不同年限封育对黄土高原典型草原地上植被的影响
2018-03-06赵凌平谭世图樊文娜王占彬王清义河南科技大学动物科技学院河南洛阳47023河南省饲草饲料资源开发与畜禽健康养殖院士工作站河南洛阳47023
赵凌平,白 欣,谭世图,2,樊文娜,王占彬,2,王清义 (.河南科技大学动物科技学院,河南 洛阳 47023;2.河南省饲草饲料资源开发与畜禽健康养殖院士工作站,河南 洛阳 47023 )
暖温性典型草原是黄土高原面积最大的一类草地类型,但由于水土流失严重,也成为了退耕还草、生态环境治理的重点区域[1]。研究表明,围栏封育是一种实现受损草地自我修复、维护草地生态平衡有效且易行的草地管理手段[2],已成为退化草地植被自然恢复的主要措施。封育可以显著改善退化草地环境,提高草地生产力和牧草品质,对草地的生态保护起着重要的作用[3]。关于封育对草地植被物种组成、群落结构、物种多样性、土壤养分和繁殖策略的影响已有大量报道[4-9]。根据草地资源的利用和可持续发展原理,封育时间不应该是无限制的。在长期封育不放牧条件下,枯落物将会在草地上堆积,达到一定厚度时容易阻碍种子进入土壤,还易于发生火灾[10-12]。因此,在对退化草地进行禁牧是必要的,对退化草地植被恢复具有重要意义,但是封育多长时间对退化草地是合适的是研究重点。
许多研究表明,退化草地围封促进了因放牧而被抑制的群落生长,植被恢复较快[6-8]。但封育时间对草地生态系统的影响存在不同的研究结果,甚至有着相矛盾的结论。李强[13]的研究表明,在松嫩平原羊草(Leymuschinensis)草地上,与封育4年的草地相比,封育6年的群落与健康草地群落间的相似性更高。刘美珍等[14]研究指出,浑善达克退化沙地植被的物种多样性、丰富度和生物量随封育年限的增加呈先增加后降低或平稳,呈抛物线的变化趋势。在宁夏盐池封育区,长期封育草地的生物量低于短期封育草地,封育5年的草地物种多样性指数最高[15]。张晶晶[16]对宁夏荒漠草原封育7、6、5、4和3年及未封育地的群落结构特征研究表明,地上植被的盖度、高度、密度和生物量随封育年限的延长而增加。在以往的研究中,封育年限大多集中在10年以下,缺乏长期封育的数据分析,很难准确评估封育年限对草地植被的影响和较适宜的封育年限。大量研究说明了不同的草地类型或不同立地条件,封育年限对草地植被的影响不同。在典型草原,有关封育年限对草地群落演替的研究很少报道,适宜的封育年限还不是很明确。因此,本研究在黄土高原典型草原,不仅研究短期封育(封育5年)对草地植被的影响,还研究长期封育(封育23年和封育32年)对草地植被特征、物种多样性和群落演替动态变化的影响,进一步探讨黄土高原典型草原适宜封育年限,以期丰富黄土高原草地生态系统群落演替相关研究成果,为其经营管理提供理论依据。
1 样地概况与研究方法
1.1 研究区自然概况
研究区设置在宁夏回族自治区固原市东北部45 km处,位于36°13′-36°19′ N, 106°24′-106°28′ E,海拔1 800~2 180 m。该区是黄土高原典型草原带的代表性区域,为温带大陆性半干旱季风气候。年平均温度5 ℃,≥10 ℃的年积温为2 370~2 882 ℃·d。年平均降水量400~450 mm,其中70%以上集中在7-9月。年平均蒸发量1 330~1 640 mm,年平均无霜期为112~140 d。植物主要以旱生和中旱生草本为主,主要为本氏针茅(Stipabungeana)、大针茅(S.grandis)、甘青针茅(S.przewalskyi)和百里香(Thymusmongolicus)等。
1.2 研究方法
