8个稻瘟病抗性基因在三系杂交粳稻亲本中的分布
2018-03-07储黄伟牛付安周继华涂荣剑罗忠永王新其曹黎明
储黄伟,程 灿,牛付安,周继华,涂荣剑,罗忠永,王新其,曹黎明
(上海市农业科学院作物育种栽培研究所,上海201403)
水稻是我国最主要的粮食作物,有超过65%的人口以水稻为主粮。由稻瘟病菌(Magnaporthe oryzae)引起的稻瘟病是水稻的主要病害之一。稻瘟病的爆发,会严重影响水稻的产量,甚至导致部分病害严重区域绝收[1]。在所有的稻瘟病防治措施中,稻瘟病抗病品系的培育和种植被认为是最有效、经济和安全的控制该病害的措施[2-3]。
传统的稻瘟病抗性育种,依赖于杂交和表型的选择,存在育种周期长,工作量大,准确率低等问题。近年来,随着分子生物学的发展,利用和已知稻瘟病抗性基因紧密连锁的分子标记进行分子标记辅助育种,逐渐成为稻瘟病抗性育种的主要手段。目前,至少已报道了70个稻瘟病抗性位点共85个主效基因[4]。在这些稻瘟病主效抗性基因中,已经有 Pb1、Pia、Pib、Pid2、Pid3、Pik、Pik-h/Pi54、Pik-m、Pik-p、Pish、Pit、Pita、Piz-t、Pi1、Pi2、Pi5、Pi9、pi21、Pi25、Pi36、Pi37、Pi56、Pi63和 PiCO39等 24个基因被成功地克隆(国家水稻数据中心,稻瘟病主效抗性基因列表http://www.ricedata.cn/gene/gene_pi.htm)。根据已经克隆的稻瘟病抗性基因与其感病等位基因之间的序列差异而设计的功能性标记,可以不用费时费力地接菌鉴定,而准确和快速地检测到水稻品系抗稻瘟病基因的存在[5],这为鉴定携带稻瘟病抗性基因的种质资源以及开展抗稻瘟病的聚合育种带来了新的契机。
在上海地区,每年的水稻种植面积大约在10万hm2左右,在这其中杂交粳稻的面积大约占1/3左右。然而,对于目前在上海地区使用的杂交粳稻骨干亲本中稻瘟病抗性基因的基因型,还没有进行鉴定。本研究利用 Pi2[6-8]、Pi5[6,9]、Pi9[6,10]、Pi54[11-13]、Pia[14-15]、Pib[16-18]、Pit[5]、Pita[6,19]等 8个稻瘟病抗性基因的功能性标记,分析这8个抗稻瘟病基因在36份恢复系和42份不育系中的分布,以期为配制携带有不同稻瘟病抗性基因的杂交水稻组合,以及利用标记辅助选择聚合多个稻瘟病抗性基因品系的培育奠定基础。
1 材料与方法
1.1 水稻品系
供试的36份恢复系和42份不育系材料(表1)均来自上海市农业科学院作物育种栽培研究所水稻杂种优势利用课题组,是课题组选育杂交粳稻的骨干亲本。
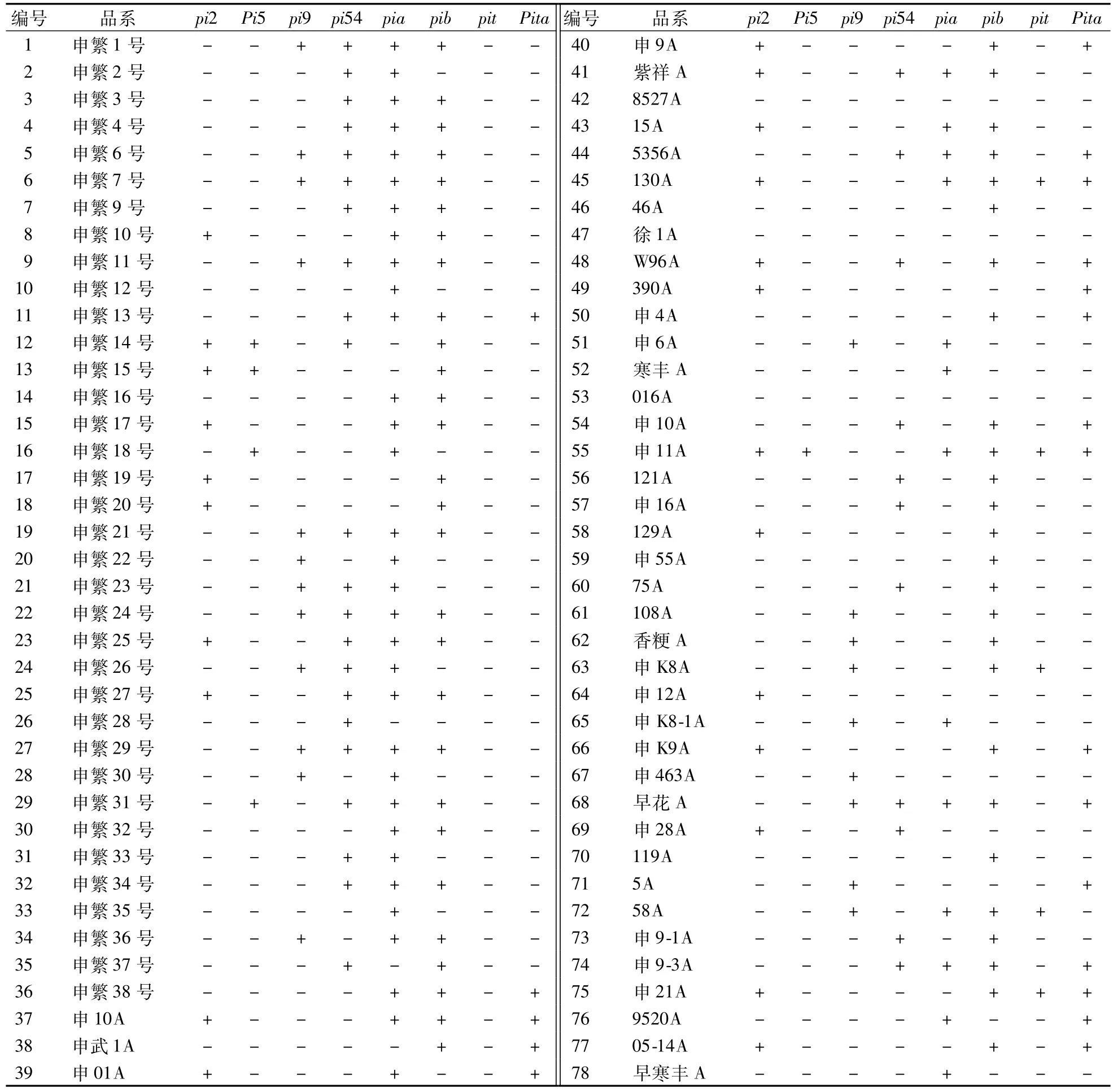
表1 水稻品系中稻瘟病抗性基因的基因型检测Table 1 Genotypic detection of blast-resistant genes in rice lines
1.2 水稻DNA提取
