大理地区碧伟蜓稚虫结构及行为观察
2018-02-02刘秉兰杨国辉张吉龙张关令
刘秉兰,杨国辉,张吉龙,张关令
(大理大学农学与生物科学学院,云南大理 671003)
碧伟蜓(AnaxparthenopejuliusBrauer,1865)属于蜻蜓目(odonata)蜓科(Aeshnidae)伟蜓属(Anax),其头部及身体均为绿色,俗称“绿大头”,在我国广泛分布[1]。碧伟蜓在云南大理地区一年发生一代,即每年5—7月成虫交配、产卵,以稚虫过冬,次年5—8月羽化。
云南的少数民族有食用蜻蜓稚虫的习惯。在大理,人们把碧伟蜓稚虫称作“水蜻蜓”,“油炸水蜻蜓”是白族人民的传统名菜。研究表明蜻蜓目昆虫稚虫含有丰富的蛋白质、脂肪、氨基酸、微量元素等营养成分,是一种有较高开发利用价值的天然昆虫资源[2-4]。
国外对蜻蜓稚虫的研究较为深入,涉及分类、分布格局、行为、生理、环境评价等方面[5-10],关于稚虫形态结构的研究,卓有成就的是英国学者Tiliyard和Cantab,其发表的专著《The Biology of Dragonflies》详细介绍了古北区部分蜻蜓稚虫的外部形态及生物学特性,并探讨了稚虫的生理结构[11],而我国蜻蜓目稚虫方面的研究相对较薄弱,分类方面仅Needham[12]、赵修复[13]、江尧桦[14]、Zhang等[15]少数几位学者对部分类群进行过研究,关于蜻蜓稚虫内部形态结构及行为的寥寥无几[16]。目前,对碧伟蜓AnaxparthenopejuliusBrauer的研究主要集中在地理分布、成虫的行为及稚虫的营养价值,对稚虫内部形态结构及行为的研究未见报道[1,3,4,17]。
对大理地区的碧伟蜓稚虫进行人工饲养,结合野外观察记录,研究其行为习性,并对其形态及内部结构进行研究,为碧伟蜓规模化人工养殖和开展其他研究奠定基础。
1 材料与方法
2015年10月和2016年1月在洱海边水质清澈、水草丰茂的地方,利用孔径为80 mm的水网,在水草下及湖滨区底泥捕捞收集碧伟蜓稚虫,带回实验室进行人工饲养。
将稚虫放入实验室的水族箱中,模拟野外环境,室温19~22 ℃,水温11~13 ℃,鱼缸底部铺设适量的沙石,用石头搭建了几个能让蜻蜓稚虫躲避,蛰伏的洞穴,再放入采自洱海的水生植物和适量水藻,每3 d投喂一次黄粉虫或者水蚯蚓,并在投食3 h后更换培养水,观察记录其捕食、运动和防御等行为,对部分被残食的稚虫用60%酒精保存,进行内部解剖、生物体视成像系统(Nikon-SM21000)记录。
2 结果与分析
2.1行为观察野外碧伟蜓稚虫生活在水质清澈、水草丰茂的水域中,夏季喜在水草中活动,冬季则潜伏在水底泥沙或残枝败叶下。实验室饲养中除白天进食活动外,一般潜伏在水底泥沙或挺水植物的基部。
2.1.1捕食行为。碧伟蜓稚虫口器结构特殊,下颏非常发达,是一个长且可折叠的结构,平时折于腹面,在捕食时快速伸出,并用前方的颏钩夹紧猎物,收回后放入口中。其捕食方式有2种,一种是主动追击,即主动寻找食物,然后追逐、捕获;另一种是偷袭,即潜伏水底等待猎物靠近时,立刻弹出长长的下颏,捕获猎物。在食物短缺时稚虫会彼此相互残食。
