miR-124靶向抑制NFATc1调控小鼠C3H10T1/2细胞软骨分化的研究
2018-01-24靳松黄爱军林昆彭建强李亚杰邹学农
靳松 黄爱军 林昆 彭建强 李亚杰 邹学农
骨髓间充质干细胞(MSC)是一类来源于骨髓的具有多向分化潜能的干细胞,具有自我更新、组织再生和抗炎修复等功能。研究表明,MSC经诱导分化后性别决定区Y框蛋白9(Sox9)、Ⅱ型胶原及缺氧诱导因子-1α(HIF-1α)的表达水平上调,提示其具有转化为类髓核细胞的潜能[1]。MSC移植修复椎间盘退变疾病的重要机制是,在椎间盘微环境中MSC向类髓核细胞分化并增加基质大分子的合成。MSC来源广泛、体外培养可诱导类髓核细胞分化,是一种非常有前景的可应用于临床治疗椎间盘退变疾病的种子细胞。微小RNA(miR)是一类含有18~24个核苷酸的非编码RNA,通过与靶基因mRNA 3’端非翻译区(3’UTR)的互补序列以碱基配对方式执行降解靶基因或抑制其翻译的功能,参与调控MSC分化过程中的多种生物学过程[2-4]。本研究通过在线软件预测miR-124的靶基因,发现活化T细胞核因子c1(NFATc1)是其中一个潜在靶点。NFAT家族是一类具有广泛生理功能的转录因子,在成软骨分化过程中,NFATc1能促进软骨基因表达[5]。NFATc1广泛参与MSC生长、分化的生物学过程,特别是在成软骨过程中有基因调控的功能[6]。由此推测,miR-124可能通过靶向调控NFATc1表达促进人MSC向软骨细胞分化。本研究采用成软骨分化诱导培养体系诱导C3H10T1/2细胞向成软骨细胞分化,并探究miR-124在此过程是否通过调控NFATc1靶基因发挥生物学作用。
材料与方法
一、主要材料
间充质干细胞株C3H10T1/2细胞系(中山大学骨科研究所提供);高糖型DMEM培养基、10%胎牛血清购自美国HyClone公司;谷氨酰胺购自美国Gibco公司;胰酶、0.02%乙二胺四乙酸(EDTA)、抗坏血酸、青霉素链霉素双抗、β-甘油磷酸钠、地塞米松购自美国Sigma公司; pmiR-RB-REPORTTM质粒载体、ribo FECTTMCP转染试剂盒购自广州锐博生物科技有限公司;LipofectamineTM2000转染试剂盒及Trizol购于美国Invitrogen公司。Spe I、Hind Ⅲ、T4 DNA连接酶和SYBR Premix ExTaqTM定量PCR试剂均为大连宝生物公司产品。实时定量PCR miR检测试剂盒购自广州天根生物公司。miR-124模拟物(mimics)及抑制物(inhibitors)为美国Ambion公司产品。定点突变试剂盒购自美国Stratagene公司。双荧光素酶活性检测试剂盒购自美国Promega公司。Smad4抗体和GAPDH抗体购自美国CST公司。NFATc1小干扰RNA(siRNA)由美国Invitorgen公司合成,引物合成及测序由深圳达生生物科技公司完成。
二、方 法
1.细胞培养与软骨诱导分化
以离心法进行软骨微团的构建,具体操作步骤参照文献[7]进行。小鼠C3H10T1/2细胞株经复苏后室温离心收集沉淀,用含10%胎牛血清的高糖型DMEM培养液混悬于37℃、5%CO2静置培养,待细胞融合至80%~90%,在胰酶消化下进行传代,每3 d更换培养液1次。将培养的C3H10T1/2细胞分为对照组与成软骨诱导组,分别加入含2%胎牛血清的DMEM和软骨诱导液诱导培养。每3 d换液1 次,于第0、7、10、14日收集细胞微球进行基因表达检测。
2.双荧光素酶报告基因构建及检测
