放牧强度对青藏高原高寒矮嵩草草甸氧化亚氮释放的影响*
2018-01-31李红琴未亚西贺慧丹杨永胜李英年
李红琴,未亚西,贺慧丹,杨永胜,李英年
放牧强度对青藏高原高寒矮嵩草草甸氧化亚氮释放的影响*
李红琴1,3,未亚西1,2,贺慧丹1,2,杨永胜1,3,李英年1,3
(1.中国科学院西北高原生物研究所,西宁 810001;2.中国科学院大学,北京 100049;3.中国科学院高原生物适应与进化重点实验室,西宁 810001)
依托青藏高原东北隅高寒矮嵩草草甸的5a放牧强度(禁牧、轻度放牧、中度放牧、重度放牧)试验平台,2016年在植物生长季的6-9月,基于静态暗箱-气相色谱法,测定N2O的释放特征及相应的环境、生物因子,探讨放牧强度对高寒草甸N2O释放特征的影响及其内在环境生物驱动机制。结果表明:环境、生物因子中仅表层土壤容积含水量、土壤容重及土壤有机碳含量对放牧强度响应显著(P<0.05)。高寒草甸N2O释放的季节特征表现出生长季的早期和晚期相对较高的“U”型趋势。禁牧样地N2O释放速率最小,极显著(P<0.01)低于其它3个放牧样地。高寒草甸N2O释放强度与放牧强度间表现出正相关趋势(R= 0.49, P<0.01)。相关分析表明,表层土壤温度是高寒草甸N2O释放速率的主要影响因子,但放牧强度改变了土壤温度的影响程度。中短期放牧管理改变了高寒草甸植被生长季N2O释放速率,但未改变其释放的季节特征。禁牧管理提高了土壤温度,进而显著降低植被生长季N2O释放强度。
氧化亚氮;放牧强度;高寒草甸;静态暗箱-气相色谱法;Spearman秩相关
氧化亚氮(N2O)是仅次于二氧化碳(CO2)和甲烷(CH4)的第三大重要温室气体,具有较大的温室效应潜能。N2O能参与许多重要的大气化学反应,被认为是21世纪最重要的消耗臭氧层物质之一[1]。土壤N2O释放主要来源于土壤微生物的硝化作用和反硝化作用,其增温效应最高,对全球温室效应的贡献度约为5%[2-3]。全球气候变暖使中国的经济发展面临着严峻威胁,N2O释放规律的研究受到广泛关注。N2O主要来源于陆地土壤,约占其释放量的65%~70%。土壤中N2O的产生释放与土壤生物化学过程、土地利用和人类活动,特别是土壤环境有着极其密切的关系[4]。
草地约占中国陆地面积的40%,是最重要的植被类型,N2O的释放强度约为0.30kgN·hm−2·a−1[5-6]。高寒草地是青藏高原的主体植被类型,约占高原面积的2/3[7],是中国草地N2O的重要释放源[8],其释放量略低于内蒙古草原地区[6, 9]。土壤系统的N2O的释放贡献率高达高寒草地系统的77%[10-11]。
放牧是草地生态系统最重要的利用方式之一,但过度放牧是人类施与草地生态系统最强大的影响因素。放牧强度明显影响高寒草地稳定性和土壤性质[12-13],势必影响草地生态系统的N2O释放。由于气候状况、植被结构、土壤理化、放牧管理、演替状态的影响,草地N2O的释放具有较大的时空变异性[14-18]。相对于碳、水循环研究的广度和深度[19-21],高寒草地N2O的释放则由于观测设备和方法的限制而略显不足[8, 10]。本研究通过在青海海北同一牧户的草地,建立不同放牧强度试验地,研究植被生长季(6−9月)N2O的释放特征及与环境因子、土壤理化性质的关系,以期对放牧管理方式提出行之有效的措施和依据,同时为预测高寒草甸对气候变化和人类活动的响应提供理论依据[22]。
1 材料与方法
1.1 试验地概况
