南极红酵母AN5的发酵及对海参体成分的影响
2018-03-03王晓飞史翠娟阚光锋李亮文华周婷
王晓飞 史翠娟 阚光锋 李亮 文华 周婷
(1哈尔滨工业大学(威海)功能食品研究室,山东 威海 264209;2山东大学(威海)海洋学院,山东 威海 264209)
0 引言
红酵母是海洋微生物的优势种群,广泛存在于世界各个海域、南北极及深海等领域[1-3]。海洋是一个高压(深海)、低营养、低光照,以及温度和盐度变化巨大的独特环境,从而使其中的酵母细胞内代谢和产物有异于陆地环境生存的酵母。海洋红酵母细胞含有丰富的蛋白质、类胡萝卜素、多糖、虾青素、不饱和脂肪酸、必需氨基酸、消化酶类、维生素等活性代谢产物,具有较高的营养价值和活性作用,不仅用以生产类胡萝卜素、单细胞蛋白和生物柴油,还可用于微生态制剂,净化水质,在医药、食品、化工、农业和环保等领域有着广泛的应用[4-5]。
海洋红酵母大小约为 4—6 μm,营养物质丰富,适口性好,不易沉淀,有较好的耐盐性,同时还具有培养周期短、适应能力强、成本低等优点,非常适宜作为海产动物幼体的开口饵料和幼体时期的补充性饵料,能显著提高水产动物幼苗存活率,提高饲料利用率,增强动物体免疫力,净化水体,是生态养殖的优良添加剂[6-7]。海洋红酵母作为养殖微生态制剂在水产养殖业中已经有广泛的应用,并推广到了广西、海南、江苏、浙江、山东等十几个省市[8]。
与一般海洋环境相比,南极具有低温、高盐、低光照、寡营养、强辐射等极端环境特点,孕育了包括酵母在内的丰富的低温微生物资源,能够产生独特的代谢物质[9]。因此,本研究以极地红酵母Rhodotorula mucilaginosaAN5为实验对象,研究其发酵培养的最佳生长条件,初步探讨作为养殖微生态制剂在水产养殖业中应用的可能性。
1 材料与方法
1.1 实验菌株
海洋极地酵母Rhodotorula mucilaginosaAN5是从第22次南极科学考察采集的海冰样品中分离所得。
1.2 酵母培养基及培养
YPD培养基: 葡萄糖2.0 g,蛋白胨2.0 g,酵母粉1.0 g,pH=6.0,海水100 mL,0.1 MPa下灭菌30 min。将活化4 d的酵母接种于液体YPD培养基中,120 r·min–1,20℃摇床培养。
1.3 酵母密度测定
采用显微计数法[10]。
1.4 海参的养殖
实验用海参为仿刺参幼苗,购自山东威海,长约0.5—0.8 cm。将幼参随机分到6个玻璃缸中人工养殖(3个为对照组,3个为实验组),每天定时投喂等量人工配合饲料,实验组每天另投喂0.3 g红酵母,每2天换水1/2,2个月后,收集海参,吸干表面水分,−20℃保存。
1.5 成分测定方法
水分含量测定采用国标GB5009.3—2010直接干燥法[11]; 蛋白质含量测定根据国标 GB5009.5—2010凯氏定氮法[12]; 粗脂肪含量测定采用索氏抽提法[13]; 多糖含量测定采用亚甲基蓝比色法[14]。
1.6 数据分析
试验数据为 3次生物学重复平均所得,以平均值±标准误差表示,用软件 SPSS 15.0中的one- way ANOVA进行方差分析和差异显著性比较,P<0.05为差异显著,P<0.01为极显著差异。
2 结果与分析
2.1 培养条件对酵母发酵生长的影响
2.1.1 碳源对酵母生长的影响
分别以2%的玉米淀粉、蔗糖、糖蜜、玉米面、豆面、小米面代替YPD培养基中的葡萄糖,其他成分和培养条件不变,4 d后显微计数法测定菌体密度(图1)。6种碳源物质中,以糖蜜为碳源进行发酵培养时,酵母细胞密度达到最大值 1.19×107cells·mL–1,其次为小米面 0.91×107cells·mL–1,玉米淀粉、玉米面、豆面和蔗糖都约为 0.65×107cells·mL–1,显著小于前2种。糖蜜含有海洋红酵母生长所必需的营养成分、维生素及微量元素等,发酵时明显优于其他碳源[15]。同时糖蜜是大宗廉价原料,价格便宜,综合考虑发酵菌量与生产成本,糖蜜为酵母菌培养的最佳碳源。分别以1.0%、2.0%、3.0%和4.0%浓度的糖蜜培养红酵母,酵母细胞密度见图2。结果发现2%的糖蜜发酵液中酵母细胞密度最大,为1.46×107cells·mL–1,明显高于其他浓度,由此确定2%糖蜜为酵母培养的最佳碳源。