2.1.2运动和防御行为。碧伟蜓稚虫在水中除了依靠3对足划水和行走外,还通过胸部和腹部气孔喷出的气流推动身体向前运动,这与蜓科物种相同[18]。
在自然环境中碧伟蜓稚虫既是捕食者,也是被捕食的对象。在研究中观察到其具有明显的防御行为。遇到捕食者会主动逃避,也会用腹部的侧刺、附器及下颏进行搏击。
2.1.3羽化。羽化前稚虫的胸部肌肉收缩,复眼变大,腹部变软,体色由棕色或绿色变成墨绿色,然后缓慢爬出水面,停栖在挺水植物上,早晨9:00左右开始羽化,首先从前胸背中部裂开一个口,胸部先从几丁质的外壳露出,然后头部、六足、腹部依次离开稚虫的外壳,最后翅膀展开,整个过程持续近0.5 h,刚羽化的碧伟蜓身体较柔弱,不能飞行。
2.2外部形态碧伟蜓稚虫一生经历6~8次蜕皮,研究用标本为4~5龄稚虫,体长在50~55 mm,体型较粗壮,体色为褐色、暗褐色或绿色,常因栖息地环境变化有差异。
稚虫体分为头、胸、腹三部分(图1)。头呈三角形,能自由活动,有触角1对,细小,刚毛状;复眼1对,较发达,单眼3个;咀嚼式口器,下颏极度延长,并分3节,尖端有一对可活动的颏钩。
胸部:前胸较小,中胸和后胸愈合,较前胸发达。前胸背板和中胸背板间有气门1对,足3对分别着生在前胸、中胸及后胸的腹面,前后翅芽位于中胸及后胸的背方。
腹部10节,每节均由1个背片、1个腹片及2个体侧片组成,每个体节的体侧片上有气孔1对,两侧体壁有侧刺。尾须较小,不分节,肛附器呈长三角形,长约8 mm。雄性在第9腹节腹片上有1个黑斑,雌性有一个长条状的生殖板,可以来辨别稚虫性别。

图1 碧伟蜓稚虫背面Fig.1 Back of larvae of Anax parthenope julius
2.3内部解剖取60%酒精保存的碧伟蜓稚虫进行解剖观察,先沿腹部2个体侧片自后向前逐步剪去背板,再沿头胸两侧剪开,置于泡沫板上,用大头针固定,剔除遮挡观察的肌肉,移至生物体视成像系统(Nikon-SM21000)下观察。
2.3.1消化系统。消化道自头至尾可分为前肠、中肠、后肠三段。前肠从口开始,经由咽、食道、嗉囊3个器官,终止于前胃,向后伸入中肠前端,以贲门瓣与中肠为界(图2)。口腔的前端开口在上唇和下颏的基部之间,开口呈喇叭形的一小段通道。咽部为前肠的最前端部分,碧伟蜓是咀嚼式口器的昆虫,咽仅是食物的通道。食道位于咽后方的一段狭长管道,是食物的通道。嗉囊是食道后端的膨大部分,其作用是暂时贮藏食物。前胃(砂囊)是前肠的最后一段,是消化道最特化的部位。其形状是一个较小的球形体,作用是用来磨碎食物。
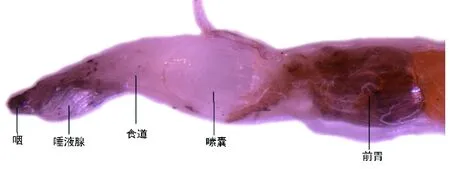
图2 碧伟蜓稚虫前肠Fig.2 Foregut of larvae of Anax parthenope julius
中肠(图3):位于消化道的中部,是一条前后相似的管道,圆筒形,前端与前胃相连,后端与后肠相连,是稚虫消化系统消化吸收的主要部位。

图3 碧伟蜓稚虫的消化和排泄系统Fig.3 Digestion and excretory system of larvae of Anax parthenope julius