地上植被于2015年7月进行调查。在云雾山自然保护区的试验区选取封育5、15、23、32年草地和放牧地(封育0年)作为试验样地。封育前样地土壤理化性质和植被特征相似[17]。在放牧地,放牧强度为中度放牧(4.6个羊单位·hm-2)。在每个样地随机选择5个小区,每个小区间距至少100 m,在每个小区随机设置6个50 cm×50 cm的样方进行地上植被调查。在样方内,测定总盖度、物种分盖度、物种组成、每个物种的个数。先用尺子测定枯落物厚度,然后收集样方内所有枯落物。分种测定地上生物量,从根茎基部齐地面刈割,然后装袋带回实验室。枯落物和植物的地上生物量均是65 ℃烘干至恒重,然后称其干重[18-20]。样地基本情况如表1所列。
1.3 物种多样性计算方法
物种多样性指数[21-23]计算公式如下:
Shannon-Wiener指数(H′):
Simpson多样性指数(D):
Pielou 均匀度指数(E):
式中:S为物种总数,N为样本中所有物种的个体总数,Pi为物种i的个体数占物种个体总数的比例。
1.4 数据处理
采用多元统计软件MVPSversion 3.2对地上植被相似性进行数量生态学分析,用非加权组平均(UPGMA)进行聚类分析,用Jaccard相似性百分比作为距离测度。
Jaccard相似性指数(C):
式中:a为样地A的物种数,b为样地B的物种数,j为样地A和B共有物种的数量。
采用SPSS 16.0软件进行单因素方差 (One-way ANOVA) 分析,比较分析不同封禁年限草地间地上植被的盖度、密度、物种多样性和生物量的差异。用Tukey’s HSD进行多重比较。P<0.05表示差异显著。用除趋势对应分析法(detrended correspondence analysis, DCA)分析地上植被的演替过程。DCA排序采用CANOCO 4.5软件完成。利用多元统计软件MVSP version 3.2对不同封育草地植被相似性进行聚类分析。
2 结果与分析
2.1 封育年限对地上植被群落特征的影响
单因素方差分析结果表明,封育年限对草地的枯落物量、枯落物厚度、盖度、密度、地上生物量和禾草地上生物量影响显著(P<0.05),而对非禾草地上生物量的影响不显著(P>0.05)(表2)。随着封育年限的延长,枯落物量和厚度呈递增趋势,且封育23和32年草地显著高于其他样地;群落盖度呈先增加后降低趋势,在封育23年草地达到最大。物种密度随封育年限的延长呈递减趋势。与放牧地相比,密度在封育5年草地未显著降低,而到封育15年显著降低。随着封育年限的延长,地上生物量和禾草地上生物量均呈先增加后降低的趋势,且均在封育23年草地达到了峰值。
FM:放牧地;FY(5):封育5年草地;FY(15):封育15年草地;FY(23):封育23年草地;FY(32):封育32年草地。
FM: grazed grassland; FY(5): 5-year fenced grassland; FY(15): 15-year fenced grassland; FY(23): 23-year fenced grassland; FY(32): 32-year fenced grassland.
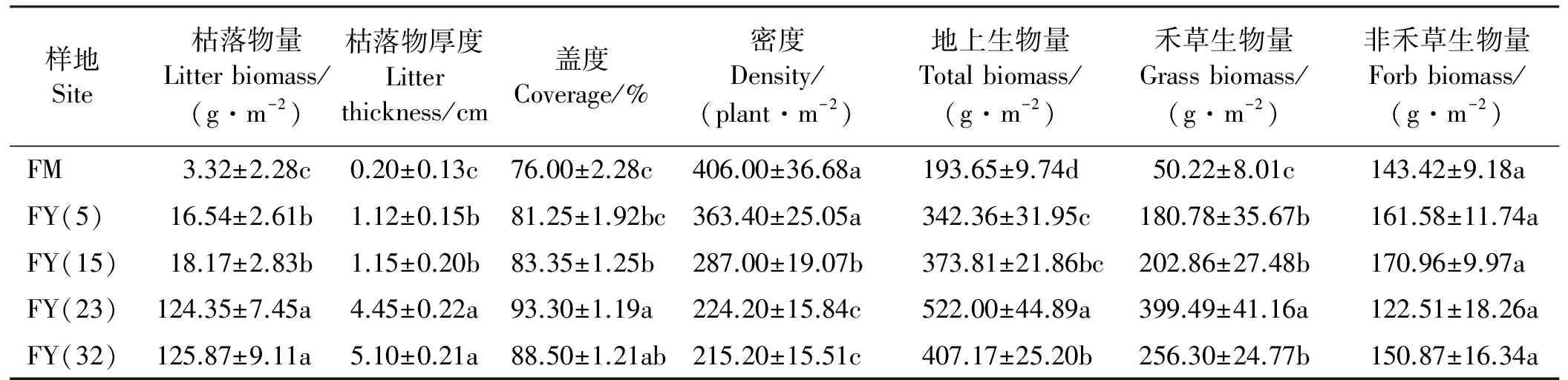
表2 不同封育年限草地的地上植被特征变化Table 2 Changes of aboveground vegetation in grasslands with different enclosure durations
同列不同小写字母表示差异显著(P<0.05),下同。
Different lowercase letters within the same column indicate significant difference at the 0.05 level; similarly for Table 3.
