、禾本科、蝶形花科、十字花科等的物种在各生境中分布均较为广泛,而蓼科(Polygonaceae)则在林地(WL)和水边(WS)属数及物种数最高,分别为3属7种和3属6种。
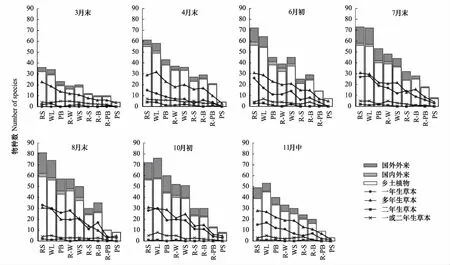
图2 物种来源及生活型的时空变化Fig.2 Temporal and spatial change characteristics of species origin and life formRS:路边,Roadside;WL:林地,Woodland; PB:公园边缘,Park boundary;R-W:园路与水体之间,Between road and water;WS:水边,Waterside;R-S:园路与广场之间,Between road and square;R-B:园路与建筑之间,Between road and building;R-PB:园路与公园边缘之间,Between road and park boundary;PS:铺装广场,Paved square
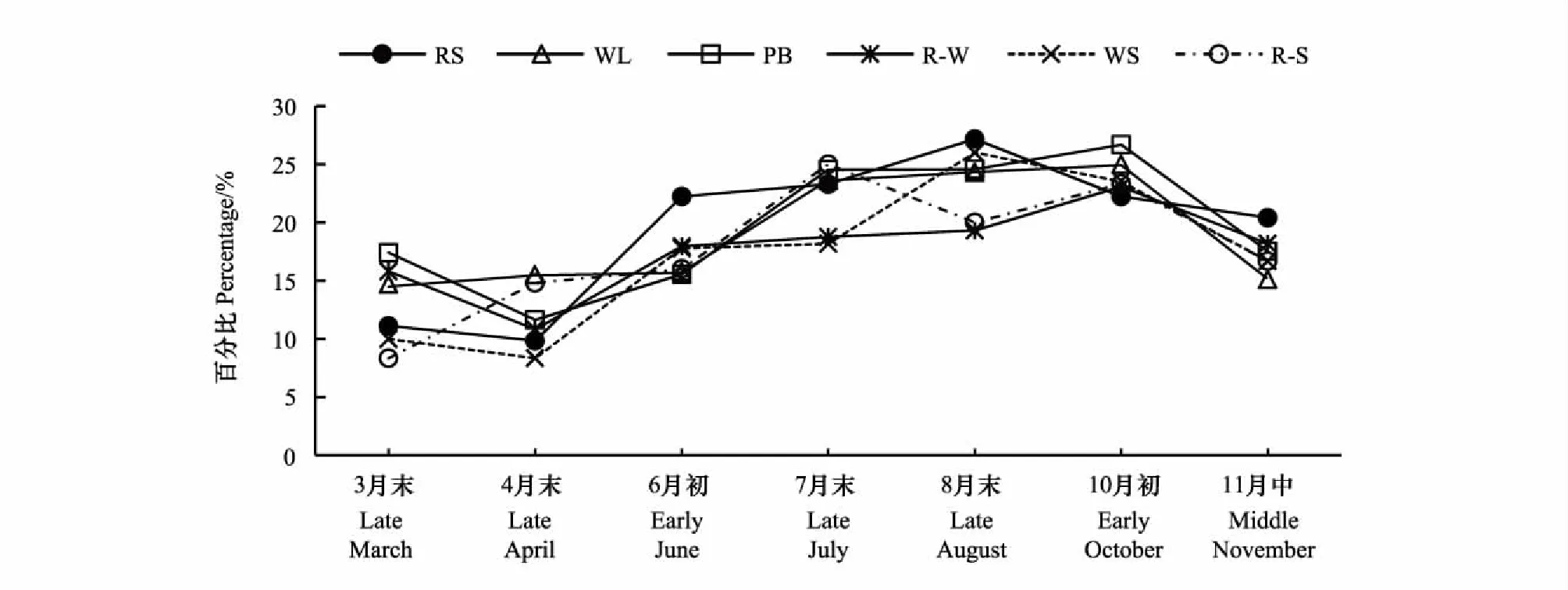
图3 外来植物物种数比例的时空变化Fig.3 Temporal and spatial variation of the percentage of alien species
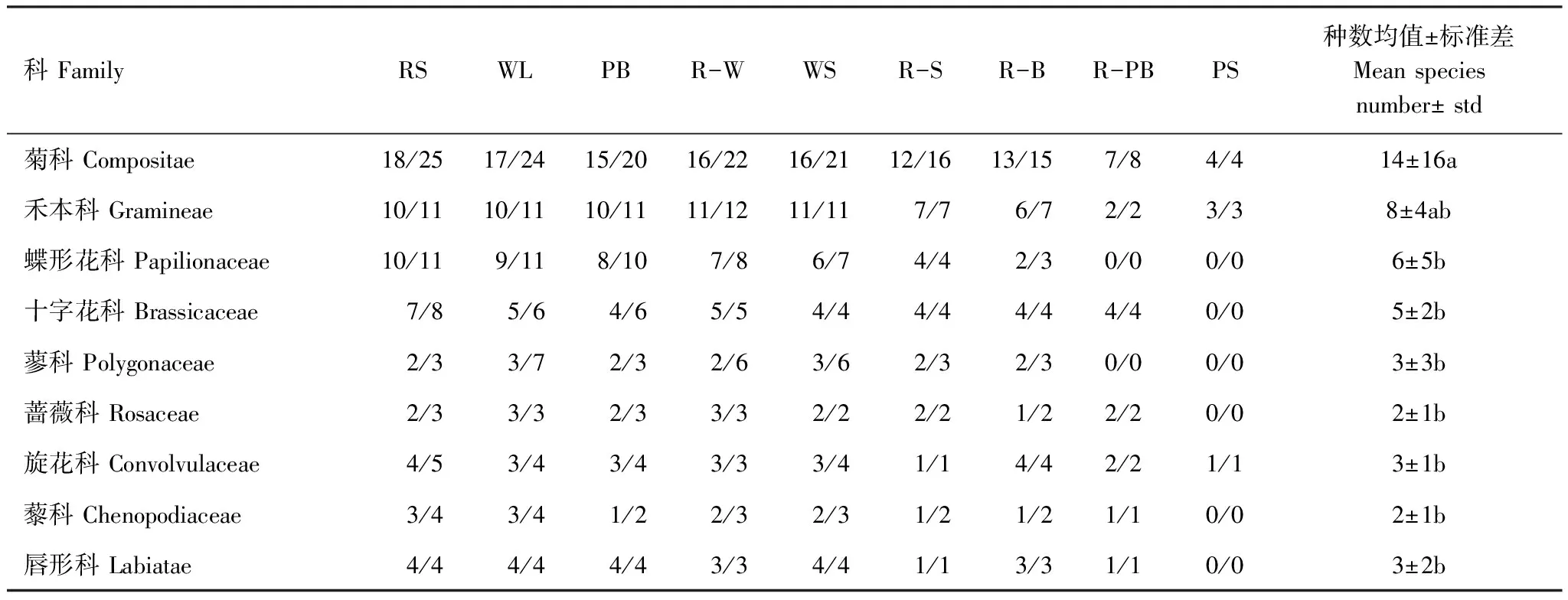
科FamilyRSWLPBR-WWSR-SR-BR-PBPS种数均值±标准差Meanspeciesnumber±std菊科Compositae18/2517/2415/2016/2216/2112/1613/157/84/414±16a禾本科Gramineae10/1110/1110/1111/1211/117/76/72/23/38±4ab蝶形花科Papilionaceae10/119/118/107/86/74/42/30/00/06±5b十字花科Brassicaceae7/85/64/65/54/44/44/44/40/05±2b蓼科Polygonaceae2/33/72/32/63/62/32/30/00/03±3b蔷薇科Rosaceae2/33/32/33/32/22/21/22/20/02±1b旋花科Convolvulaceae4/53/43/43/33/41/14/42/21/13±1b藜科Chenopodiaceae3/43/41/22/32/31/21/21/10/02±1b唇形科Labiatae4/44/44/43/34/41/13/31/10/03±2b