每个水稻品系取50 mg左右叶片,用剪刀剪碎,放入2 mL离心管,加入600μL 1.5×CTAB溶液(1.5%CTAB,75 mmol/L Tris-HCl,15 mmol/L EDTA,1.05 mol/L NaCl,pH 8.0)和1粒直径5 mm的钢珠,在快速研磨仪上70 Hz的频率震荡研磨90 s;研磨后的样品在56℃水浴锅中温浴20 min,加入450μL氯仿,剧烈震荡后,12 000 r/min离心10 min;取450μL上清液到另一个1.5 mL的离心管中,加入900μL的无水乙醇,混匀后在-20℃冰箱放置10 min,12 000 r/min离心10 min,弃上清,晾干后加入200μL的双蒸水溶解DNA,置于-20℃冰箱备用。
1.3 稻瘟病抗性基因的检测
本研究检测 Pi2[6-7]、Pi5[6,9]、Pi9[6,10]、Pi54[11-13]、Pia[14-15]、Pib[16-18]、Pit[5]、Pita[6,19]等 8个稻瘟病抗性基因,其功能性分子标记的引物信息见表2。PCR扩增以及产物检测的方法参照表2中相应的文献。

表2 PCR扩增所用的引物Table 2 Primers used for PCR amplification
1.4 数据统计分析
根据PCR产物的电泳检测结果,检出稻瘟病抗性等位基因记为1,检出感病等位基因记为0。然后利用NTsys 2.1e软件中的SHAN子集的UPGMA方法进行聚类分析。
2 结果与分析
2.1 稻瘟病抗性基因的检测
检测了36个恢复系和42个不育系中Pi2、Pi5、Pi9、Pi54、Pia、Pib、Pit和Pita等8个稻瘟病抗性基因的基因型(表1)。结果表明:Pib稻瘟病抗性基因在本研究检测的水稻品系中分布最为广泛,在54个(28个不育系和26个恢复系)品系中检测到了抗性基因的存在;其次从高到低依次为Pia、Pia54、Pi9、Pi2和Pita,分别在45个(15个不育系和30个恢复系)、33个(11个不育系和22个恢复系)、24个(12个不育系和12个恢复系)、23个(15个不育系和8个恢复系)和20个(18个不育系和2个恢复系)品系中检测到;而Pi5和Pit是在本研究检测的水稻品系中出现频率最低的稻瘟病抗性基因,分别只在5个品系中检测到,其中Pi5抗性基因在1个不育系和4个恢复系中,而含有Pit抗性基因的5个品系都是不育系(图1)。
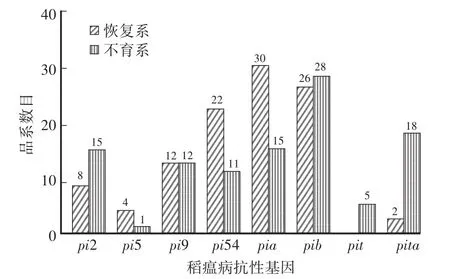
图1 稻瘟病抗性等位基因在检测水稻品系中的分布频率Fig.1 Frequency distribution of blast-resistant genes in rice lines
在所有检测的78个水稻品系中,在8527A、徐1A和016A等3个品系中未检测到稻瘟病抗性基因的存在,而含有稻瘟病抗性基因最多品系申11A含有6个抗性基因,其余含有1—5个抗性基因的分别有10个、25个、18个、19个和2个水稻品系(图2)。

图2 稻瘟病抗性等位基因数目在检测水稻品系中的分布频率Fig.2 Frequency distribution of blast-resistant gene numbers in rice lines
2.2 遗传差异聚类分析
根据8个稻瘟病抗性基因的基因型特征,对78个水稻品系进行聚类分析。结果表明:检测的水稻品系间遗传相似系数的变化范围为0.55—1,在相关系数大于0.67的水平上,可以将78个水稻品系分为5组,其中组Ⅰ包含31个品系,组Ⅱ包含4个品系,组Ⅲ包含20个品系,组Ⅳ包含13个品系,组Ⅴ包含10个品系(图3)。