2.2 封育年限对地上植被物种多样性的影响
封育年限对草地的物种丰富度和Shannon-Wiener指数影响显著(P<0.05),而对Simpson指数和Pielou指数影响不显著(P>0.05)(表3)。地上植被的物种丰富度随封育年限的延长呈先增加后降低趋势。封育5年和封育15年草地的物种丰富度显著高于放牧地。物种丰富度最大值出现在封育15草地,到了封育23年又显著降低,且与放牧地差异不显著。与放牧地相比,Shannon-Wiener多样性指数在封育5年草地显著增加,但到了封育32年草地又降到与放牧地同一水平。可见,短期封育能显著提高草地的物种多样性,长期封育反而降低草地的物种多样性。在该区,退化草地封育15年对物种多样性较为有利。
2.3 不同封育年限草地地上植被演替的梯度分析
DCA排序结果表明,第1排序轴明显反映出地上植被群落的演替进程(图1)。由于本研究选取的放牧地的放牧强度为中度放牧,所以放牧地的样方在排序轴上与封育15年的样方处于同一水平。除放牧地外,排序轴从左到右代表着不同封育年限样地地上植被群落演替的方向,即扁穗冰草(Agropyroncristatum)+长叶火绒草(Leontopodiumcalocephalum)-杂类草群落→本氏针茅+白莲蒿(Artemisiasacrorum)-百里香群落→本氏针茅+赖草(Leymussecalinus)-火绒草群落→本氏针茅群落。显然DCA排序演替分析结果正好与实际的演替过程相吻合。由此可见,随着封育年限的延长,该区退化草地群落演替进程为杂草阶段→丛生禾草阶段→本氏针茅群落相对稳定阶段。另外,封育23年与封育32年的草地群落植被之间的相关性最强,即地上植被物种最相似(图1)。

表3 不同封育年限草地物种多样性的变化Table 3 Changes of species diversity in grasslands with different enclosure durations

图1 不同封育年限草地DCA排序Fig. 1 Different enclosure durations ordination plot
数字代表样方号。▼:放牧地;△:封育5年草地;◆:封育15年草地;×:封育23年草地;○封育32年草地。
numbers represents samples. ▼:grazed grassland;△:5-year fenced grassland;◆:15-year fenced grassland;×:23-year fenced grassland;○:32-year fenced grassland.
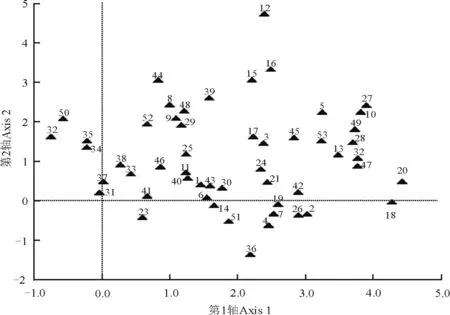
图2 不同封育年限样地地上植被的DCA排序Fig. 2 Different species in different enclosure durations ordination plot
数字代表物种。1野韭;2远志;3赖草;4茅香;5甘菊;6狼毒;7柴胡;8冷蒿;9猪毛蒿;10披碱草;11苦荬菜;12苦苣菜;13白莲蒿;14茵陈蒿;15大针茅;16百里香;17火绒草;18蓬子菜;19委陵菜;20岩败酱;21田葛缕子;22扁穗冰草;23小果黄耆;24裂叶堇菜;25青海苜蓿;26紫花地丁;27细叶沙参;28长柱沙参;29糙隐子草;30干生苔草;31鳞叶龙胆;32甘青针茅;33本氏针茅;34黄毛棘豆;35多叶棘豆;36多茎委陵菜;37星毛委陵菜;38西山委陵菜;39二裂委陵菜;40散穗早熟禾;41硬质早熟禾;42密毛白莲蒿;43大苞点地梅;44直立点地梅;45翼茎风毛菊;46白花枝子花;47白花草木樨;48达乌里秦艽;49北方还阳参;50长叶火绒草;51中华苦荬菜;52阿尔泰狗娃花;53多毛并头黄芩。
Species are represented by numbers: 1Alliumramosum; 2Polygalatenuifolia; 3Leymussecalinus; 4Hierochloeodorata; 5Chrysanthemumlavandulifolium; 6Stellerachamaejasme; 7Bupleurumchinense; 8Artemisiafrigida; 9Artemisiascoparia; 10Elymusdahuricus; 11Ixerisdenticulate; 12Sonchusoleraceus; 13Artemisiasacrorum; 14Artemisiacapillaries; 15Stipagrandis; 16Thymusmongolicus; 17Leontopodiumcalocephalum; 18Galiumverum; 19Potentillachinensis; 20Patriniarupestris; 21Carumburiaticum; 22Agropyroncristatum; 23Astragalustataricus; 24Violadissecta; 25Medicagoarchiducis; 26Violaphilippica; 27Adenophorapaniculata; 28Adenophorastenanthina; 29Cleistogenessquarrosa; 30Carexaridula; 31Gentianasquarrosa; 32Stipaprzewalskyi; 33Stipabungeana; 34Oxytropisochrocephala; 35Oxytropismyriophylla; 36Potentillamulticaulis; 37Potentillaacaulis; 38Potentillasischanensis; 39Potentillabifurca; 40Poasubfastigiate; 41Poasphondylodes; 42Artemisiasacrorum; 43Androsacemaxima; 44Androsaceerecta; 45Saussureajaponica; 46Dracocephalumheterophyllum; 47Melilotusalbus; 48Gentianadahurica; 49Crepiscrocea; 50Leontopodiumlongifolium; 51Ixerischinensis; 52Heteropappusaltaicus; 53Scutellariascordifolia.