不同小写字母表示同一月份不同生境间差异显著(P<0.05);RS:路边,Roadside;WL:林地,Woodland; PB:公园边缘,Park boundary;R-W:园路与水体之间,Between road and water;WS:水边,Waterside;R-S:园路与广场之间,Between road and square;R-B:园路与建筑之间,Between road and building;R-PB:园路与公园边缘之间,Between road and park boundary;PS:铺装广场,Paved square
2.3 自生植物群落多样性的时空变化
各月份所有生境Shannon-Wiener多样性指数的值域范围以及各生境平均值排序如图4。总体上,Shannon-Wiener多样性指数值位于1.000—14.283之间,各月份间差异性极显著(P<0.001)。10月初群落多样性显著高于其他月份,Shannon-Wiener指数均值为7.24;其次是8月末、4月末和6月初,指数均值分别为6.58、6.13和6.09;而3月末多样性最低,均值仅为3.50(图4)。10月初和8月末,春季发芽、夏秋开花的一年生植物(如禾本科、藜科和蓼科)与多年生植物交相辉映;4月末和6月初则是二年生和秋冬季发芽、翌年春季开花的一年生植物(如十字花科)达到生长的顶峰,因此这四个月份的多样性指数在各生境下均较高,各群落更为稳定,资源分配更均匀。整个生长季中各生境多样性指数均值排序变化较大,路边(RS)和林地(WL)多样性指数值域较广,并且在各月份均呈现出RS > WL的特征,且在8月末差异显著;水边(WS)的多样性于8月末和10月初达到高峰,其值域范围也变得更小。Duncan检验表明各生境的Shannon-Wiener多样性指数差异的显著性随着时间的变化表现出一定的差异,8月末、11月中各生境多样性差异显著,而其他月份各生境的多样性基本上差异不显著且展现出多变的特点。
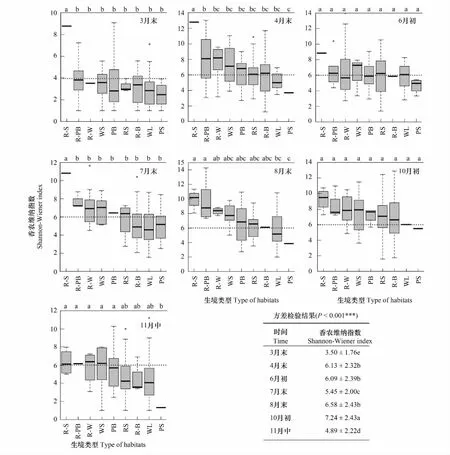
图4 群落多样性的时空变化Fig.4 Community diversity variation of temporal and habitats distribution不同小写字母表示不同生境及月份间差异显著(P<0.05)
2.4 公园环境下自生植物的群落分类
本着自生植物群落应能反映季相特点及其与生境之间的联系,并更加适宜园林绿地应用的原则,以季节为一级分类指标,生境为二级指标,以物种的简化优势度为依据进行群落聚类;将公园环境下7次调研样方数大于5的5种生境下787个(剔除物种盖度<5%的样方)群落分为春季群落(3、4月)、夏季群落(6、7、8月)和秋季群落(10、11月),通过组间Pearson相关系数聚类法在距离20处共得到群落组42个,分别以优势种进行命名,其所属的季节、生境和该生境下所占比例及主要群落(优势种+亚优势种)见表2。