使用在线靶基因预测软件(PicTar、TargetScan、miRanda、miRDB),预测NFATc1 mRNA的3’UTR上173~185 nt为miR-124的结合位点(GCACTTT)。人工合成NFATc1 mRNA 3’UTR 区的靶点野生型序列(Wt)以及定点突变的突变序列(Mut, GCACTTT→CGTGAAA); XhoⅠ及NotⅠ双酶切pmiR-RB-REPORTTM 质粒,再将Wt和Mut基因片段分别克隆到酶切后的pmiR-RBREPORTTM载体中,构建重组质粒pNFATc1-Wt或pNFATc1-Mut;基因测序确认重组质粒载体构建成功。按照试剂盒说明书突变NFATc1 3’UTR上miR-124结合位点,构建突变型pGL3-Mut-NFATc1真核表达载体。
取培养的C3H10T1/2细胞,根据转染质粒的不同分为4组:①pNFATc1-Wt质粒对照组,为转染pNFATc1-Wt质粒及miR-124 mimics对照;②pNFATc1-Wt质粒实验组,为转染pNFATc1-Wt质粒和miR-124 inhibitors;③pNFATc1-Mut质粒对照组,为转染pNFATc1-Mut质粒与miR-124 mimics对照;④pNFATc1-Mut质粒实验组,为转染pNFATc1-Mut质粒与miR-124 inhibitors。各组转染24 h后收集细胞,用双荧光素酶报告基因系统检测各组细胞的荧光素酶活性。每组实验重复3次,结果取平均值。另外,将NFATc1的3’UTR用XhoI/NotI点双酶切,插入到psiCHECKTM-2质粒中复制,与此相关的NFATc1 3’UTR片段被融合到荧光素酶中,并被转移到C3H10T1/2细胞,根据转染质粒分为不转染对照组(空白对照组)、miR-124 mimics组、miR-124 inhibitors组、pNFATc1-Wt质粒或pNFATc1-Mut组(正常对照组)、pNFATc1-Wt质粒或pNFATc1-Mut质粒加miR-124 inhibitors组(正常对照inhibitors组),分析miR-124对NFATc1荧光素酶活性的影响。
3.实时定量PCR检测
在成软骨诱导分化第0、7、10、14日收集C3H10T1/2细胞微球,按RNA提取试剂盒说明书提取总RNA,并检测总RNA的含量及纯度,再按照逆转录试剂盒说明书进行逆转录,合成模板DNA第一链,实时定量PCR扩增目的基因,引物序列见表1。反应条件为95℃10 min,95℃15 s,60℃ 30 s,72℃ 20 s,40个循环,内参为18 s。一步加A法逆转录miR,反应条件为95℃ 30 s,95℃ 3 s,60℃ 30 s,40个循环,内参为U6。上下游引物序列分别为5’-CTCGCTTCGGCAGCACA-3’和5’-AACGCTTCACGAATTTGCGT-3’,使用2-ΔΔCt法计算基因相对表达量。每组实验重复3次,结果取平均值。记录软骨分化的相关标志基因Aggrecan、Ⅱ型胶原蛋白(Col2a1)和Ⅹ型胶原蛋白(Col10a1)及Sox9 mRNA表达水平。另外,取成软骨诱导7 d的C3H10T1/2细胞转染pNFATc1-Wt质粒(Control组)、pNFATc1-Wt质粒及miR-124 mimics(miR-124组)、经siRNA干预的pNFATc1-Wt质粒(si-NFATc1组),按前述方法检测NFATc1 mRNA表达水平。此外,取成软骨诱导7 d的C3H10T1/2细胞,转染miR-124 mimics和(或)pNFATc1-Wt质粒,转染成功后分别加入miR-124 mimics 0、50、100 nmol/L miR-124 mimics或inhibitors培养24 h,用前述同样方法分析各组NFATc1和miR-124 mRNA表达水平。