试验样地位于青海海北高寒草甸生态系统国家野外科学观测研究站(海北站)。海北站(37°37′N,101°19′E,3200m)地处青藏高原东北隅的祁连山高地,具有明显的高原大陆性季风气候。区域仅有冷暖两季之分,年平均气温−1.7℃,夏季温暖多雨,冬季寒冷干燥。年降水量约580mm,植物生长季集中了全年降水的近80%。土壤为暗沃寒冻雏形土(),土壤富含有机质但缺乏可利用的氮[23]。以矮嵩草()为建群种,主要优势种为垂穗披碱草()、异针茅()、麻花艽(a)、鹅绒委陵菜()和紫羊茅()等。
1.2 实验设计
放牧强度实验平台于2011年8月在海北站的高寒矮嵩草草甸建立,建立前为牧户冬季放牧草场。参照以往研究放牧强度的经验[24],设置禁牧(对照,CK)、轻度放牧(light grazing,LG,4.5只羊·hm−2)、中度放牧(moderate grazing,MG,7.5只羊·hm−2)和重度放牧(heavy grazing,HG,15只羊·hm−2)4种放牧强度。用网围栏将草地围封为面积不同的4个样地,每个样地均放牧2只当地藏系绵羊,放牧时间为9月20日−翌年5月31日,与牧民放牧时间一致。鉴于高寒草地N2O释放存在明显的年际特征,且针对多年的数据分析时,主流的统计做法多将年作为一个变量因子进行分析,探讨年与主要控制因子(如增温梯度、降水梯度、放牧梯度)的主效应和交互效应[25]。而2016年的年均气温(−1.2℃)和年降水量(521.3mm)与研究地点(海北站)的多年平均气温(−1.1℃)和降水量(511.7mm)基本持平,故本研究选用2016年数据对研究区N2O释放的平均年际特征进行分析。连续进行5a放牧强度梯度实验后,于2016年6−9月进行样品采集和相关生物、土壤要素的测定。
1.3 要素监测与分析
N2O释放监测:在每个放牧强度样地设置3个重复,3个重复间隔尽量远以降低假重复的影响。基于国际通用的静态暗箱-气相色谱法进行N2O气体的采集及测定[26]。不锈钢采集底座为0.5m×0.5m×0.1m,垂直砸入地表5cm;箱体大小为0.5m×0.5m×0.5m,在箱体侧面内置两个小风扇,保障箱内气体混合均匀。为尽量减小采样箱内部的温度变化,在采样箱外部包裹一层2cm厚的海绵保温材料。测量时利用水封保证静态暗箱内部的气密性[8]。
在6−9月(植被生长季)每月中旬选择典型晴天于9:00−11:00[9],采用带有三通阀的100mL注射器抽气后,关闭稳压管,密闭10min后再使用另一气密注射器采气,每个箱体采集四针气体样品,用于测定N2O的体积混合比的变化速率。采样结束后,移开密闭采样箱,立即采用便携式铂电阻数字温度计和时域反射仪,测定土壤5cm温度和容积含水量。N2O气体混合比的测定采用气相色谱(HP4890D,Agilent)在室内完成,内装电子捕获检测器。测定的色谱条件为:柱温和检测温度分别为70℃和300℃;标气浓度为355nL×L−1,最小因子检测限为±5nL×L−1。
N2O释放强度(F,mg·m−2·h−1)的计算式为


生物量和土壤理化指标测定:在采集气体样本的同时,在每个样地进行植被地上、地下生物量和表层土壤有机碳、土壤硝态氮(NO– 3-N)和铵态氮(NH+ 4-N)及全磷样品的采集。植被地上、0−10cm地下部分生物量测定分别采用50cm×50cm的样方框和内径7cm的根钻,基于标准收获法获取[27],3个重复。采集的0−10cm表层土壤样品,在试验室内完成碳、氮、磷的分析测试。其中,有机碳和全磷的测定分别采用重铬酸钾法和H2SO4-HCLO4-钼锑抗比色法;土壤硝态氮(NO– 3-N)和铵态氮(NH+ 4-N)分别采用紫外分光光度法和靛酚蓝比色法[28]。
0−10cm表层土壤容重采用环刀法取样,带回实验室测定,考虑到土壤容重的季节变化较小,仅在2016年8月上旬测定一次。
1.4 统计分析