春季群落共包括22个群落组,优势群落组为二月蓝、抱茎苦荬菜、附地菜,属于单一生境的有13个,如路边的斑种草(Bothriospermumchinense)、园路与水体之间的巴天酸模(Rumexpatientia)等;夏季群落共包括22个群落组,优势群落组为狗尾草、早开堇菜、紫苜蓿,属于单一生境的有9个,如路边的牛膝菊、水边的鼠掌老鹳草(Geraniumsibiricum)等;秋季群落共包括26个群落组,优势群落组为二月蓝、蒲公英、抱茎苦荬菜,而属于单一生境的群落组有17个,如林地的紫苜蓿、公园边缘的附地菜等。三季均被划分出的群落组有9个,如抱茎苦荬菜、紫苜蓿、旋覆花(Inulajaponica)等。
3 讨论
3.1 城市公园环境下自生植物的物种组成特征及启示
从前人研究结果来看,相比西安城市建成区自生植物82种[13]、上海中心城区的107种[18-19]以及宁波城市区域的127种[31],研究范围更小的奥林匹克森林公园自生植物物种数达到128种,表明北京市自生植物比较丰富,且公园绿地是自生植物重要的栖息地之一。研究还发现,奥林匹克森林公园中菊科和禾本科是物种数最多的两大优势科,与哈尔滨、上海、宁波等地区在优势科上差异不大,同样以菊科和禾本科为主[16-19,31]。从优势种来看,奥森公园以狗尾草、蒲公英、抱茎苦荬菜为主,与前人研究的北方地区不同用地类型中优势种相似,而与南方地区差异明显,如狗牙根、香附子(Cyperusrotundus)等在南方较北方地区更具优势[31-32],这些物种在植物景观地域特色营造中具有重要作用。虽然气候条件会影响城市自生植物的物种组成[33],但也有一些物种适应性极强,如狗尾草、蒲公英、草地早熟禾等,在全国及国外地区都有广泛的分布[34-35],这类广布种在环境贫瘠的场地中可作为建群种考虑。从物种来源来看,本研究中乡土植物的比例约为76.56%,远远高于北京市建成区所有植物种类中乡土植物的比例(47%)[36],表明自生植物中有较为丰富的可应用的乡土植物资源,许多具有较高观赏及生态价值的物种尚有待开发。外来植物的物种数和所占比例与物种总数在时间上的变化一致,均在夏季达到高峰,部分是由于外来植物多来自菊科(比例为29.17%),而菊科多为一年生或夏季开花的多年生植物,在夏季达到生长高峰,如小蓬草、牛膝菊等。另外,有研究表明丰富的热量、充足的水分、稳定的气候和多样的生境更有利于外来植物的入侵和扩散[21],因此,夏季应注意控制外来物种的蔓延。