不同封育年限样地的地上植被差异较大(图2)。扁穗冰草和长叶火绒草出现在第一排序轴的最左面,即这两种草只出现在放牧地。多叶棘豆、黄毛棘豆、鳞叶龙胆和星毛委陵菜主要出现在演替初期的群落中(封育5年和封育15年样地),排在第一排序轴的偏左面。随着演替的推进,蓬子菜和岩败酱等物种开始出现在封育32年草地群落中。在演替进程的前期和后期,本氏针茅、白莲蒿、二裂委陵菜、干生苔草、火绒草和阿尔泰狗娃花等物种均有出现,说明这几个物种属于泛化种,受群落演替阶段的影响较小。
2.4 不同封育年限草地地上植被演替的聚类分析
树状图基部的相似值是52%,说明不同封育年限草地地上植被的相似性较高(图3)。封育23年和32年草地的地上植被相似性最高,其次为封育5年草地和放牧地之间的相似性。封育15年草地与封育23年和封育32年草地相似性较高,与放牧地和封育5年草地之间的物种组成差异较大。草地地上植被之间相关性的聚类分析结果与DCA排序结果一致。
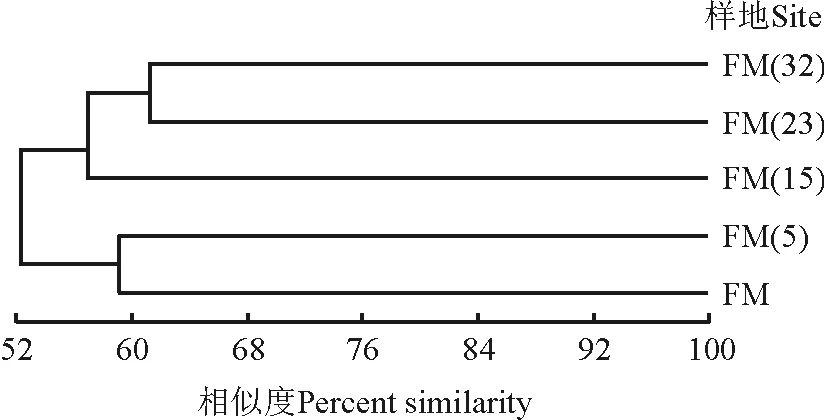
图3 不同草地群落地上植被的UPGMA聚类分析Fig. 3 A UPGMA cluster analysis of vegetation in grasslands with different enclosure durations
3 讨论
由于野外长期监测样地较少及野外选择样地较困难,本研究采用空间分布代替时间演替的方法,研究了不同年限封育对黄土高原典型草原地上植被的影响。虽然该方法有一定的局限性,但许多关于群落演替的研究均采用此方法,且结果准确可靠[24-25]。本研究表明,随着封育年限的延长草地枯落物量和厚度逐渐增长,而群落密度却呈逐渐降低趋势。在松嫩草原,地上枯落物的积累能够显著降低植物总密度,对植物总生物量以及各植物类群生物量则无统一作用规律[26]。导致这一结果可能有两个原因,一是枯落物的积累对草地优势物种的生长具有一定的抑制作用[26];二是枯落物的积累而形成一层厚厚的未分解层,阻碍了牧草种子与土壤的接触,降低了发芽率,影响幼苗生长,导致密度下降[8,16]。在本研究中,随着围封年限的延长,地上总生物量和禾草生物量均呈先增加后降低趋势。在昭苏马场春秋牧场上,地上生物量在封育5年后最大,在封育6年后开始下降[27]。在内蒙古锡林郭勒盟退化羊草草原,群落生物量在封育2年后就达到最大,然后随着封育时间的延长生物量逐渐下降[3]。在内蒙古白音郭勒牧场,地上生物量是在封育6年草地达到最大,在封育26年草地明显下降[28]。但王海瑞[29]研究指出,地上生物量随着封育年限的延长而增加,且在封育27年草地达到峰值。而在高寒草甸,地上生物量随着封育年限的延长呈先显著降低后显著增加,再显著降低趋势,且在第4年生物量最大[30]。可见,封育后草地植被初级生产力变化情况还无统一结论,可能与草地植被类型、封育时间长短和气候条件等因素有关。在该区封育草地,是以多年生禾草为优势种,多年生禾草的生物量占总生物量的60%以上,因此多年生禾草生物量随封育年限的动态变化直接决定着植被初级生产力的变化情况。随着封育年限的增加,植被群落经历了杂草阶段→丛生禾草阶段→本氏针茅群落相对稳定阶段的演替进程。因此,在黄土高原地区,典型草原群落类型可能是解释地上生物量随封育年限动态变化的关键因素。