图1 不同碳源对南极红酵母AN5发酵的影响Fig.1.Effect of different carbon sources on Antarctic yeast AN5 growth

图2 糖蜜浓度对南极酵母AN5发酵的影响Fig.2.Effect of different molasses concentration on Antarctic yeast AN5 growth
2.1.2 氮源对酵母发酵的影响
以上述优化的 2%的糖蜜为碳源,分别以液体玉米浆、固体玉米浆、糖蜜、玉米面、酵母粉、黄豆饼粉、蛋白胨、大豆蛋白胨、硫酸铵、硝酸钾为氮源,发酵培养 4 d后,测定酵母细胞密度(图3)。在10种氮源物质中,对AN5生长量的作用大小为酵母粉>大豆蛋白胨>蛋白胨>玉米面>固体玉米浆>硝酸钾>液体玉米浆>糖蜜>黄豆饼粉>硫酸铵。在8种有机氮源中,以酵母粉作为氮源进行发酵培养时,酵母细胞密度达最大值1.3×107cells·mL–1,因此选取酵母粉作为发酵培养的最佳氮源,与杨莺莺等[16]的研究结果一致。两种无机氮源中,硝态氮的作用效果远高于铵态氮,杨莺莺等[16]对酵母RH1的发酵研究表明,硫酸铵和氯化铵作氮源时生长缓慢,与本研究结果一致。筛选出最佳氮源后,分别以0.25%、0.50%、0.75%和1.0%酵母粉浓度培养酵母(图4)。有图可知,以浓度为 0.5%的酵母粉进行发酵培养时,发酵液中酵母细胞密度可以达到2.03×107cells·mL–1,随酵母粉浓度增加酵母细胞密度基本不变。考虑到发酵的密度和成本,选择0.5%酵母粉为酵母培养的最佳氮源。

图3 不同氮源对南极酵母AN5发酵的影响Fig.3.Effect of different nitrogen sources on Antarctic yeast AN5 growth

图4 酵母粉浓度对南极酵母AN5发酵的影响Fig.4.Effect of different yeast powder concentration on Antarctic yeast AN5 growth
2.1.3 初始pH对酵母生长的影响
以上述优化的碳氮源为培养基,调节初始pH值,使其 pH值分别为 2.0、3.0、4.0、5.0、6.0、7.0、8.0,培养 4 d后测定菌体的密度(图5)。当pH=3.0时,发酵液中酵母细胞密度最大,为1.19×107cells·mL–1; 当pH=2.0 时,菌体密度迅速下降,达到最低密度值 0.32×107cells·mL–1; 当pH>3时,随 pH的增大,细胞密度变小,但下降缓慢,pH=8.0 时密度仍为 0.54×107cells·mL–1。由此可知,海洋酵母细胞培养的最适初始 pH=3.0,显著低于常温酵母最适pH为4.0—5.0的报道[5,16]。
2.1.4 温度对酵母生长的影响
以上述优化的碳氮源为培养基,分别在10℃、15℃、20℃、25℃和30℃下培养酵母菌,4 d后酵母细胞密度见图6。由图可知,当温度从10℃升高到20℃时,菌体密度达到最大值 0.96×107cells·mL–1。其后随温度的继续升高,酵母密度显著下降。因此,AN5菌株生长的最适温度为20℃,发酵时应控制在这一温度。酵母菌的最适生长温度因种属或菌株的来源不同而有所差别,杨莺莺等[16]和童应凯等[17]发现海洋红酵母Rhodotorulasp.的最佳培养温度为28℃,郭馨泽等[18]实验证明海洋红酵母003菌株的最佳生长温度为22℃。实验中所用酵母为海洋极地酵母,属适冷酵母种类,在低温环境下仍能保持活力,一般低于常温酵母的最适生长温度25—28℃[5]。