在生活型组成方面,总体上一、二年生物种数高于多年生,这与前人在城市不同环境中的研究结果一致[16-19],表明了公园中的外来干扰导致对于自生植物很难具备从一年生演替到多年生阶段的稳定生境,因此在今后城市公园绿地建设中应引起一定的重视,在规划时合理预留低干扰的自生植物栖息生境。一、二年生与多年生植物在时间上的变化特征与其生活史密切相关,在未来规划设计时应选择多样的生活型进行合理搭配,达到一、二年生与多年生植物更充分地利用资源的同时形成丰富的季相。

表2 各季节下群落组所属生境及其在该生境中的比例
3.2 自生植物群落多样性的时空特征及影响因素
已有研究表明景观多样性、异质性与植物多样性正相关[37],但本研究结果却与此不尽相同。由于公园游客量较大再加之养护管理工作的影响,各生境群落多样性的时空特征呈现出较为复杂的动态变化。在空间格局上,水边群落主要以禾本科、蓼科、菊科植物为主,大部分为夏季开花的一年生或多年生植物,因此在10月初和8月末多样性达到最高;路边边缘效应产生了一定的影响,自生植物群落多样性在各时间段均高于林地;公园边缘、园路与水体之间等生境由于边缘效应也表现出较高多样性,而园路与广场之间、铺装广场由于硬质场地大、人为干扰高,群落多样性较低。在时间变化上,由于优势种在各类生境中广泛分布,不同生境的群落多样性仅在8月末和11月中表现出差异,推测这可能是因为特定植物在特定生境中生长时间的分化,如蓼科植物夏季生长旺盛并倾向生长于水边。此外,林地、路边的夏季除草等人为干扰也可能对群落多样性产生了一定的影响。优势种在各类生境中的广泛分布同时也导致了各生境类型之间的群落聚类所得群落组存在较高的相似性,如狗尾草和二月蓝,几乎在所有生境中都占有较高比例,因此若要增加景观多样性,可有针对性地对这些优势种进行适度的干预。春、夏、秋季,属于单一生境的群落组分别有13、9、17个,表明在特定的季节这些群落组在特定的生境下更具优势、对该生境更为适应,这些群落组及物种组合为以自生植物为主的低维护景观规划、保护和应用提供了重要依据。植物群落多样性水平与其生态效益的发挥密切相关[38],10月初及8月末、4月末多样性较高的群落为构建多样、稳定的低维护植物景观提供了一定的参考,同时群落多样性的时空格局变化特征反映了公园环境下自生植物群落多样性的维持机制,如生境的环境条件及人为干扰等因素的影响,未来还需针对各影响因子进行更为细致深入的研究以为城市绿地低维护景观规划及管理提供更多科学依据。
3.3 自生植物的群落动态及观赏特征