4.蛋白免疫印迹法检测
提取各组细胞的总蛋白,测定蛋白浓度,加4倍SDS上样缓冲液8 μl和二硫苏糖醇2 μl混匀沸水浴5 min后作十二烷基硫酸钠聚丙烯酰胺SDS-PAGE凝胶电泳电泳,用电转印仪(100 V电压)电泳将凝胶上的蛋白转印至聚偏氟乙烯(PVDF)膜。将蛋白Marker剪下,避光保存,余下膜用5%脱脂牛奶室温封闭1 h,TBST缓冲液洗涤3次后,一抗4℃孵育过夜,TBST缓冲液再次洗涤3次,加入二抗室温孵育2 h,同前洗涤。按体积比1∶1混合ECL试剂盒中A液和B液,混合液铺在膜表面,凝胶成像系统曝光,Quantity one软件分析结果。

表1 PCR引物序列
三、统计学处理
结 果
一、C3H10T1/2软骨分化相关基因及蛋白表达的变化
成软骨诱导组中,在成软骨诱导分化第7、10、14日的软骨分化相关基因Aggrecan mRNA表达水平均逐渐升高(与诱导前比较,t分别为30.274、
46.132、160.752,P均<0.001);Col2a1 mRNA相对表达量在第7、10日升高(与诱导前比较,t分别为10.246、67.202、45.311,P均<0.001),第14日稍下降(与诱导前比较,t=53.244、P<0.001;与第10日比较,t=2.376、P>0.05);Col10a1第7、10日增加缓慢(与诱导前比较,P均>0.05),但第14日大幅上调(与诱导前比较,t=330.784,P<0.001)。软骨分化后, 软骨化基因表达逐渐稳步增加,但并未达到峰值;Sox 9和NFATc1 mRNA表达水平随着软骨分化进展而增加(与对照组0 d及10 d时比较,t分别为65.266、80.232,P均<0.001),见图1A~D。蛋白免疫印迹法显示,成软骨诱导组Col2a1、Sox 9和NFATc1蛋白呈逐渐增强表达,表明软骨发生的进展,而肥厚性标记Col10a1蛋白显示较稳定表达(图1E)。
二、miR-124对NFATc1的靶向性检测
通过Targetscan和miRanda等在线软件分析,发现NFATc1是miR-124的潜在靶基因之一, NFATc1 mRNA的3’UTR与miR-124的种子区存在理论上的互补碱基配对序列(图2A)。转染NFATc1 Wt质粒的对照组荧光素酶活性低于实验组(t=3.837,P<0.05);转染NFATc1 Mut质粒的实验组与对照组间荧光素酶活性比较差异无统计学意义(P>0.05);转染NFATc1 Wt质粒的实验组NFATc1蛋白表达水平高于对照组(图2B),表明miR-124直接作用于NFATc1。

图1 C3H10T1/2诱导成软骨分化的相关基因表达水平变化
miR-124组与si-NFATc1组的NFATc1 mRNA表达水平均低于Control组(LSD-t分别为5.642、4.338,P均<0.01),miR-124组与si-NFATc1组的NFATc1 mRNA表达水平接近(P>0.05);蛋白免疫印迹检测结果与实时定量PCR结果基本一致(图2C),表明miR-124能在C3H10T1/2软骨分化的过程中靶向调控NFATc1表达。

图2 miR-124 通过靶向NFATc1促进C3H10T1/2细胞软骨分化
A:互补碱基配对序列;B:NFATc1 荧光素酶活性检测结果和NFATc1蛋白表达;C:miR-124抑制NFAc1 mRNA和蛋白表达;与共转染NFATc1 Wt质粒的对照组荧光素酶活性比较,*P<0.01;与Control组比较,#P<0.01