后肠(图3):碧伟蜓稚虫后肠包括回肠、结肠、直肠三段。回肠前粗后细,前端内面具幽门瓣;结肠较短,连接回肠和直肠,细而弯曲;直肠在后肠后端,较为粗大,形状呈长椭圆形,末端的开口是稚虫肛门。
消化腺有唾腺,呈葡萄状颗粒,开口在舌下基部,能分泌唾液帮助消化。
2.3.2排泄系统。碧伟蜓稚虫的排泄器官是位于中肠与后肠交界处的2根马氏管,长而粗壮,端段和基端的内容物相似,均为清水状,基部连通直肠。
2.3.3循环系统。碧伟蜓稚虫的循环系统是闭合式循环,大部分器官的供血靠背血管,极少部分靠腹血管。背血管(图4)为背板中央一条纵贯头部至腹末的血管。心脏由位于1~9腹节的8个心室及心脏瓣、心门组成。腹血管为腹部体腔中线上一条纵贯头部至腹末的管状器官。辅搏器位于蜻蜓稚虫的触角、足或翅膀的基部。
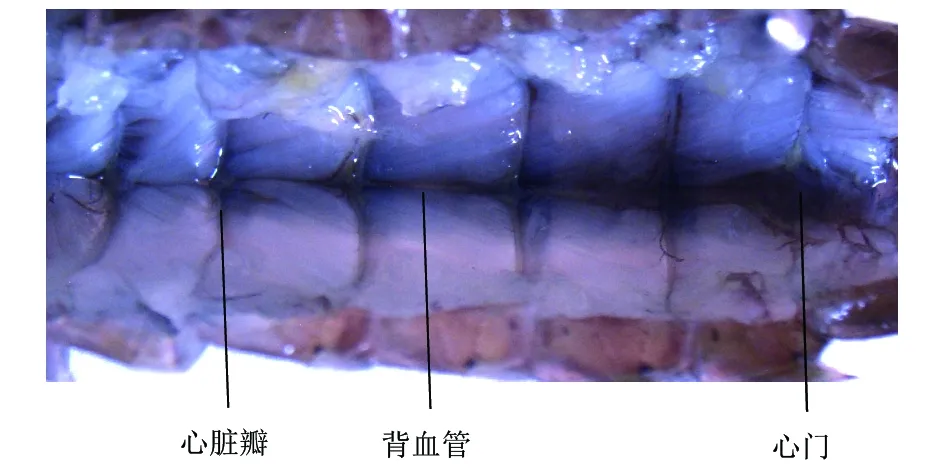
图4 碧伟蜓稚虫的背血管及心门Fig.4 Digestion and excretory system of larvae of Anax parthenope julius
2.3.4呼吸系统。碧伟蜓稚虫的呼吸系统包括气管及直肠鳃两大部分(图5)。气管位于消化道两侧的白色管状结构,纵贯头、胸及腹部,通过中胸气门、腹部的各节气门与外界相通,离开水面后也能自主呼吸。直肠鳃位于稚虫的直肠部位,是由直肠的特殊上皮细胞变形而成的众多的鳃蓝子集合体。
2.3.5神经系统。碧伟蜓稚虫的神经系统包括神经及感觉器官两部分。神经从上而下由脑、围咽神经节、食道下神经节、胸神经节及腹神经节及神经组成,分别位于头顶、前胸、中胸、后胸及腹部,除脑及围咽神经节外,其余均位于纵贯全身的背血管下方,呈白色的线状。感觉器官有触角、复眼及单眼等,均集中在头部。

图5 碧伟蜓稚虫的直肠鳃及气管Fig.5 Rectal gill and trachea of Anax parthenope julius
2.3.6生殖系统。昆虫的生殖系统包括外生殖器和内生殖器。而蜻蜓稚虫外生殖器官发育还不成熟,从外观上看,雄性在第9腹节腹板中央上有一个黑斑(图6A),雌性是一个长条形的生殖板(图6B),以此来辨别稚虫的性别。雄性的精巢发育不成熟,仅能观察到雌性稚虫的卵巢。
碧伟蜓稚虫雌性生殖系统包括1对卵巢、2根位于侧面的侧输卵管和一根开口于生殖板处的中输卵管。卵巢位于蜻蜓稚虫消化道的两侧,由卵巢管组成,附着有脂肪体(图7)。

注:A-雄性,B-雌性Note:A-male ,B-female图6 碧伟蜓稚虫腹部末端腹面观(示雌雄外生殖孔)Fig.6 Terminal abdomen and ventral side of Anax parthenope julius(female and male external gonopore)
侧输卵管在卵巢两侧,连接卵巢和中输卵管,是1对等长的管道。中输卵管为侧面2条侧输卵管并入的地方,开口于第9腹节处,内陷形成蜻蜓生殖腔,在中输卵管下方有一开口于生殖板的结构。
图7 碧伟蜓稚虫雌性卵巢Fig.7 Female ovary of Anax parthenope julius1
[1] 王治国.中国蜻蜓分类名录(蜻蜓目)[J].河南科学,2017,35(1):48-77.