自生植物群落随着时空变化呈现出一定的动态,也体现着较高的观赏价值。春季开花植物形成了以蓝紫色系(如二月蓝、早开堇菜、刺儿菜等群落)、白色系(如夏至草、点地梅(Androsaceumbellata)、糙叶黄芪等群落)和黄色系(白屈菜、蒲公英、抱茎苦荬菜等群落)野花灿烂点缀的主旋律;夏季是以红色系观花(如圆叶牵牛、打碗花、益母草等群落)和绿色展叶植物(如车前、铁苋菜、葎草等群落)形成的锦绣二重唱;秋季各植物开始枯萎和衰败,则呈现出以红紫色、黄褐色叶片以及果穗混合而成的低婉交响曲。因环境条件不同,不同生境有其特定的优势种、偶见种及群落组合,因而也呈现出不同的景观效果。林地蕴含丰富的物种资源,但由于人为干扰的影响植株高度普遍较低,艾蒿、野艾蒿、活血丹、夏至草和三齿萼野豌豆为林地特有群落组;路边群落聚集度较高,以狗尾草(夏季45.2%)、牛筋草(夏季10.3%)、车前(夏季7.1%)等群落组形成的绿色条带或二月蓝(春季27.1%)、附地菜(春季15.3%)、抱茎苦荬菜(春季15.3%)等群落组形成的繁花小境为主,斑种草、酢浆草、牛膝菊等为特有群落组;水边的群落组体量较大、繁茂紧凑,野趣盎然更充满活力,狗尾草(夏季55.6%)、二月蓝(秋季54.5%)、旋覆花(秋季45.5%)、紫苜蓿(夏季22.2%)较具优势,仅独行菜群落组为特有;公园边缘干扰低,荒野气息十足,呈现出以狗尾草(夏季52.1%)、二月蓝(春季45.2%)、早开堇菜(夏季31.3%)群落组为优势的格局,而打碗花、蛇含、阿拉伯婆婆纳3个群落组为特有;园路和水体之间则以蒲公英(秋季33.3%)、狗尾草(夏季29.6%)、抱茎苦荬菜(秋季27.3%)群落组更具优势,巴天酸模、葎草、大画眉草、菵草4个为特有群落组。研究表明生境多样性是决定乡土植物丰富度的关键因素[3],保护生物多样性的根本就是保持和维护乡土生物与生境的多样性[39],因此,低干扰生境的营造以及不同生境下植物群落的搭配十分重要,这些不同生境类型下的优势群落组、特有群落组和物种组合模式随着时间变化展现出丰富多彩的景观效果,在未来郊野公园、森林公园及生态修复项目中具有广泛的应用前景。
4 结论
奥林匹克森林公园自生植物种类较为丰富,本次共调查到128种。由于人为干扰的影响,一年生植物较多,外来植物表现出夏季比例增多的特点;群落多样性在春、夏季最高,而各生境下群落多样性相对不稳定;全年群落共被划分成了42个群落组,不同季节和生境下具有不同的分配与组合并且展现出各自独特的景观效果。利用自生植物营建低维护景观,不仅可以增添自然野趣,同时节约人力、物力以及资源的消耗。在未来可持续、低维护绿地植物景观规划设计中,应根据自生植物在特定时空中的物种组成、多样性、景观表现合理进行群落营造,并有针对性地采取养护管理措施。
[1] Sudha P, Ravindranath N H. A study of bangalore urban forest. Landscape and Urban Planning, 2000, 47(1/2): 47- 63.
