长鳍吻免疫因子基因cDNA部分序列克隆及其组织表达差异
2018-01-27李学梅吴兴兵王旭歌朱永久杨德国
李学梅,吴兴兵,王旭歌,朱永久,杨德国
(中国水产科学研究院长江水产研究所,农业部淡水生物多样性保护重点实验室,武汉 430223)
应激的监测不仅在于判断应激发生与否及应激程度,还在于判断鱼体的敏感性,对改善鱼类养殖管理,提高养殖效益具有重要意义。目前,长鳍吻的研究主要集中在资源量调查、种群生态学、生物学及人工繁殖技术等[1,5-7]。关于长鳍吻应激监测方面的研究尚未见报道。本研究以常见应激因子HSP70 (热休克蛋白)、特异性免疫因子IgM(免疫球蛋白)、抑炎性因子IL-10(白细胞介素10)和促炎性因子IL-1b(白细胞介素1)为监测对象,初探感染和未感染小瓜虫的长鳍吻皮肤和肠道组织中这些免疫因子的表达差异,旨在为长鳍吻应激监测方面提供理论依据。
1 材料与方法
1.1 材料

表1 不同处理组长鳍吻体长体重(平均值±标准差) Tab.1 The body length and body weight of R.ventralis in different group (Mean±SD)
1.2 引物合成
在GeneBank 数据库中下载鲤(Cyprinuscarpio)、斑马鱼(Daniorerio)、草鱼(Ctenopharyngodonidella)、鲢(Hypophthalmichthysmolitrix)、鲫(Carassiusauratus)的HSP70、IgM、IL-10和 IL-1b基因的cDNA核苷酸序列,通过Clustal X2比对分析保守区,应用 Primer Premier 5.0 软件,按照引物设计的基本要求,设计并合成4对引物(表2),用于扩增长鳍吻HSP70、IgM、IL-10和 IL-1b基因的保守片段。根据获得的保守片段,设计出HSP70-S/HSP70-A、IgM-S/IgM-A、IL10-S/IL10-A和IL1b-S/IL1b-A等 4对荧光定量表达的基因特异性引物,用实验室已有的四川裂腹鱼的βactin-S/βactin-A为荧光定量表达的内参引物(表3)。因目前长鳍吻相关的内参基因尚未有报道,因此选择与吻属同源性较高的已有报道的裂腹鱼属(同源性99%)βactin基因作为内参进行初探,且对该内参进行了预实验分析。上述引物均由武汉天一辉远生物科技有限公司设计和合成。
表2 长鳍吻不同免疫因子保守序列扩增所用引物Tab.2 Amplification primers of different cytokine gene in R.ventralis
表3 长鳍吻不同免疫因子基因定量PCR扩增所用引物Tab.3 Quantitative PCR amplification primers of different cytokine genes in R.ventralis
1.3 DNA提取与克隆
以所合成的cDNA为模板,以HSP70-S/HSP70-A、IgM-S/IgM-A、IL10-S/IL10-A、IL1b-S/IL1b-A和βactin-S/βactin-A为引物进行PCR扩增。20 μL PCR 反应体系:灭菌的双蒸水 6.0 μL、2× qPCR Mix (SYBR® Select Master Mix,ABI)10.0 μL、2.5 μmol/L基因正反引物各1.0 μL、cDNA 2.0 μL。PCR 反应条件:95 ℃预变性 10 min;95 ℃ 15 s,59 ℃/60 ℃ 1 min,40个循环,60 ℃延伸5 min。在60 ℃→95 ℃过程中,每升温0.3 ℃检测一次,形成溶解曲线。反应产物经1%琼脂糖凝胶电泳,经Gel DocTMXR+凝胶成像系统(Bio-rad,美国)分析图像。结果以相同起始量的β-actin 进行校正,并以4个特异基因和β-actin 电泳带相对吸光度的比值来表示。