以放牧强度为单因素,环境、生物要素为因变量进行单因素方差分析(采用最小方差法LSD进行放牧强度之间的多重比较)探讨放牧管理对环境、生物要素的驱动强度。再以放牧强度和季节梯度为固定因素,基于一般线性模型分析放牧强度和季节因子对N2O释放速率的贡献强度。由于地上生物量等因子的数据分布不能满足正态要求,故采用Spearman秩相关分析环境、生物要素与N2O释放速率的相关程度。同时,依据N2O的全球变暖潜能值(Global warming potential,GWP)为296,其释放量可转化为296倍CO2的当量值,探讨N2O释放对高寒草甸系统CO2气体释放量的贡献程度。统计分析和参数拟合在OriginLab 8.5完成。
2 结果与分析
2.1 放牧强度对植被生长及其土壤环境的影响
由表1可见,经过5a的冬季放牧管理试验,高寒草甸表层土壤容积含水量、土壤容重及土壤有机碳含量差异显著,其它环境因子和土壤理化性状在不同放牧强度之间均无显著差异。2016年6−9月平均地上生物量表现出轻牧样(LG)地最大,重牧样地(HG)最小的趋势,但统计分析表明不同放牧强度之间差异不显著。月均表层地下生物量在中牧样地(MG)最高,仅显著高于轻牧样地约14%,但与重牧、禁牧样地(CK)无显著差异。轻牧样地的表层土壤(0−10cm)容重最小,显著低于重度放牧和禁牧样地。禁牧样地的表层土壤容重相对最大,但与重度放牧和中度放牧差异不显著。表层土壤有机碳和表层土壤容重表现出相似的变化趋势,轻牧样地显著最小,重牧样地相对最大。表层土壤硝态氮、铵态氮和全磷含量未随放牧强度表现出显著差异,呈现出轻牧样地相对较小,禁牧样地相对较大的趋势。由于植被生物量和土壤理化性质随放牧强度呈现出一定的差异,造成各处理土壤水热环境略有区别。表层土壤含水量在中牧和重牧样地最小,显著低于禁牧和轻牧样地。但可能由于高原太阳辐射较强,导致土壤温度未呈现显著差异。因此,5a放牧强度可能仅通过践踏和选择性采食作用对高寒草甸植被生物量和土壤水、碳产生一定影响。
2.2 放牧强度对草地N2O释放速率的影响
高寒矮嵩草草甸生长季的N2O释放速率平均为0.035±0.015mg·m−2·h−1。放牧强度对高寒草甸生长季不同月份的N2O释放速率均产生了显著影响(图1)。高寒草甸N2O释放速率在9月相对最高(0.047± 0.016mg·m−2·h−1),6月次之(0.038±0.012mg·m−2·h−1),但二者无显著差异,且均显著(P<0.05)高于7月(0.025±0.011mg·m−2·h−1)和8月(0.028±0.001mg·m−2·h−1)的N2O释放速率。
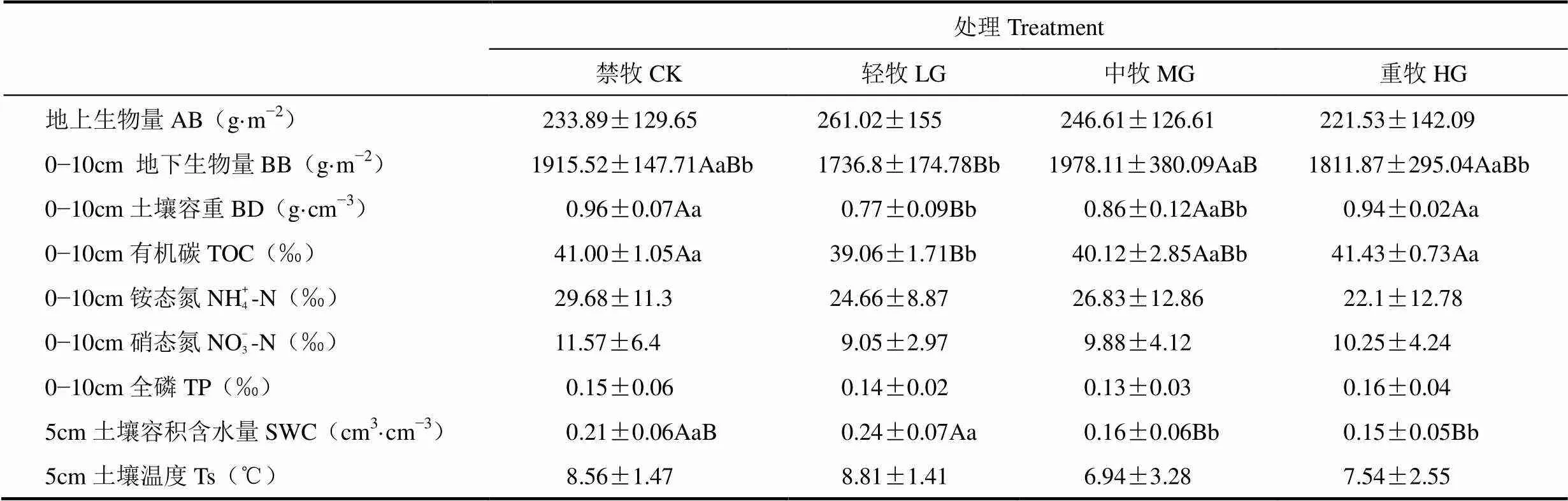
表1 生长季6−9月不同放牧强度样地月平均植被生物量和土壤理化指标比较(平均值±标准差)
注:小写字母表示处理间在0.05水平上的差异显著性,大写字母表示处理间在0.01水平上的差异显著性。下同。
Note:AB: above ground biomass; BB: below ground biomass; BD: bulk density; TOC: total organic carbon; TP: total phosphorus; SWC: soil water content; Ts: temperature of soil. Lowercase indicates the difference significance among treatments at 0.05 level, and capital letter indicates the difference significance among treatments at 0.01 level. The same as below.