[2] Smart S M, Bunce R G H, Marrs R, LeDucc M, Firbanka L G, Maskella L C, Scotta W A, Thompsond K, Walkere K J. Large-scale changes in the abundance of common higher plant species across Britain between 1978, 1990 and 1998 as a consequence of human activity: tests of hypothesised changes in trait representation. Biological Conservation, 2005, 124(3): 355- 371.
[3] 侯冰飞, 贾宝全, 冷平生, 王文和. 北京市城乡交错区绿地和植物种类的构成与分布. 生态学报, 2016, 36(19): 6256- 6265.
[4] Woodward S L. Spontaneous vegetation of the Murray springs area, San Pedro valley, Arizona. Journal of the Arizona Academy of Science, 1972, 7(1): 12- 16.
[5] Jaritz G. Manurial experiments on fallows of long or several years′ duration with spontaneous pasture vegetation in North-West-Tunisia and their importance for elaborating measures for pasture improvement. Zeitschrift Fuer Acker und Pflanzenbau, 1974, 139(4): 273- 306.
[6] Sonneveld I S. Classification and evaluation of forest, also with the aid of the spontaneous vegetation. Nederlands Bosbouwtijdschrift, 1977, 49(2): 44- 65.
[7] Choi Y D. Theories for ecological restoration in changing environment: toward ‘Futuristic’ restoration. Ecological Research, 2004, 19(1): 75- 81.
[8] Sagoff M. Do non-native species threaten the natural environment? Journal of Agricultural and Environmental Ethics, 2005, 18(3): 215- 236.
[9] Luliana P, Adelina D, Valentin S, Doina C, Georgel M. Ecological and aesthetic role of spontaneous flora in urban sustainable landscapes development. Journal of Plant Development, 2011, 18: 169- 177.
[10] Boechat C L, Pistóia V C, Gianelo C C, de Oliveira Camargo F A. Accumulation and translocation of heavy metal by spontaneous plants growing on multi-metal-contaminated site in the Southeast of Rio Grande do Sul state, Brazil. Environmental Science and Pollution Research, 2016, 23(3): 2371- 2380.
[11] Cavalca L, Corsini A, Canzi E, Zanchi R. Rhizobacterial communities associated with spontaneous plant species in long-term arsenic contaminated soils. World Journal of Microbiology and Biotechnology, 2015, 31(5): 735- 746.
[12] Wang J, Zhang C B, Ke S S, Qian B Y. Different spontaneous plant communities in Sanmen Pb/Zn mine tailing and their effects on mine tailing physico-chemical properties. Environmental Earth Sciences, 2011, 62(4): 779- 786.
[13] Cervelli E W, Lundholm J T, Du X. Spontaneous urban vegetation and habitat heterogeneity in Xi′an, China. Landscape and Urban Planning, 2013, 120: 25- 33.
[14] Kühn N. Intentions for the unintentional——spontaneous vegetation as the basis for innovative planting design in urban areas. Journal of Landscape Architecture, 2006, 1(2): 46- 53.
[15] 褚建君. 杂草科学的发展与度规的变革. 杂草科学, 2002, (3): 1- 7.
[16] 陈晓双, 梁红, 宋坤, 达良俊. 哈尔滨城区杂草群落多样性及其分类体系. 应用生态学报, 2014, 25(8): 2221- 2228.
[17] 陈晓双, 梁红, 宋坤, 达良俊. 哈尔滨中心城区杂草物种多样性及其在异质生境中的分布特征. 生态学杂志, 2014, 33(4): 946- 952.
[18] 田志慧, 陈克霞, 达良俊, 古红梅. 城市化进程中上海植被的多样性、空间格局和动态响应(Ⅲ)——高度城市化影响下上海中心城区杂草区系特征. 华东师范大学学报: 自然科学版, 2008, (4): 49- 57.