三、抑制或过表达miR-124时C3H10T1/2的NFATc1表达水平变化
与未加入miR124 mimics相比,100 nmol/L的miR-124 mimics加入C3H10T1/2细胞后,miR-124 mRNA表达水平约增加60倍,而NFATc1 mRNA表达水平约降低60%(图3A)。当100 nmol/L miR-124 inhibitors加入C3H10T1/2细胞时,与未加入时相比,miR-124 mRNA表达水平减少了大约60%,而NFATc1增加了大约1倍(图3B)。转染NFATc1-Wt质粒的miR-124 mimics组NFATc1荧光素酶活性低于其他4组(F=8.256,LSD-t分别为5.436、7.442、3.433、6.282,P均<0.05),转染NFATc1-Mut质粒的miR-124 mimics组NFATc1荧光素酶活性与其他4组相近(P均>0.05),表明miR-124可作为NFATc1的抑制剂(图3C)。
讨 论
C3H10T1/2细胞系是一种小鼠胚胎源性间充质干细胞,具有多向分化潜能,转染效率高且稳定,易获得大量转染成功的活细胞,现已广泛用于研究MSC分化潜能的实验中[8]。miR作为一类重要的转录后调控因子,在MSC的分化过程中参与调控细胞的多种生物学过程[9-10]。已有研究发现,miR在调控MSC或前体细胞成软骨、成骨及成脂的分化过程中发挥重要生物学功能[11]。miR-124能抑制MSC向软骨分化[12]。上述研究证实,miR可通过抑制靶基因的表达参与MSC分化的调控。然而其中的分子机制和调控的靶基因尚不清楚。本课题组前期实验首先诱导的C3H10T1/2细胞向软骨的定向分化,在此过程中发现miR-124表达逐渐下调,提示其在MSC软骨分化中可能具有重要的调控作用;然后通过生物信息学分析预测出miR-124潜在的其中一个靶基因NFATc1。本研究亦显示,软骨分化后NFATc1和软骨化相关基因及蛋白表达逐渐稳步增加。
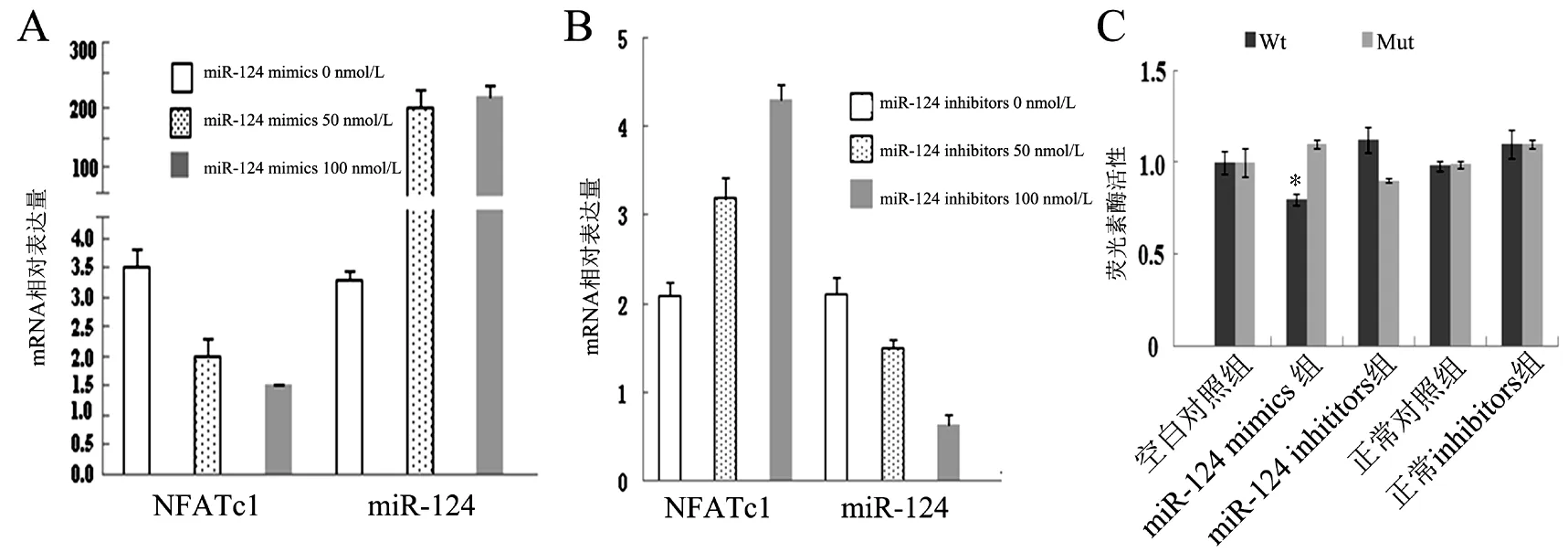
图3 干扰NFATc1或过表达miR-124时C3H10T1/2的NFATc1表达水平变化
与其他4组比较,*P<0.05
本研究将NFATc1 3’-UTR克隆入荧光素酶报告载体并且与miR-124共转染C3H10T1/2细胞,结果显示miR-124抑制NFATc1 mRNA的3’UTR荧光素酶活性,发现其特异性与NFATc1 mRNA的3’UTR相结合。然而在生理状态下,靶基因3’UTR序列中的miR结合位点有可能由于mRNA的构象而被掩盖,导致该miR无法发挥抑制靶基因表达的功能。因此进一步检测该miR与NFATc1 3’UTR的结合能力及其对NFAc1表达的抑制作用,结果表明miR-124能抑制软骨分化过程中的NFATc1 mRNA及蛋白表达,并且其强度与NFATc1 siRNA干扰NFATc1内源性表达一致,证明了miR-124直接靶向调控NFATc1表达。通过功能研究siRNA干扰NFATc1表达能抑制MSC的成软骨化,它能部分模拟miR-124调控MSC软骨分化的功能。另外,蛋白免疫印迹检测结果也显示转染miR-124 mimics后,NFATc1的蛋白相对表达量比对照组明显下调;相反,转染miR-124 inhibitor后NFATc1的蛋白相对表达量与对照组比较则上调,表明在C3H10T1/2细胞成骨分化过程中miR-124通过调控靶基因NFATc1介导MSC向软骨方向分化。