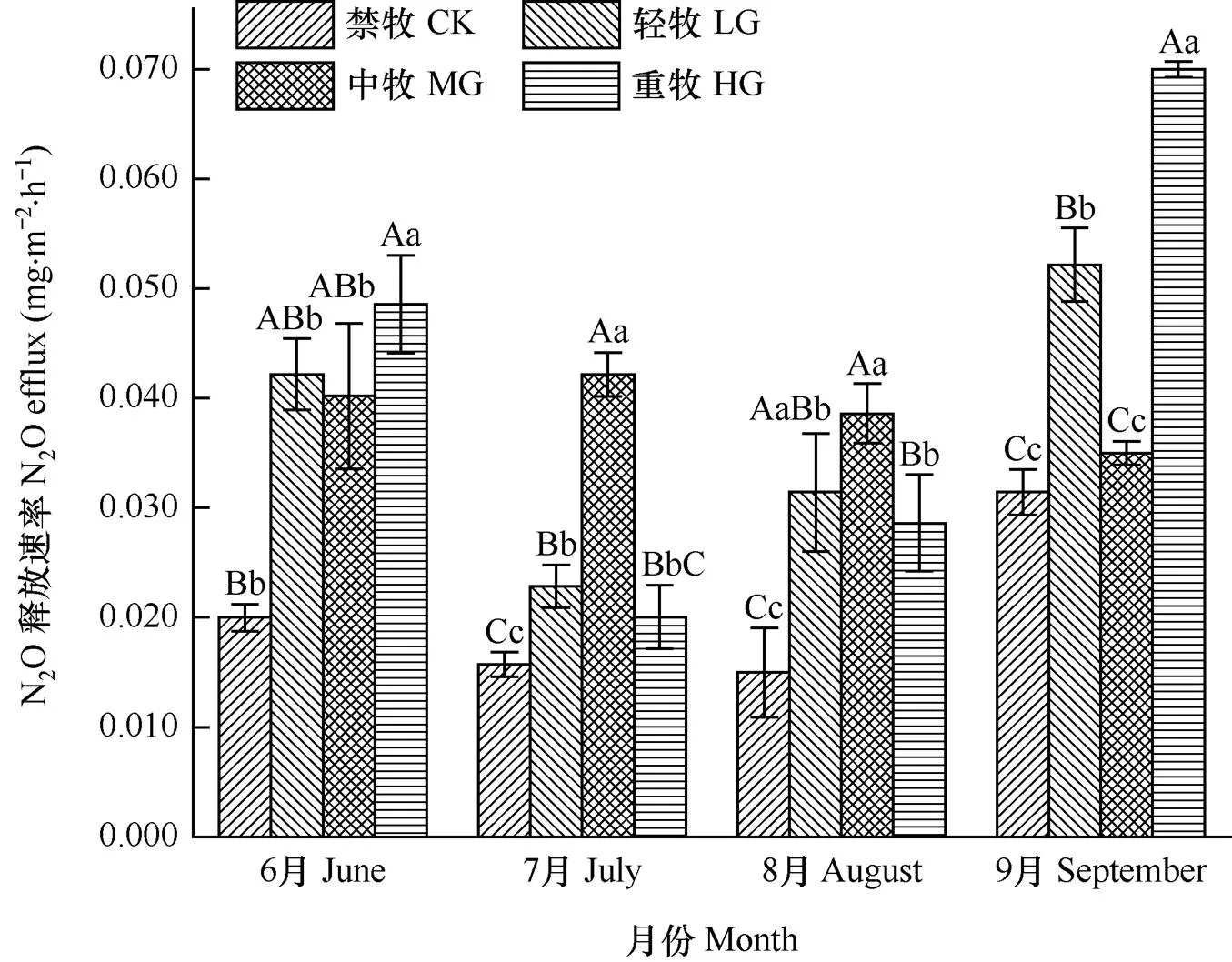
图1 各月不同放牧强度下高寒草甸N2O释放速率的比较
注:误差线为标准误
Note: The error bar is standard error
禁牧样地的N2O释放速率最小(0.021±0.007 mg·m−2·h−1),极显著(P<0.01)小于放牧样地。轻牧、中牧和重牧样地N2O释放速率分别为0.037±0.012、0.036±0.0074、0.042±0.021mg·m−2·h−1,但三者之间无显著差异。高寒草甸N2O释放速率与放牧强度和季节梯度的一般线性模型(R2=0.94,P<0.001)表明放牧强度、季节梯度及二者的交互作用均十分显著(P<0.01),其中季节梯度的主效应略大于放牧强度,但二者主效应远大于二者的交互效应。
将每月N2O的释放速率作为该月的平均值,经简单计算可知禁牧、轻牧、中牧和重牧高寒矮嵩草草甸在植被生长季6−9月的N2O的累计释放量分别为89.7、162.2、170.3和182.5mg·m−2,呈现出随放牧强度增加而增大的趋势,按照296倍CO2的当量值计算,其累计CO2释放当量分别为26.5、48.0、50.4和54.0g·m−2。可见,高寒矮嵩草草甸N2O释放强度由放牧管理和季节变化共同调控。在季节尺度上表现出在生长季早期(6月)和晚期(9月)较高,在放牧强度中呈现出禁牧样地较小的趋势。
2.3 土壤环境对N2O释放速率的影响

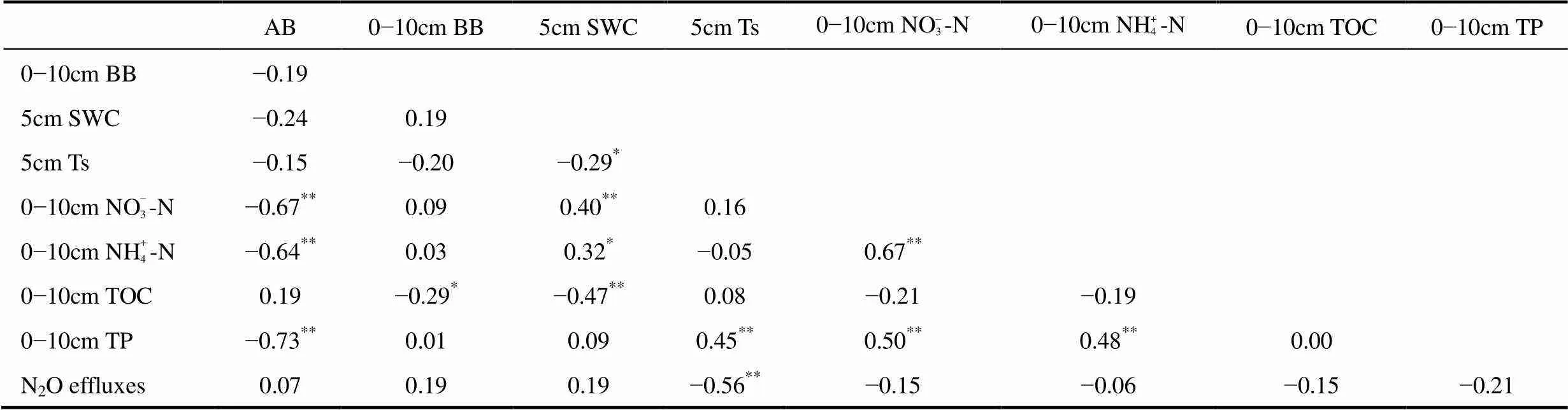
表2 环境、生物要素、N2O释放速率的Spearman秩相关系数(N=48)
注:*、**分别表示相关系数通过0.05、0.01水平的显著性检验。
Note:*is P<0.05,**is P<0.01.
3 结论与讨论
3.1 讨论
高寒矮嵩草草甸生长季N2O释放速率平均为0.035±0.015mg·m−2·h−1,该研究结果与海北高寒金露梅灌丛草甸[8, 11]、红原高寒高山嵩草草甸[29]及锡林郭勒羊草草原[9]接近,但显著高于纳木错高寒针茅草原[9]。因此,气候因素对高寒草地N2O释放强度的影响可能大于植被类型的作用[30]。本研究的N2O释放表现出生长季早期和晚期相对较高的“U”型趋势,这可能主要由于高寒系统N2O释放和温度负相关所致[9, 29],而且温度较低的非生长季和夜间N2O释放远高于生长季的释放也印证了这种负相关现象[8-9]。在植被生长季中,轻牧管理下高寒矮嵩草草甸N2O的释放量为162.2mg·m−2,其CO2当量(296倍)可换算为48g·m−2,占植被生长季CO2吸收总量[31](146gCO2·m−2)的32%,略低于荷兰高寒人工草地45%的贡献率[15]。但由于N2O释放的时空异质性极强[17, 30],需要进行更长期的研究来评估其对高寒草甸生态系统温室效应的贡献。