有关封育年限对草地物种多样性的研究也较多,但不同的研究得出了不同的结论。随封育年限的增加,有研究发现物种多样性一直呈增加趋势[31],有研究指出,物种多样性呈先下降后上升趋势[27],也有研究指出物种多样性呈先增加后降低趋势[32],还有研究表明物种多样性呈先上升后下降再上升的趋势[33-34]。本研究表明,地上植被的物种丰富度和Shannon-Wiener多样性指数均呈先升高后下降的趋势。短期封育可以显著增加物种多样性,长期封育反而不利于物种多样性的保护。这与苗仁辉等[35]和王黎黎等[36]的研究结果一致,他们指出植被多样性指数和均匀度指数随封育年限的增加而下降,长期封育不利于物种多样性的维持。产生研究结果不一致的原因较多,主要包括群落类型、封育年限、地理位置和环境条件等。本研究中,封育15年草地的物种多样性达到峰值。
本研究表明,放牧地和封育5年草地相似性较高,封育23年草地和封育32年草地相似性较高。在放牧地,本氏针茅群落中高大的优良的禾草被家畜采食,而家畜不喜食的低矮的非禾本科草逐渐占据优势,形成了杂类草大量出现的状态。草地封育后,退化草地恢复的动力来自于生态系统的自身演替。草地封育5年后,优良禾草如本氏针茅才开始大量出现,但还未在草地群落中形成优势地位,群落中还大量保存着围封以前的耐牧植物,因此封育5年草地和放牧地的物种相似性较高。围封15年后,草地的物种多样性和生物量都显著提高,是围封年限最佳时期。封育23年草地和封育32年草地是本氏针茅群落稳定发展阶段,但这个时期物种多样性和生物量开始下降。高大的本氏针茅在群落中占有绝对优势地位,一些低矮的不耐阴的物种会从群落中退出。针对草地群落演替,生态学家提出了多种演替机制理论或学说[37-38],但由于群落类型和立地条件的差异,目前还没有形成统一的演替理论。
4 结论
1)随着封育年限的增加,枯落物量和厚度呈递增趋势,群落盖度呈先增加后降低趋势,群落密度呈递减趋势。地上生物量和禾草地上生物量均呈先显著增加后降低的趋势,且均在封育23年草地达到了峰值。地上植被的物种丰富度和Shannon-Wiener多样性指数均呈先增加后降低趋势。综合生物量和物种多样性,围封年限最佳为15年。2)封育23年和32年草地的地上植被相似性最高,其次为封育5年草地和放牧地之间的相似性。随着封育年限的增加,该区草地群落经历了杂草阶段→丛生禾草阶段→本氏针茅群落相对稳定阶段的演替进程。
References:
[1] 程杰,呼天明,程积民.黄土高原半干旱区云雾山封禁草原30年植被恢复对气候变化的响应.生态学报,2010,30(10):2630-2638.
Cheng J,Hu T M,Cheng J M.Response of vegetation restoration to climate change during the past 30 years in enclosed grassland of Yunwu Mountain in semi-arid region of the Loess Plateau.Acta Ecologica Sinica,2010,30(10):2630-2638. (in Chinese)
[2] 闫玉春,唐海萍,辛晓平,王旭.围封对草地的影响研究进展.生态学报,2009,29(9):5039-5046.