图3 78个水稻品系遗传差异聚类分析Fig.3 Genetic distance-based cluster analysis of 78 rice lines
3 讨论
在目前已经报道的85个稻瘟病抗性主效基因中,除了Pi21[20]和 Pi55[21]两个是隐性基因,其他的稻瘟病抗性等位基因都是显性的(国家水稻数据中心,稻瘟病主效抗性基因列表http://www.ricedata.cn/gene/gene_pi.htm)。解释杂种优势形成机理的“显性假说”认为在等位基因中多数显性基因是有利的,相对的隐性基因是不利的。在杂交后代中来自一个亲本的显性等位基因可以掩盖来自另一个亲本的隐性等位基因,使得在杂交后代中具有比单个亲本更多的有利显性基因,从而使杂合后代表现出更好的生长、抗逆和抗病等等性状[22-23]。根据本研究稻瘟病抗性基因的鉴定和遗传差异的聚类分析结果,在配制杂交组合亲本选择的过程中,可以按照稻瘟病抗性基因的互补及远缘杂交优势的原则进行。根据这一原则,用不育系申9A分别与恢复系申繁24号和申繁26号配制了2个杂交组合申优415和申优26。在遗传差异的聚类分析结果中,申9A、申繁24号和申繁26号分别属于组Ⅴ、组Ⅰ和组Ⅲ,申9A含有P i2、Pib和Pita等3个抗性基因,申繁24号含有Pi9、Pi54、Pia和Pib等4个抗性基因,而申繁26号含有Pi9、Pi54和Pia等3个抗性基因,这样就使得杂交组合申优415和申优26都含有了Pi2、Pi9、Pi54、Pia、Pib和Pita等6个稻瘟病抗性基因。在这6个基因中,Pi2和Pi9是同一个基因位点上的复等位基因[24]。Pi2对稻瘟病的抗谱相当广,将携带Pi2的亲本C101A51接种从华东、华南和西南12个省区收集来的792个稻瘟病小种,对其中92.45%的小种表现出抗性[25],此外Pi2还对来自13个国家的43个稻瘟病菌株中的36个表现抗性[24];Pi9的抗性同样很广,对来自13个国家的43个稻瘟病菌株均表现出很高的抗性[24];Pia抗性基因的抗谱较窄,对来自于中国10个省区的稻瘟病群体接种鉴定,仅对来自江苏的稻瘟病群体表现出较强的抗[15];Pib基因对来自日本的大多数稻瘟病小种以及中国的菌株ZB13和ZC15表现出较好的抗性[16];Pi54是一个广谱的稻瘟病抗性基因,对来自印度的大多数稻瘟病小种表现出抗性[13];Pita对稻瘟病菌菌株ZN57(IC-17)和ZN61(IB-49)表现出较好的抗性[26]。通过亲本的选择,使最终用于生产的杂交组合申优415和申优26中都含有了6个稻瘟病抗性主效基因,保证了这2个组合具有了比较广谱的稻瘟病抗性,这2个组合在上海及周边地区生产试验中表现出较强的稻瘟病抗性。
本研究检测的8个稻瘟病抗性基因中,Pi5和Pit这2个基因的分布较少,Pi5在4个恢复系和1个不育系中存在,而Pit只在5个不育系中检测到。对于这类基因,可以应用分子标记辅助选择的手段转入到更多的水稻品系中。