图5 初始pH对南极酵母AN5生长的影响Fig.5.Effect of different initial pH on Antarctic yeast AN5 growth

图6 温度对南极酵母AN5生长的影响Fig.6.Effect of different temperature on Antarctic yeast AN5 growth
2.1.5 装液量对酵母生长的影响
在250 mL的三角瓶中加入培养基和酵母种子液,使其终体积分别为30、50、70、90、110、130、150 mL,4 d后对菌液密度进行测定(图7)。当装液量为 50 mL时,酵母细胞密度达最大值0.99×107cells·mL–1; 装液量为 30 mL和70 mL时,细胞密度显著降低; 当装液量大于70 mL时,菌体密度无显著变化,维持在 0.58×107cells·mL–1。装液量的不同对酵母菌生长的影响体现在两个方面:(1)装液量越大,营养物质含量越多,为酵母菌的生长提供物质保证;(2)装液量越大意味着剩余空间越小,三角瓶内空气越少,随着时间增加,瓶内氧气消耗,废气增多,对酵母菌的生长起到抑制作用。本实验中,50 mL/250 mL的装液量为酵母生长的最佳体积,与郭馨泽等[18]的研究结果一致。
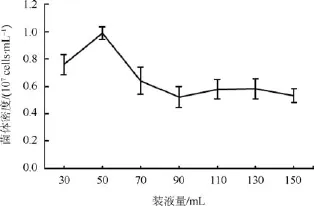
图7 装液量对南极酵母AN5发酵的影响Fig.7.Effect of different liquid volume on Antarctic yeast AN5 growth
2.1.6 转速对酵母生长的影响
以优化的碳氮源为培养基,装液量为 50 mL/250 mL,设置摇床的转速分别为80、100、120、140、160 和 180 r·min–1,4 d 后对菌液密度进行测定(图8)。由图8可以看出,随着摇床转速的增加,酵母发酵密度呈上升趋势,由 80 r·min–1时的0.93×107cells·mL–1到 140 r·min–1时的1.59×107cells·mL–1,增加了 71%。此后,随转速的升高,菌体密度几乎无变化,故本实验的最佳摇瓶转速为140 r·min–1。摇瓶培养的转速和装液量对菌体发酵密度的影响因素基本一致,影响培养基中氧气的含量,转速越高,培养基中氧气的含量增加,利于酵母菌的生长繁殖,但当氧气含量达到一定限度后,酵母菌的密度就不再增加。
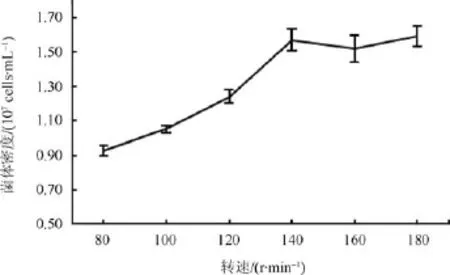
图8 转速对南极酵母AN5发酵的影响Fig.8.Effect of different rotation rate on Antarctic yeast AN5 growth
经上述条件优化,得到了极地红酵母AN5的最佳发酵条件,此时酵母密度达到 5.81×107cells·mL–1,与周鲜娇等[19]的研究结果类似,但低于其他常温红酵母[16,18]。
2.2 极地红酵母主要成分含量的测定
以优化后的培养条件大量发酵红酵母 AN5,所得菌体进行水分和主要营养成分的测定。经测定,极地红酵母AN5的平均含水量为69.15%。主要营养成分测定结果见表1,从表中可以看出,干酵母中粗蛋白含量约为42.00%,与常温红酵母中蛋白质含量相似,符合水产养殖中养殖动物对生物饵料的蛋白需求[20]。粗脂肪含量仅为0.39%,显著低于蔡诗庆等[21]测定的4.67%—6.35%。总糖是生物体能量的主要来源,酵母菌AN5的总糖含量为 32.08%,高于常温酵母[14,22],充足的糖类可被用以生物体生长能源的消耗,在水产养殖中发挥重要作用[23]。

表1 红酵母Rhodotorula sp.的主要营养成分Table 1.Main nutrient composition in several red yeasts Rhodotorula sp.