Yau Y C,Tang H P,Xin X P,Wang X.Advance in resrarch on the effects of exclosure on grasslands.Acta Ecologica Sinica,2009,29(9):5039-5046.(in Chinese)
[3] 左万庆,王玉辉,王风玉,师广旭.围栏封育措施对退化羊草草原植物群落特征影响研究.草业学报,2009,18(3):12-19.
Zuo W Q,Wang Y H,Wang F Y,Shi G X.Effect of enclosure on the community characterist ofLeymuschinenesisin degenerated steppe.Acta Prataculturae Sinica,2009,18(3):12-19.(in Chinese)
[4] 许中旗,李文华,许晴,闵庆文,王英舜,何旭生.禁牧对锡林郭勒典型草原物种多样性的影响.生态学杂志,2008,27(8):1307-1312.
Xu Z Q,Li W H,Xu Q,Min Q W,Wang Y S,He X S.Effects of grazing-prohibition on species biodiversity of typical steppe in Xilinguole.Chinese Journal of Ecology,2008,27(8):1307-1312.(in Chinese)
[5] Zhao L P,Su J S,Wu G L,François G.Long-term effects of grazing exclusion on aboveground and belowground plant species diversity in a steppe of the Loess Plateau,China.Plant Ecology and Evolution,2011,144(3):313-320.
[6] Wu G L,Liu Z H,Zhang L,Chen J M,Hu T M.Long-term fencing improved soil properties and soil organic carbon storage in an alpine swamp meadow of western China.Plant and Soil,2010,332:331-337.
[7] Cheng J,Wu G L,Zhao L P,Li Y,Cheng J M.Accumulative effects of 20-year fencing on above and belowground biomass of typical steppe communities in arid areas of the Loess Plateau,China.Plant,Soil and Environment,2011,57(1):40-44.
[8] 赵凌平,谭世图,白欣,王占彬,程积民,王清义.封育和放牧对黄土高原典型草原繁殖更新的影响.西北农林科技大学学报(自然科学版),2016,44(9):27-34.
Zhao L P,Tan S T,Bai X,Wang Z B,Cheng J M,Wang Q Y.Effect of grazing and grazing exclusion management on offspring recruitment in a semiarid steppe on the Loess Plateau.Journal of Northwest A & F University (Natuhe Science Edition),2016,44(9):27-34.(in Chinese)
[9] 何念鹏,韩兴国,于贵瑞.长期封育对不同类型草地碳贮量及其固持速率的影响.生态学报,2011,31(15):4270-4276.
He N P,Han X G,Yu G R.Carbon and nitrogen sequestration rate in long-term fenced grasslands in Inner Mongolia,China.Acta Ecologica Sinica,2011,31(15):4270-4276.(in Chinese)
[10] 高凯,朱铁霞,韩国栋.围封年限对内蒙古羊草-针茅典型草原植物功能群及其多样性的影响.草业学报,2013,22(6):39-45.
Gao K,Zhu T X,Han G D.Impact of enclosure duration on plant functional and species diversity in Inner Mongolian grassland.Acta Prataculturae Sinica,2013,22(6):39-45.(in Chinese)
[11] 刘小丹,张克斌,王晓,马福江.围封年限对沙化草地群落结构及物种多样性的影响.水土保持通报,2015,35(3):39-43.
Liu X D,Zhang K B,Wang X,Ma F J.Influence of enclosure ages on community structure and species diversity of sandy grassland.Bulletin of Soil and Water Conservation,2015,35(3):39-43.(in Chinese)
[12] 郝红敏,刘玉,王冬,杨政,武高林.典型草原开垦弃耕后不同年限群落植物多样性和空间结构特征.草地学报,2016,24(4):754-759.
Hao H M,Liu Y,Wang D,Yang Z,Wu G L.Influence of restoration on community plant diversity and structure of typical grasslands.Acta Agrestia Sinica,2016,24(4):754-759.(in Chinese)
[13] 李强.不同恢复措施对松嫩平原退化草地的作用.长春:东北师范大学硕士学位论文,2010.
Li Q.The effect of different restoration measures on degraded grassland in Songnen Plain.Master Thesis.Changchun:Northeast Normal University,2010.(in Chinese)
[14] 刘美珍,蒋高明,于顺利,李永庚,高雷明,牛书丽,姜闯道,彭羽.浑善达克退化沙地恢复演替18年中植物群落动态变化.生态学报,2004,24(8):1187-1193.