检测结果还发现8527A、徐1A和016A等3个不育系中不含本研究中检测的8个稻瘟病抗性基因中的任何一个,另外还有10个品系中只检测到了一个稻瘟病抗性基因。在应用这类水稻品系配制杂交组合时,应尽量选择与含有多个稻瘟病抗性基因的品系进行配组。然而在杂交水稻制种的过程中,选用这类稻瘟病抗性较差的品系作为亲本,存在着一定的风险,今后将考虑应用标记辅助选择的手段,将稻瘟病抗性基因引入这些品系。
目前已知的稻瘟病主效抗性基因已经有85个,已经克隆的也有24个,而本研究中只检测了8个稻瘟病抗性基因。在今后的工作实践中,有必要去检测更多的稻瘟病抗性基因在本课题组核心种质资源中的分布,为杂交组合的配制及分子标记辅助选择育种提供理论基础。
[1]OU SH.Rice diseases[M].Kew,UK:Commonwealth Mycological Institute,1985.
[2]FUKUOKA S,SAKA N,KOGA H,et al.Loss of function of a proline-containing protein confers durable disease resistance in rice[J].Science,2009,325(5943):998-1001.
[3]JEUNG JU,KIM B R,CHO Y C,et al.A novel gene,Pi40(t),linked to the DNA markers derived from NBS-LRR motifs confers broad spectrum of blast resistance in rice[J].Theor Appl Genet,2007,115(8):1163-1177.
[4]ZHENGW,WANG Y,WANG L,etal.Geneticmapping andmolecularmarker development for Pi65(t),a novel broad-spectrum resistance gene to rice blast using next-generation sequencing[J].Theor Appl Genet,2016,129(5):1035-1044.
[5]HAYASHIK,YASUDA N,FUJITA Y,et al.Identification of the blast resistance gene Pit in rice cultivars using functionalmarkers[J].Theor Appl Genet,2010,121(7):1357-1367.
[6]高利军,高汉亮,颜群,等.4个抗稻瘟病基因分子标记的建立及在水稻亲本中的分布[J].杂交水稻,2010(S1):294-298.
[7]ZHOU B,QU S,LIU G,et al.The eight amino-acid differences within three leucine-rich repeats between Pi2 and Piz-t resistance proteins determine the resistance specificity to Magnaporthe grisea[J].Mol Plant Microbe Interact,2006,19(11):1216-1228.