[2] 杨玉琼,刘红,杨姗.蜻蜓蛹中微量元素及其营养价值[J].微量元素与健康研究,2009,26(6):36-37.
[3] 冯颖,陈晓鸣,王绍云,等.蜻蜒目的3种食用种类与营养价值[J].林业科学研究,2001,14(4):421-424.
[4] 蒋筠雅,何钊,赵敏,等.6种常见食用蜻蜓稚虫含油率与脂肪酸组成分析[J].中国油脂,2017,42(3):135-139.
[5] CLARK T F,SAMWAYS M J.Dragonflies(Odonata)as indicators of biotope quality in the Kruger National Park,South Africa [J].Joumal of applied ecology,1996,33(5):1001-1012.
[6] CHOVANEC A,WARINGER J,RAAB R,et al.Lateral connectivity of a fragmented large river system:Assessment on a macroscale by dragonfly surveys(Insecta:Odonata)[J].Aquatic conservation:Mariue and freshwater ecosystems,2004,14(2):163-178.
[8] FRANCO G M S,TAKEDA A M.Spatial and temporal variation of Odonata larvae associated with macrophytes in two floodplain lakes from the upper Paraná River,Brazil [J].Acta Sci,2002,24(2):345-351.
[9] GUILLERMO-FERREIRA R,BISPO P C.Description of the larva ofTelebasisgriffinii(Martin,1896)(Zygoptera:Coenagrionidae)[J].Odonatologica,2013,42(4):403-407.
[10] HONKANEN M,SORJANEN A,MÖNKKONEN M.Deconstructing responses of dragonfly species richness to area,nutrients,water plant diversity and forestry[J].Oecologia,2011,166(2):457-467.
[11] TILLYARD R J.The biology of dragonflies[M].Cambridge:Cambridge University Press,1917.
[12] NEEDHAM J G A.Manual of the dragonflies of China.A monographic study of the Chinese odonata[M].Peiping:Fan Memorial Institute of Biology,1930.
[13] 赵修复.中国春蜓分类[M].福州:福建科学技术出版社,1990.
[14] 江尧桦.六种蜻蜓及豆娘稚虫雌雄特征区别的研究[J].昆虫知识,1996,33(1):45-47.
[15] ZHANG H M,TONG X L.Descriptions ofBoyeriakarubeiYokoi andPeriaeschnaf.flintiAsahina larvae from China(Anisoptera:Aeshnidae)[J].Odonatologica,2011,40(1):57-65.
[16] 钟觉民.幼虫分类学[M].北京:农业出版社,1990:17-20.
[17] 刘保平.伏击法捕捉碧伟蜓技术研究[J].安徽农业科学,2013,41(28):11383-11385.
[18] CORBET P S.Dragonflies behavior and ecology of odonata[M].New York:Cornell University Press,1999.