Liu M Z,Jiang G M,Yu S L,Li Y G,Gao L M,Niu S L,Jiang C D,Peng Y.Dynamics of plant community traits during an 18-year natural restoration in the degraded sandy grassland of Hunshandak Sandland.Acta Ecologica Sinica,2004,24(8):1187-1193.(in Chinese)
[15] 庞吉林,张克斌,王海星,王黎黎,苏朋飞,王晓.干旱半干旱区人工封育对地表植被组成及多样性的影响:以宁夏盐池为例.东北林业大学学报,2013,41(2):40-43.
Pang J L,Zhang K B,Wang H X,Wang L L,Su P F,Wang X.Effects of artificial enclosure on vegetation composition and plant diversity in arid and semi-arid region:Yanchi County,Ningxia as an example.Journal of Northeast Forestry University,2013,41(2):40-43.(in Chinese)
[16] 张晶晶.封育对宁夏荒漠草原植被的影响研究.银川:宁夏大学硕士学位论文,2012.
Zhang J J.Effects of fencing on vegetation in desert steppe in Ningxia.Master Thesis.Yinchuan:Ningxia University,2012.(in Chinese)
[17] Qiu L P,Wei X R,Zhang X C,Cheng J M.Ecosystem carbon and nitrogen accumulation after grazing exclusion in semiarid grassland.PloS One,2013,8(1):1-7.
[18] 张爱宁,安沙舟,张蕊思,夏小伟,卡斯达尔·努尔旦别克,董磊.不同退化梯度下大尤尔都斯高寒草原植物种群生态位特征.草业科学,2017,34(2):302-309.
Zhang A N,An S Z,Zhang R S,Xia X W,Kasidar Nuerdanbieke,Dong L.Study on niche characteristics of the plant community in the alpine grassland of Big Youerdusi under a degeneration gradient.Pratacultural Science,2017,34(2):302-309.(in Chinese)
[19] 罗久富,周金星,赵文霞,董林水,郑景明.围栏措施对青藏高原高寒草甸群落结构和稳定性的影响.草业科学,2017,34(3):565-574.
Luo J F,Zhou J X,Zhao W X,Dong L S,Zheng J M.Effect of fences on functional groups and stability of the alpine meadow plant community in the Qinghai-Tibet Plateau.Pratacultural Science,2017,34(3):565-574.(in Chinese)
[20] 马培杰,李亚娇,潘多锋,陈本建,李心诚,王德利.牦牛干扰下草原毛虫对小嵩草高寒草甸植物群落特征的影响.草业科学,2017,34(4):698-705.
Ma P J,Li Y J,Pan D F,Chen B J,Li X C,Wang D L.Effect of grassland caterpillar on the characteristics of the vegetation ofKobresiapygmaeaalpine grassland under the interference of the yak grazing.Pratacultural Science,2017,34(4):698-705.(in Chinese)
[21] 马克平,黄建辉,于顺利,陈灵芝.北京东灵山地区植物群落多样性的研究Ⅱ丰富度、均匀度和物种多样性指数.生态学报,1995,15(3):268-277.
Ma K P,Huang J H,Yu S L,Chen L Z.Plant community diversity in Dong Ling Mountain,Beijing,China:Ⅱ.Species richness,evenness and species diversity.Acta Ecologica Sinica,1995,15(3):268-277.(in Chinese)
[22] 王琳,张金屯,上官铁梁,樊龙锁.历山山地草甸的物种多样性及其与土壤理化性质的关系. 应用与环境生物学报,2004,10(1):18-22.
Wang L,Zhang J T,Shangguan T L,Fan L S.Species diversity of mountain meadow of Lishan and the relation with the soil physicochemical properties.Chinese Journal of Applied and Environmental Biology,2004,10(1):18-22.(in Chinese)
[23] 张金屯.数量生态学.2版.北京:科学出版社,2011.
Zhang J T.Quantitative Ecology.2 edition.Beijing:Science Press,2011.(in Chinese)
[24] 江洪,张艳丽,Strittholt J R.干扰与生态系统演替的空间分析.生态学报,2003,23(9):1861-1876.
Jiang H,Zhang Y L,Strittholt J R.Spatial analysis of disturbance and ecosystem succession.Acta Ecologica Sinica,2003,23(9):1861-1876.(in Chinese)
[25] 王本洋,余世孝,王永繁.植被演替过程中种群格局动态的分形分析.植物生态学报,2006,30(6):924-930.
Wang B Y,Yu S X,Wang Y F.Fractal analysis of the dynamics of population patterns during vegetation succession.Journal of Plant Ecology,2006,30(6):924-930.(in Chinese)
[26] 李晓菲.氮沉降与枯落物输入对松嫩草地植物及土壤特征的影响.长春:东北师范大学博士学位论文,2015.