2.3 极地红酵母对幼参水分和主要营养成分的影响
以每天投喂0.3 g鲜酵母喂养海参幼苗,2个月后,测定海参体内的水分和主要营养成分含量(图9)。正常养殖的海参,含水量为92.02%,投喂红酵母的海参,含水量为 91.83%,水分含量没有显著差异; 投喂红酵母后,蛋白质干重由 35.84%增加到 43.19%,含量显著提高; 脂肪含量由7.77%变为7.95%,无明显变化; 但添加红酵母的海参中多糖的含量为 6.75%干重,较对照的5.46%增加了23.63%。

图9 南极酵母AN5作为微生态制剂对海参主要成分的影响.*表示对照组和实验组之间差异显著(P<0.05)Fig.9.Effect of Antarctic yeast AN5 as microecologics on main components of juvenile sea cucumber
3 讨论
3.1 酵母菌的发酵生产
酵母菌的大规模发酵生产是红酵母应用的前提和基础。科学家研究发现,不同的培养条件和发酵工艺会影响海洋红酵母的生物量,通过调整培养基中碳源、氮源、无机盐、生长因子等物质的种类和添加量,改变培养过程中的pH值、接种量、装液量、通气量等条件,可显著提高酵母菌的产量[14,18]。本实验所用碳氮源均为工业发酵原料,廉价易得,经过发酵工艺的优化,酵母细胞密度增加了6.21倍,达到了常温酵母发酵的生物量,为酵母细胞的大规模发酵生产奠定了基础。但本实验仅对单一的碳氮源进行了筛选,已有研究表明多种营养物质的组合更有利于酵母的生长[18-19],这是下一步研究的方向。另外,该研究只以生物量为指标进行了发酵优化,发酵条件对主要功能活性因子的影响有待进一步研究,以提高酵母菌的营养价值。值得一提的是,该红酵母的最佳发酵温度为 20℃,最适初始 pH为3.0。低温和低pH值培养可抑制杂菌的生长、减少污染。
3.2 红酵母作为饲料添加剂的应用潜力
红酵母是一种在海洋中广泛存在的单细胞微生物,复杂的生存环境造就了体内独特的代谢产物和活性物质[5],且酵母本身无毒副作用,安全性高[24],在果实保鲜[25-26]、畜禽养殖[27-28]等方面有着广泛的应用。
海洋红酵母及其发酵产物富含蛋白质、糖类、虾青素、维生素、矿物质等,含有较多的必需氨基酸,营养全面,是集营养与保健为一体的养殖微生态制剂,这些特殊营养成分弥补了常规饵料(或添加剂)的营养缺陷,可以提高水产动物抵抗力,减少抗生素的使用,提高水产品质量[29]。海洋红酵母Rhodotorulasp.作为微生物饵料在水产苗种培育领域的应用始于20世纪90年代,科学家们进行了大量的研究。陈昌福等[30]和Yang等[31]研究发现,饵料中添加海洋红酵母能够提高对虾的成活率和生长速率,使血清中抗氧化酶系统活性增强,增加对环境因子的耐受力。孙建男等[5]综述文献指出,海洋红酵母对养殖动物的生长免疫有明显的促进作用,可能是由于酵母生长中产生的消化酶类、维生素、生长因子等,促进了动物的消化吸收,提高了饲料的利用率。但张瑞玲等[32]在白鱼粉为主的大菱鲆饲料中添加海洋红酵母,则不能提高大菱鲆的摄食和生长性能。
本研究中,极地红酵母蛋白质和多糖的含量较高,作为微生态制剂应用于幼参的养殖中,能显著提高海参中蛋白和多糖的含量。海参饲喂酵母后,可在幼参体内存活定殖[33-34],体内超氧化物歧化酶、多酚氧化酶、过氧化氢酶、酸性磷酸酶、碱性磷酸酶、溶菌酶、淀粉酶、纤维素酶、褐藻酸酶、一氧化氮合酶、胰酶、脂肪酶等酶类活性显著升高[34-39],促进了饵料的消化和吸收,使海参体内的蛋白质和多糖等主要营养成分物质含量增加[35],对生长具有较明显的促进作用[2,40],免疫反应增强,抗病力和存活率提高[33,36-38]。同时,红酵母作为营养型生物饵料直接为稚参提供营养物质,可提高海参幼体变态率和成活率[41]。除此之外,酵母具有较高的硝化能力,可有效去除海参养殖污水中的亚硝酸盐,改良养殖水质[42]。这些研究为酵母在海参养殖中的应用奠定了理论基础。
南极红酵母易于大规模发酵生产,且营养价值丰富,具有作为水产饵料生物的潜力,是较好的饲料原料,可用作鱼粉等蛋白源的替代物,具有良好的市场前景和开发潜力。
1 Connell L,Barrett A,Templeton A,et al.Fungal diversity associated with an active deep sea volcano: Vailulu'u seamount,Samoa[J].Geomicrobiology Journal,2009,26(8): 597—605.