1.4 数据分析
采用 IBM SPSS statistics 22 软件进行Student's t-test统计分析与显著性检验。
2 结果与分析
2.1 HSP70、IgM、IL-10和 IL-1b基因序列分析
通过优化PCR 扩增的条件,获得HSP70、IgM、IL-10和IL-1b基因的特异扩增产物,经回收和测序后得到长度分别为412 bp、463 bp、520 bp和217 bp的部分cDNA序列(图1),获得GenBank登陆号为KY696266- KY696269。
与GeneBank中已有的鱼类序列进行比较,本研究中长鳍吻HSP70、IgM、IL-10和 IL-1b基因序列与圆口铜鱼(CoreiusguichenotiKF471123.1)、草鱼(HM452137.1)、鲢(DQ058296.1)、圆筒吻(R.cylindricusKF724884.1)的同源性依次为99%、87%、96%和97%。
2.2 长鳍吻HSP70、IgM、IL-10和IL-1b基因组织表达差异
利用定量PCR方法检测得到HSP70、IgM、IL-10和 IL-1b基因在感染小瓜虫组和对照组长鳍吻皮肤和肠道组织的表达差异,并以上述4个基因与β-actin电泳带吸光度的比值绘制了他们在长鳍吻皮肤和肠道组织中表达的相对丰度(图2)。由图 2可知,在皮肤组织中,IgM和IL-10在感染小瓜虫鱼体中表达量显著高于对照鱼(P<0.05),HSP70和IL-1b的表达则没有差异;在肠道组织中,IgM、IL-10和 IL-1b在感染鱼体中表达量显著增高(P<0.05),HSP70的表达量在感染鱼体和对照鱼体中则没有差异。


3 讨论
HSP70作为一种应激蛋白,其家族分子在帮助机体细胞应对各种不利环境中起关键作用,它通常作为危险信号以提高机体适应热的能力及对抗多种压力刺激,进而帮助生物体迅速适应环境变化以抵御外界不良损害[9-11]。本研究中通过分析发现,HSP70不论是在皮肤组织还是肠道组织中,在感染小瓜虫和对照组长鳍吻中表达量都没有显著差异(图2)。说明本研究中因应激刺激而感染小瓜虫后的长鳍吻在皮肤和肠道组织中并没有诱导出HSP70蛋白的合成。赵建华在研究圆口铜鱼的急性操作胁迫和长途运输胁迫的应激反应中发现,HSP70在鱼鳍的表达量最低[12]。但林亚秋等[13]研究表明,热刺激后草鱼鱼鳍中HSP 70的表达量最高,这可能是因为鱼类种类不同,热应激因子
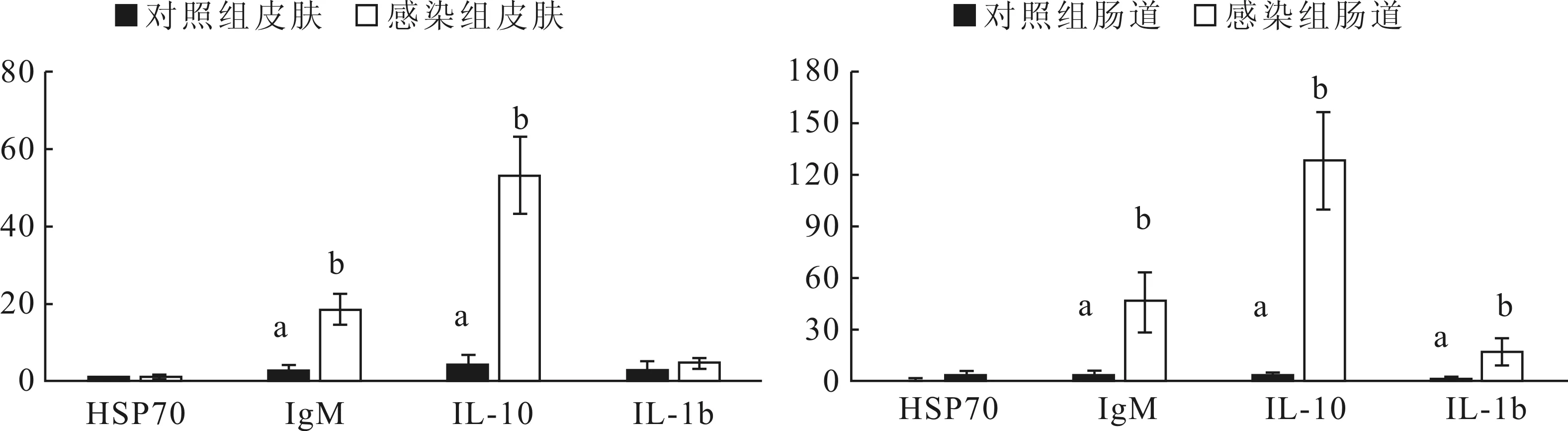
IgM作为鱼类的先天性免疫因子,是鱼体的天然抗体,存在于鱼类的血清、组织液、肠道、皮肤和鳃的粘液中[14]。本研究中,IgM在感染组长鳍吻中的皮肤和肠道中的表达量均显著提高(图2,P<0.05)。说明在长期吻的免疫应答系统中,抗体应答作用显著。