温度是影响N2O释放量的重要因子,调节着土壤整体环境,以其影响土壤中微生物活性,显著影响N2O的释放量。N2O主要来源是土壤微生物的硝化和反硝化过程[32]。高寒草地土壤中有机物质的分解和矿化作用是土壤中NO– 3-N和NH+ 4-N主要来源,而土壤温度,相对土壤含水量是调节高寒系统土壤有机质分解和矿化过程的主要因素[33-34]。本研究发现仅土壤温度与高寒草地N2O释放显著负相关,暗示高寒嵩草草甸的N2O主要来源于反硝化作用,这与海北、纳木错高寒草地的研究结果相似[9, 30]。这主要由于研究区土壤含水量较高,易形成厌氧微环境,从而利于土壤微生物的反硝化[9],而反硝化产物的N2/N2O比例与温度呈负相关关系[33]也印证了土壤温度的负作用。表层土壤的NO– 3-N含量与N2O释放呈现微弱的负相关关系,暗示高寒草地反硝化作用的控制因子较为复杂。
禁牧管理显著降低了高寒草地N2O释放强度,这主要可能由于禁牧样地常年的凋落物阻碍了N2O的释放过程[9]。而轻牧、中牧和重牧样地的N2O释放没有显著差异,这主要由于植被系统(绿色植被和地下根系)的根冠较大,土壤微生物硝化和反硝化的底物相对充足,导致其对高寒草甸系统N2O释放强度的贡献较小[10-11]。加之植物物种组成变化相对较小,也导致了放牧管理对草地系统N2O释放的驱动有限[35]。高寒矮嵩草草甸N2O释放和放牧强度呈现正相关趋势,与内蒙古半干旱温性草原的研究结果相反[16],这可能由于干旱温性草原主要以硝化反应为主[6, 9]。因此,中短期放牧管理可能是通过家畜的践踏作用改变了表层土壤的理化性质来调控土壤中氧气、微生物种类和反应物基质,进而影响高寒草甸的N2O释放特征[14, 36]。但N2O释放的控制因子较多,且因子的交互作用可能会掩盖或抵消其它因子的作用[3, 37],加之生态系统对放牧年限的非线性响应[34],因此,需要更进一步研究高寒草地N2O释放特征对放牧管理的响应机制。
3.2 结论
5a放牧强度试验仅对表层土壤容积含水量、土壤容重及土壤有机碳含量产生了显著影响,暗示高寒系统碳、水交换对放牧管理较为敏感。禁牧、轻牧、中牧和重牧的高寒草甸N2O释放强度均表现出生长季早期和晚期较高的季节特征。禁牧样地N2O释放强度显著低于其它3个样地,而其它3个处理则无显著差异,仅表现出正相关于放牧强度。N2O释放强度与表层土壤温度显著负相关。
[1]Ravishankara A R,Daniel J S,Portmann R W.Nitrous Oxide (N2O):the dominant ozone-depleting substance emitted in the 21st century[J].Science,2009,326(5949):123-125.