[19] 田志慧, 蔡北溟, 达良俊. 城市化进程中上海植被的多样性、空间格局和动态响应(Ⅷ)——上海乡土陆生草本植物分布特征及其在城市绿化中的应用前景. 华东师范大学学报: 自然科学版, 2011, (4): 24- 34.
[20] 姚和金, 赵建华, 姚宏昌, 万光龙, 叶飞. 白三叶草对马尼拉草坪杂草群落及其多样性的影响. 南京林业大学学报: 自然科学版, 2014, 38(4): 75- 80.
[21] 冯建孟, 董晓东, 徐成东. 中国外来入侵植物物种多样性的空间分布格局及与本土植物之间的关系. 西南大学学报: 自然科学版, 2010, 32(6): 50- 57.
[22] 吴海荣, 强胜. 南京市秋季外来杂草定量调查研究. 生物多样性, 2003, 11(5): 432- 438.
[23] 牛建忠, 刘育俭, 李红云, 张卉, 王艳. 天坛公园野生草地持续利用与管理. 北京园林, 2013, 29(4): 47- 52.
[24] 李文, 吕秀娟, 李树华. 清华校园春季野生草本地被植物多样性与群落分类. 东北林业大学学报, 2010, 38(8): 31- 33.
[25] 张金屯. 数量生态学. 北京: 科学出版社, 2004: 19- 19, 123- 125.
[26] 张金龙, 马克平. 种间联接和生态位重叠的计算: SPAA程序包//马克平编. 中国生物多样性保护与研究进展X. 北京: 气象出版社, 2014: 165- 173.
[27] 贺士元, 邢其华, 尹祖棠, 江先甫. 北京植物志. 北京: 北京出版社, 1993.
[28] 中国科学院中国植物志编辑委员会. 中国植物志. 北京: 科学出版社, 2002.
[29] 刘全儒, 于明, 周云龙. 北京地区外来入侵植物的初步研究. 北京师范大学学报: 自然科学版, 2002, 38(3): 399- 404.
[30] 杨景成, 王光美, 姜闯道, 赵洪涛, 张志东. 城市化影响下北京市外来入侵植物特征及其分布. 生态环境学报, 2009, 18(5): 1857- 1862.
[31] 赵娟娟, 孙小梅, 陈珊珊, 江波, 陈春娣. 城市野生草本植物种类构成的特征——以宁波市为例. 生态环境学报, 2016, 25(1): 43- 50.
[32] 杨剑, 卢昌义, 于兴娜. 深圳市草坪杂草发生季节变化及杂草群落聚类分析. 武汉植物学研究, 2006, 24(6): 518- 524.
[33] Satrapová J, Hyvönen T, Venclová V, Soukup J. Growth and reproductive characteristics of C4 weeds under climatic conditions of the Czech Republic. Plant, Soil and Environment, 2013, 59(7): 309- 315.
[34] Melander B, Holst N, Grundy A C, Kempenaar C, Riemens M M, Verschwele A, Hansson D. Weed occurrence on pavements in five North European towns. Weed Research, 2009, 49(5): 516- 525.
[35] Fagot M, De Cauwer B, Beeldens A, Boonen E, Bulcke R, Reheul D. Weed flora in paved areas in relation to environment, pavement characteristics and weed control. Weed Research, 2011, 51(6): 650- 660.
[36] Zhao J J, Ouyang Z Y, Zheng H, Zhou W Q, Wang X K, Xu W H, Ni Y M. Plant species composition in green spaces within the built-up areas of Beijing, China. Plant Ecology, 2010, 209(2): 189- 204.
[37] Cornelis J, Hermy M. Biodiversity relationships in urban and suburban parks in Flanders. Landscape and Urban Planning, 2004, 69(4): 385- 401.
[38] 潘声旺, 袁馨, 雷志华, 胡明成. 乡土植物生活型构成对川渝地区边坡植被水土保持效益的影响. 生态学报, 2016, 36(15): 4654- 4663.
[39] 俞孔坚, 李迪华, 吉庆萍. 景观与城市的生态设计: 概念与原理. 中国园林, 2001, 17(6): 3- 10.

附表1 北京奥林匹克森林公园自生植物调研名录
*入侵植物