与哺乳类相比,鱼类的特异免疫系统尚不够发达,在鱼类对外界刺激及病原生物侵袭的防御反应中,非特异性免疫起着重要作用,鱼类非特异性免疫细胞和体液免疫因子包括溶菌酶、干扰素、补体、白细胞介素、铁传递蛋白、抗菌肽等[15]。IL-10是一种抑炎性细胞因子,主要功能是调节免疫和炎症反应,从而最大限度地减少因病原体或自身免疫系统对宿主的损害[16]。本研究中,IL-10在感染组长鳍吻的皮肤和肠道组织中均显著提高(图2,P<0.05),说明IL-10在长鳍吻免疫应答反应中较为敏感。相比较,IL-1b作为典型的促炎性细胞因子,仅在感染组长鳍吻的肠道组织中显著提高(图2,P<0.05),尽管有报道称小瓜虫感染的鲤鱼皮肤中IL-1b表达水平远高于血液中的表达,在36 h达到峰值[17]。上述结果表示在本研究中,相对于非特异性免疫因子IL-1b,IL-10在长鳍吻免疫应答反应中更敏感。
[1]管 敏,曲焕韬,胡美洪,等.长鳍吻人工繁育的初步研究[J].水产科学,2015,34(5):294-299.
[2]程晓凤.长江上游特有鱼长鳍吻(Rhinogobioventralis)遗传结构分析[D].重庆:西南大学,2013.
[3]刘 军.长江上游特有鱼类受威胁及优先保护顺序的定量分析[J].中国环境科学,2004,24(4):395-399.
[4]李继龙,王国伟,杨文波,等.国外渔业资源增殖放流状况及其对我国的启示[J].中国渔业经济,2009,27(3):111-123.
[5]邓辉胜,何学福.长江干流长鳍吻的生物学研究[J].西南农业大学学报(自然科学版),2005,27(5):704-708.
[6]辛建峰,杨宇峰,刘焕章.长江上游长鳍吻年龄与生长的研究[J].四川动物,2010,29(3):352-356.
[7]曲焕韬,刘 勇,鲁雪报,等.长江上游长鳍吻(Rhinogobioventralis)的个体繁殖力[J].水产学杂志,2016,29(4):17-22.
[8]杨德国,周剑光,吴国犀,等.长江大鳍鳆的人工繁殖[J].中国水产科学,1998,5(2):26-30.
[9]Lindquist A,Craig E A.The heat-shock proteins[J].Annu Rev Genet,1988,22(2):631-677.
[10]Yoshimi T,Hiroshige O Y.Induction profile of HSP70-cognate genes by environmental pollutants in Chironomidae[J].EnvironToxicol Pharmacol,2009,8(2):294-301.
[11]吴圣楠,隗黎丽,李海军,等.三角帆蚌热休克蛋白70基因克隆及其表达分析[J].水生生物学报,2017,41(1):50-55.
[12]赵建华.圆口铜鱼应激生理机制及缓解策略研究[D].武汉:华中农业大学,2014.
[13]林亚秋,郑玉才,吉 红,等.草鱼HSP70基因cDNA部分序列克隆及其组织表达差异[J].淡水渔业,2009,39(4):67-71.
[14]张永安,聂 品.鱼类体液免疫因子研究进展[J].水产学报,2000,24(4):376-380.
[15]李 莉,李春梅.鱼类非特异性免疫研究进展[J].河南农业科学,2012,41(2):26-32.
[16]代礼平.草鱼NCCRP_1和IL_10基因的克隆和表达[D].广东湛江:广东海洋大学,2011.
[17]Gonzalez S F,Buchmann K,Nielsen M E.Real-time gene expression analysis in carp (CyprinuscarpioL.)skin:inflammatory responses caused by the ectoparasiteIchthyophthiriusmultifiliis[J].Fish Shellfish Immunol,2007,22(6):641-650.