[2]IPCC.Climate change 2013-the physical science basis:working group I contribution to the fifth assessment report of the intergovernmental panel on climate change[M].Cambridge, United Kingdom and New York,USA:Cambridge University Press,2013:3-29.
[3]Brown J R,Blankinship J C,Niboyet A,et al.Effects of multiple global change treatments on soil N2O fluxes[J].Biogeochemistry, 2012,109(1):85-100.
[4]李香兰,徐华,曹金留,等.水分管理对水稻生长期N2O释放的影响[J].土壤,2006,38(6):703-707.
Li X L,Xv H,Cao J L,et al.Effect of water management on N2O emission in rice-growing season[J]. Soils,2006,38(6): 703-707.(in Chinese)
[5]Cai Z.Greenhouse gas budget for terrestrial ecosystems in China[J].Science China:Earth Sciences,2012,55(2):173-182.
[6]罗云鹏.中国森林和草地氧化亚氮释放估算[D].杨凌:西北农林科技大学,2015.
Luo Y P.Estimating Nitrous Oxide emission from Chinese forest and grassland[D].Yangling:Northwest A&F University, 2015.(in Chinese)
[7]张镱锂,祁威,周才平,等.青藏高原高寒草地净初级生产力(NPP)时空分异[J].地理学报,2013,68(9):1197-1211.
Zhang Y L,Qi W,Zhou C P,et al.Spatial and temporal variability in the net primary production(NPP) of alpine grassland on Tibetan Plateau from 1982 to 2009[J].Acta Geographica Sinica, 2013,68(9):1197-1211.(in Chinese)
[8]Du Y,Cui Y,Xu X,et al.Nitrous oxide emissions from two alpine meadows in the Qingha-Tibetan Plateau[J].Plant and Soil, 2008,311(1):245-254.
[9]李梓铭,杜睿,王亚玲,等.中国草地N2O通量日变化观测对比研究[J].中国环境科学,2012,32(12):2128-2133.