[8]YU Z H,MACKILL D J,BONMAN JM,et al.Tagging genes for blast resistance in rice via linkage to RFLPmarkers[J].Theor Appl Genet,1991,81(4):471-476.
[9]LEE SK,SONG M Y,SEO Y S,et al.Rice Pi5-mediated resistance to Magnaporthe oryzae requires the presence of two coiled-coil-nucleotidebinding-leucine-rich repeat genes[J].Genetics,2009,181(4):1627-1638.
[10]QU S,LIU G,ZHOU B,et al.The broad-spectrum blast resistance gene Pi9 encodes a nucleotide-binding site-leucine-rich repeat protein and is amember of amultigene family in rice[J].Genetics,2006,172(3):1901-1914.
[11]RAMKUMAR G,SRINIVASARAO K,MOHAN KM,etal.Developmentand validation of functionalmarker targeting an InDel in themajor rice blast disease resistance gene Pi54(Pik h)[J].Molecular Breeding,2011,27(1):129-135.
[12]SHARMA TR,RAIA K,GUPTA SK,et al.Broad-spectrum blast resistance gene Pi-k h cloned from rice line tetep designated as Pi54[J].Journal of Plant Biochemistry and Biotechnology,2010,19(1):87-89.
[13]SHARMA T R,MADHAV M S,SINGH B K,et al.High-resolution mapping,cloning and molecular characterization of the Pi-k h gene of rice,which confers resistance to Magnaporthe grisea[J].Molecular Genetics and Genomics,2005,274(6):569-578.
[14]潘庆华,潘金媚,张玉,等.稻瘟病抗性基因Pia-C功能特异性分子标记及其方法与应用:CN102676512A[P/OL].2012-09-19[2016-12-12].http://d.g.wanfangdata.com.cn/Patent_CN201210164522.6.aspx.
[15]曾晓珊,杨先锋,赵正洪,等.稻瘟病抗病基因Pia的抗性分析及精细定位[J].中国科学:生命科学,2011,41(1):70-77.
[16]刘洋,徐培洲,张红宇,等.水稻抗稻瘟病Pib基因的分子标记辅助选择与应用[J].中国农业科学,2008,41(1):9-14.
[17]WANG Z X,YANOM,YAMANOUCHIU,etal.The Pib gene for rice blast resistance belongs to the nucleotide binding and leucine-rich repeat class of plant disease resistance genes[J].Plant J,1999,19(1):55-64.
[18]FJELLSTROM R,CONAWAY-BORMANSC A,MCCLUNG A M,et al.Development of DNA markers suitable formarker assisted selection of three Pi genes conferring resistance tomultiple Pyricularia grisea pathotypes[J].Crop Science,2004,44(5):1790-1798.
[19]JIA Y,JIA Y,WANG Z,et al.Development of dominant rice blast Pi-ta resistance genemarkers[J].Crop Sci.,2002,42(6):2145-2149.
[20]FUKUOKA S,OKUNO K.QTL analysis and mapping of pi21,a recessive gene for field resistance to rice blast in Japanese upland rice[J].Theoretical and Applied Genetics,2001,103(2):185-190.
[21]何秀英,刘新琼,王丽,等.稻瘟病新隐性抗病基因 pi55(t)的遗传及定位[J].中国科学:生命科学,2012,42(2):125-141.
[22]DAVENPORTC B.Degeneration,albinism and inbreeding[J].Science,1908,28(718):454-455.
[23]BRUCE A B.The Mendelian theory of heredity and the augmentation of vigor[J].Science,1910,32(827):627-728.
[24]LIU G,LU G,ZENG L,et al.Two broad-spectrum blast resistance genes,Pi9(t)and Pi2(t),are physically linked on rice chromosome 6[J].Molecular Genetics and Genomics,2002,267(4):472-480.
[25]CHEN H L,CHEN B T,ZHANG D P,et al.Pathotypes of Pyricularia grisea in rice fields of central and southern China[J].Plant Disease,2001,85(8):843-850.
[26]王忠华,贾育林,吴殿星,等.水稻抗稻瘟病基因 Pi-ta的分子标记辅助选择[J].作物学报,2004,30(12):1259-1265.