Li X F.Effects of nitrogen deposition and litter input on plant and soil characteristics of songnen grassland.PhD Thesis.Changchun:Northeast Normal University,2015.(in Chinese)
[27] 李军保,朱进忠,吐尔逊娜依·热依木江,刘洪来.围栏封育对昭苏马场春秋草场植被恢复的影响.草原与草坪,2007,125(6): 45-48.
Li J B,Zhu J Z,Tursunnayi·Reyimujiang,Liu H L.Effects of enclosing spring-autumn grassland on vegetative restoration at Zhaosu horse ranch.Grassland and Turf,2007,125(6):45-48.(in Chinese)
[28] 闫玉春,唐海萍,常瑞英,刘亮.典型草原群落不同围封时间下植被、土壤差异研究.干旱区资源与环境,2008,22(2):145-151.
Yan Y C,Tang H P,Chang R Y,Liu L.Study on the difference of vegetation and soil in typical steppe communities under different fenced time.Journal of Arid Land Resources and Environment,2008,22(2):145-151.(in Chinese)
[29] 王海瑞.典型草原退化群落不同恢复演替阶段土壤物理性质及渗水性能的研究.呼和浩特:内蒙古大学硕士学位论文,2011.
Wang H R.The study of soil physical properties and infiltration capacity of typical grassland in different recovery stages.Master Thesis.Huhhot:Inner Mongolia University,2011.(in Chinese)
[30] 苗福泓,郭雅婧,缪鹏飞,郭正刚,沈禹颖.青藏高原东北边缘地区高寒草甸群落特征对封育的响应.草业学报,2012,21(3):11-16.
Miao F H,Guo Y J,Miao P F,Guo Z G,Shen Y Y.Influence of enclosure on community characteristics of alpine meadow in the northeastern edge region of the Qinghai-Tibetan Plateau.Acta Prataculturae Sinica,2012,21(3):11-16.(in Chinese)
[31] 吴建波,包眹影,李洁,赵念席,高玉葆.不同围封年限对典型草原群落及大针茅种群特征的影响.草地学报,2010,18(4):490-495.
Wu J B,Bao X Y,Li J,Zhao N X,Gao Y B.Influence of fencing duration on community and population ofStipagrandisin a typical steppe.Acta Agrectir Sinica,2010,18(4):490-495.(in Chinese)
[32] 赵菲,谢应忠,马红彬,沈艳.封育对典型草原植物群落物种多样性及土壤有机质的影响.草业科学,2011,28(6):887-891.
Zhao F,Xie Y Z,Ma H B,Shen Y.Effects of enclosure on species diversity and soil organic matter of typical steppe.Pratacultural Science,2011,28(6):887-891.(in Chinese)
[33] 单贵莲,徐柱,宁发,马玉宝,李临杭.围封年限对典型草原群落结构及物种多样性的影响.草业学报,2008,17(16):1-8.
Shan G L,Xu Z,Ning F,Ma Y B,Li L H.Influence of exclosure year on community structure and species diversity on a typical steppe.Acta Prataculturae Sinica,2008,17(16):1-8.(in Chinese)
[34] 贾晓妮.云雾山25年本氏针茅草地群落特征及演替的研究.北京:中国科学院研究生院硕士学位论文,2008.
Jia X N.Vegetation characteristics and succession of 25-yearStipabungeanacommunity in Yunwu mountain.Master Thesis.Beijing:University of Chinese Academy of Sciences,2008.(in Chinese)
[35] 苗仁辉,蒋德明,王永翠.科尔沁沙质草地封育过程中的植被变化及其机制.干旱区研究,2013,30(2):264-270.
Miao R H,Jiang D M,Wang Y C.Change and mechanism of vegetation in the fenced sandy grassland in Horqin Sandy Land.Arid Zone Research,2013,30(2):264-270.(in Chinese)
[36] 王黎黎,张克斌,程中秋,常进,张静雯,苏鹏飞,王晓.围栏封育对半干旱区植物群落稳定性的影响.甘肃农业大学学报,2011,48(5):86-90.
Wang L L,Zhang K B,Cheng Z Q,Chang J,Zhang J W,Su P F,Wang X.Effects of fencing on stability of plant community in semi-arid area.Journal of Gansu Agricultural University,2011,48(5):86-90.(in Chinese)
[37] Clements F E.Plant Succession:An Analysis of the Development of Vegetation.Washington:Carnegie Institute Scientific Publication,1916.
[38] Tilman D.Mechanisms of Plant Competition for Nutrients:The Elements of a Predictive Theory of Competition.San Diego:Academic Press,1990.