Li Z M,Du R,Wang Y L,et al.Comparison of diurnal variation of nitrous oxide fluxes from grassland of China[J].China Environmental Science,2012,32(12):2128-2133.(in Chinese)
[10]Pei Z Y,Ouyang H,Zhou C P,et al.N2O exchange within a soil and atmosphere profile in alpine grasslands on the Qinghai-Xizang Plateau[J].Acta Botanica Sinica,2004,46(1): 20-28.
[11]Du Y G,Cao G M,Deng Y C,et al.Contribution of the vegetation layers in the nitrous oxide emission from alpine Kobresia humilis Serg.meadow ecosystem on the Tibetan Plateau[J].Polish Journal of Ecology,2010,58(1):115-124.
[12]Hassink J.Effect of soil texture on the size of the microbial biomass and on the amount of C and N mineralized per unit of microbial biomass in dutch grassland soils[J].Soil Biology & Biochemistry,1994,26(11):1573-1581.
[13]Kroon P S,Schrier Uijl A P,Hensen A,et al.Annual balances of CH4and N2O from a managed fen meadow using eddy covariance flux measurements[J].European Journal of Soil Science,2010,61(5):773-784.
[14]Wolf B,Zheng X,Brüggemann N,et al.Grazing-induced reduction of natural nitrous oxide release from continental steppe[J].Nature,2010,464(7290):881-884.
[15]付晓青,李勇.土壤氧化亚氮释放时空变异性及其方法研究进展[J].生态学杂志,2012,31(3):724-730.
Fu X Q,Li Y.Spatiotemporal variability of soil nitrous oxide emission and its study methods:a review[J].Chinese Journal of Ecology,2012,31(3):724-730.(in Chinese)
[16]Merbold L,Eugster W,Stieger J,et al.Greenhouse gas budget (CO2,CH4and N2O) of intensively managed grassland following restoration[J].Global Change Biology,2014,20 (6):1913.
[17]Theurillat J P,Guisan A.Potential impact of climate change on vegetation in the Europeanalps:a review[J].Climatic Change, 2001,50(1):77-109.
[18]Li H,Zhang F,Li Y,et al.Seasonal and inter-annual variations in CO2fluxes over 10 years in an alpine shrubland on the Qinghai-Tibetan Plateau,China[J].Agricultural and Forest Meteorology,2016,(228-229):95-103.
[19]Li J,Zhang F,Lin L,et al.Response of the plant community and soil water status to alpine Kobresia meadow degradation gradients on the Qinghai-Tibetan Plateau,China[J].Ecological Research,2015,30:589-596.
[20]曹广民,龙瑞军.放牧高寒嵩草草甸的稳定性及自我维持机制[J].中国农业气象,2009,30(4):553-559.
Cao G M,Long R J.System stability and its self-maintaining mechanism by grazing in alpine Kobresia meadow[J]. Chinese Journal of Agrometerology,2009,30(4):553-559.(in Chinese)
[21]Xie R,Wu X Q.Effects of grazing intensity on soil organic carbon of rangelands in Xilin Gol League,Inner Mongolia, China[J].Journal of Geographical Sciences,2016(11):1550- 1560.
[22]丁仲礼,段晓男,葛全胜,等.2050年大气CO2浓度控制:各国释放权计算[J].中国科学(D辑),2009,39(8):1009-1027.
Ding Z L,Duan X N,Ge Q S,et al.Control of atmospheric CO2concentrations by 2050:a calculation on the emission rights of different countries[J].Science in China(Series D:Earth Science),2009,39(8):1009-1027.(in Chinese)
[23]周兴民.中国嵩草草甸[M].北京:科学出版社,2001:1-297.
Zhou X M.Kobresia meadow in China[M].Beijing:Science Press,2011:1-297.(in Chinese)
[24]王启基,周立,王发刚.放牧强度对冬春草场植物群落结构及功能的效应分析[M].北京:科学出版社1995:353-364.
Wang Q J,Zhou L,Wang F G.Effect of grazing intensity on plant community structure and function in winter and spring grassland[M].Beijing:Science Press,1995:353-364.(in Chinese)
[25]Gao X,Thomas B W,Beck R,et al.Long-term grazing alters soil trace gas fluxes from grasslands in the foothills of the Rocky Mountains,Canada[J].Land Degradation & Development, 2017.DOI: 10.1002/ldr.2664.
[26]李志安,邹碧,曹裕松,等.地面氧化亚氮释放静态箱测定技术[J].土壤与环境,2002,11(4):413-416.
Li Z A,Zou B,Cao Y S,et al.Technique of static chamber in determining nitrous oxide emission from land surface[J].Soil and Environmental Science,2002,11(4):413-416.(in Chinese)
[27]董鸣.陆地生物群落调查观测与分析[M].北京:中国标准出版社,1996.