2 Zhang X,Hua M X,Song C L,et al.Occurrence and diversity of marine yeasts in Antarctica environments[J].Journal of Ocean University of China,2012,11(1): 70—74.
3 Singh P,Singh S M,Tsuji M,et al.Rhodotorula svalbardensissp.nov.,a novel yeast species isolated from cryoconite holes of Ny-Ålesund,Arctic[J].Cryobiology,2014,68(1): 122—128.
4 刘雨,陈霞,宋琦,等.海洋红酵母的作用机理及其在畜禽水产养殖中的应用[J].饲料博览,2014,(4): 42—44.
5 孙建男,谢为天,刘颖,等.海洋红酵母的研究进展[J].安徽农业科学,2015,43(4): 84—88.
6 刘铁钢,赵文,刘钢,等.5种微生态制剂对刺参幼参的生态安全性[J].大连海洋大学学报,2012,27(2): 129—136.
7 王芸,郑宗琳.微生态制剂在水产养殖中的应用研究进展[J].饲料与畜牧,2013,(2): 18—24.
8 尹安伟,路怀灯.海洋红酵母作为水产饲料添加剂的开发应用研究[J].科学养鱼,2010,(5): 64—65.
9 Thomas D N,Dieckmann G S.Antarctic sea ice-a habitat for extremophiles[J].Science,2002,295(5555): 641—644.
10 沈萍,范秀荣,李广武.微生物学实验[M].3版.北京: 高等教育出版社,1996: 90—92.
11 中华人民共和国卫生部.GB 5009.3—2010 食品安全国家标准 食品中水分的测定[S].北京: 中国标准出版社,2010.
12 中华人民共和国卫生部.GB 5009.5—2010 食品安全国家标准 食品中蛋白质的测定[S].北京: 中国标准出版社,2010.
13 玉晏飞,薛飞燕.索氏提取法测定黏红酵母含油量的改进[J].中国粮油学报,2013,28(2): 110—112.
14 杨世平,吴灶和,简纪常.一株海洋红酵母(Rhodosporidium paludigenum)的营养组分分析[J].饲料工业,2011,32(10): 52—54.
15 何海燕,覃拥灵,李楠,等.甘蔗糖蜜培养高铁营养酵母发酵条件优化的研究[J].食品工业科技,2007,28(8): 105—108.
16 杨莺莺,陈永青,杨铿,等.海洋红酵母RH1菌株发酵培养条件的研究[J].南方水产科学,2012,8(1): 67—74.
17 童应凯,韦东胜,刘婷,等.海洋红酵母培养条件的研究[J].食品研究与开发,2006,27(8): 75—79.
18 郭馨泽,刘彤,黄忠刚,等.海洋红酵母的发酵条件[J].大连工业大学学报,2015,34(1): 1—5.
19 周鲜娇,田丽,梁如璞,等.一株海洋红酵母的鉴定及其培养基的优化[J].水产科学,2012,31(2): 107—110.
20 Shiau S Y.Nutrient requirements of penaeid shrimps[J].Aquaculture,1998,164(1—4): 77—93.
21 蔡诗庆,胡超群,任春华.三株海洋酵母的生化营养成分分析[J].热带海洋学报,2009,28(2): 62—65.
22 杨铿,杨莺莺,李卓佳,等.胶红酵母(Rhodotorula mucilaginosa)营养成分分析[J].广东农业科学,2014,41(5): 146—149,154.
23 Sajeevan T P,Philip R,Singh I S B.Dose/frequency: a critical factor in the administration of glucan as immunostimulant to Indian white shrimpFenneropenaeus indicus[J].Aquaculture,2009,287(3—4): 248—252.
24 徐占利,张红印,黄星奕.胶红酵母的急性毒性及遗传毒性研究[J].食品工业科技,2010,31(12): 303—305.
25 束兆林,杨红福,陈红州,等.胶红酵母(Rhodotorula mucilaginosa)对梨果采后青霉病、灰霉病的控制效果[J].江苏农业科学,2015,43(8): 110—112.
26 束兆林,张红印,杨红福,等.胶红酵母(Rhodotorula mucilaginosa)对常温贮藏梨果保鲜效果的研究[J].保鲜与加工,2015,15(6):47—50.
27 孙锡凤,蒋万春.红酵母在动物生产中的应用研究进展[J].饲料研究,2014,(15): 1—5,14.
28 蒋万春,孙锡风,杨俊琦,等.海洋红酵母对肉鸡生产性能、屠体性能及小肠形态结构的影响[J].中国饲料,2015,(10): 15—19.
29 赵述淼,韩继宏.浅谈酵母在水产养殖上的应用[J].中国水产,2003,(7): 68—69.
30 陈昌福,姚鹃,肖雪生,等.海洋1号红酵母饲养日本对虾幼虾的效果[J].淡水渔业,2004,34(6): 55—57.
31 Yang S P,Wu Z H,Jian J C,et al.Effect of marine red yeastRhodosporidium paludigenumon growth and antioxidant competence ofLitopenaeus vannamei[J].Aquaculture,2010,309(1—4): 62—65.
32 张瑞玲,牛思佳,杨振才,等.饲料中海洋红酵母不同添加量对大菱鲆摄食生长的影响[J].河北师范大学学报(自然科学版),2013,37(6): 618—621.
33 Yang Z P,Sun J M,Xu Z.Beneficial effects ofRhodotorulasp.C11 on growth and disease resistance of juvenile Japanese spiky sea cucumberApostichopus japonicus[J].Journal of Aquatic Animal Health,2015,27(2): 71—76.
34 Yang Z P,Sun J M,Xu Z,et al.Beneficial effects ofMetschnikowiasp.C14 on growth and intestinal digestive enzymes of juvenile sea cucumberApostichopus japonicus[J].Animal Feed Science and Technology,2014,197: 142—147.
35 王上.酵母胞外多糖制备及饲喂海参试验[D].大连: 大连工业大学,2016.
36 Wang J H,Zhao L Q,Liu J F,et al.Effect of potential probioticRhodotorula benthicaD30 on the growth performance,digestive enzyme activity and immunity in juvenile sea cucumberApostichopus japonicus[J].Fish & Shellfish Immunology,2015,43(2): 330—336.
37 Ma Y X,Liu Z M,Yang Z P,et al.Effects of dietary live yeastHanseniaspora opuntiaeC21 on the immune and disease resistance againstVibrio splendidusinfection in juvenile sea cucumberApostichopus japonicus[J].Fish & Shellfish Immunology,2013,34(1): 66—73.
38 Liu Z M,Ma Y X,Yang Z P,et al.Immune responses and disease resistance of the juvenile sea cucumberApostichopus japonicusinduced byMetschnikowiasp.C14[J].Aquaculture,2012,368—369: 10—18.
39 宋盛亮,李静,刘佳亮,等.海洋红酵母对仿刺参浮游期、稚参期、幼参期生长和消化的影响[J].齐鲁渔业,2016,(3): 4—7.
40 杨志平,徐哲,周倩,等.饵料中添加海洋红酵母(Rhodotorulasp.)C11对幼参消化酶及免疫反应的影响[J].渔业科学进展,2015,36(6): 107—112.
41 薛德林,胡江春,王国安,等.海洋胶红酵母、光合细菌、生化黄腐酸在海参育苗和养成中的应用效果[J].腐植酸,2009,(4): 19—24.
42 孙蕾,李成,马树瑞,等.一株红酵母的筛选、鉴定及硝化作用[J].大连工业大学学报,2016,35(2): 88—91.