Dong M.Survey,observation and analysis of terrestrial biocommunities[M].Beijing:China Standards Press,1996.(in Chinese)
[28]刘光崧,蒋能慧,张连第.中国生态系统研究网络观测与分析标准方法:土壤理化分析与剖面描述[M].北京:中国标准出版社,1996.
Liu G S,Jiang N H,Zhang L D.Soil physical and chemical analysis and description of soil[M].Beijing:China Standards Press,1996.(in Chinese)
[29]詹伟.高寒草甸放牧草地土壤氧化亚氮释放研究[D].杨凌:西北农林科技大学,2015.
Zhan W.Grazing effects on Nitrous Oxide emission of alpine meadow in Tibet Plateau[D].Yangling:Northwest A&F University,2015.(in Chinese)
[30]Lin X,Wang S,Ma X,et al.Fluxes of CO2,CH4,and N2O in an alpine meadow affected by yak excreta on the Qinghai-Tibetan plateau during summer grazing periods[J]. Soil Biology & Biochemistry,2009,41(4):718-725.
[31]Kato T,Tang Y H,Gu S,et al.Temperature and biomass influences on interannual changes in CO2exchange in an alpine meadow on the Qinghai-Tibetan Plateau[J].Global Change Biology,2006,12(7):1285-1298.
[32]Schimel J P,Jay G.Microbial community structure and global trace gases[J].Global Change Biology,1998,4(7):745-758.
[33]Mcneill A,Unkovich M.The Nitrogen cycle in terrestrial ecosystems[M].Nutrient Cycling in Terrestrial Ecosystems, Marschner P,Rengel Z,Berlin,Heidelberg:Springer Berlin Heidelberg,2007:37-64.
[34]Cowan N J,Levy P E,Famulari D,et al.The influence of tillage on N2O fluxes from an intensively managed grazed grassland in Scotland[J].Biogeosciences,2016,13(16):4811-4821.
[35]Abalos D,De Deyn G B,Kuyper T W,et al.Plant species identity surpasses species richness as a key driver of N2O emissions from grassland[J].Global Change Biology,2014,20 (1):265-275.
[36]Oenema O,Velthof G L,Yamulki S,et al.Nitrous oxide emissions from grazed grassland[J].Soil Use & Management, 1997,13(4):288-295.
[37]Shi F,Chen H,Chen H,et al.The combined effects of warming and drying suppress CO2and N2O emission rates in an alpine meadow of the eastern Tibetan Plateau[J].Ecological Research,2012,27(4):725-733.
Effects of Grazing Density on Nitrous Oxide Effluxes in Alpine Kobresia Humilis Meadow on the Qinghai-Tibetan Plateau
LI Hong-qin1,3, WEI Ya-xi1,2, HE Hui-dan1,2, YANG Yong-sheng1,3, LI Ying-nian1,3
(1. Northwest Institute of Plateau Biology,Chinese Academy of Sciences,Xining 810001,China;2. University of Chinese Academy of Sciences,Beijing 100049;3.Key Laboratory of Adaptation and Evolution of Plateau Biota,Chinese Academy of Sciences,Xining 810001)
Based on the experimental platform of 5 years grazing density including fencing (CK), light grazing (LG), moderate grazing (MG) and heavy grazing (HG) over the alpine Kobresia meadow on the northeastern Qinghai-Tibet Plateau, the characteristics of N2O effluxes and its main environmental factors of different grazing density were studied by static closed chamber-gas chromatography method from June to September in 2016.The results showed that only the topsoil water content, topsoil bulk density and topsoil organic carbon content responded significantly (P<0.05) to the grazing density. The seasonal patterns of N2O effluxes showed the “U” shape with relatively higher rate during early and late stage of growing season in alpine meadow. The N2O effluxes in fencing treatments was clearly (P<0.01) lower than that of other three grazing gradients. There was a little positive trend between N2O effluxes and grazing density. The surface soil temperature played a predominated role in the seasonal variations of the N2O effluxes in alpine meadow and its magnitude was influenced by grazing density. The results suggested that only the magnitude, not the direction, of N2O effluxes was significantly influenced by the medium (short) -term grazing management during growing season. The fencing management could significantly reduce the N2O emission caused by improving the soil temperature.
Nitrous Oxide;Grazing density;Alpine meadow;Static closed chamber-gas chromatography method;Spearman's rank correlation coefficient
10.3969/j.issn.1000-6362.2018.01.004
李红琴,未亚西,贺慧丹,等.放牧强度对青藏高原高寒矮嵩草草甸氧化亚氮释放的影响[J].中国农业气象,2018,39(1):27-33
2017−03−27
青海省国际科技合作项目(2015−HZ−804);国家自然科学基金项目(31300385;31270523)
李红琴(1981−),女,博士,副研究员,主要从事全球变化生态学研究。E-mail: lihongqin_00@126.com