综上所述,本研究运用生物信息学方法对miR-124和NFATc1基因的靶向配对关系进行预测,发现两者靶向配对良好,采用荧光素酶报告系统、实时定量PCR及蛋白免疫印迹法发现miR-124可靶向抑制NFATc1的表达而负向调控C3H10T1/2细胞成软骨分化。鉴于NFATc1是钙形成信号通路中的重要转导因子之一,因此miR-124调控成软骨分化的作用机制可能是通过抑制NFATc1的表达,因此阻碍其下游信号通路的激活,进而抑制MSC的成软骨分化。
[1] Almalki SG, Agrawal DK. Key transcription factors in the differentiation of mesenchymal stem cells. Differentiation,2016,92(1-2):41-51.
[3] Zhang Z, Kang Y, Zhang Z, Zhang H, Duan X, Liu J, Li X, Liao W.Expression of microRNAs during chondrogenesis of human adipose-derived stem cells.Osteoarthritis Cartilage,2012,20(12):1638-1646.
[4] 田南南,杨建新,陈晓兰.miR-10b介导乳腺癌转移的机制与靶向治疗.新医学,2013,44(1):8-10.
[5] Lin SS, Tzeng BH, Lee KR, Smith RJ, Campbell KP, Chen CC.Cav3.2 T-type calcium channel is required for the NFAT-dependent Sox9 expression in tracheal cartilage.Proc Natl Acad Sci U S A,2014,111(19):E1990-E1998.
[6] Sitara D, Aliprantis AO.Transcriptional regulation of bone and joint remodeling by NFAT.Immunol Rev,2010,233(1):286-300.
[7] Ray P, Chapman SC.Cytoskeletal reorganization drives mesenchymal condensation and regulates downstream molecular signaling.PLoS One,2015,10(8):e0134702.
[8] Zou F, Xu JC, Wu GH, Zhou LL, Wa QD, Peng JQ, Zou XN.Effects of oncostatin M on cell proliferation and osteogenic differentiation in C3H10T1/2.J Musculoskelet Neuronal Interact,2016,16(4):377-385.
[9] Zhou X, Wang J, Sun H, Qi Y, Xu W, Luo D, Jin X, Li C, Chen W, Lin Z, Li F, Zhang R, Li G.MicroRNA-99a regulates early chondrogenic differentiation of rat mesenchymal stem cells by targeting the BMPR2 gene.Cell Tissue Res,2016,366(1):143-153.
[10] Hou C, Yang Z, Kang Y, Zhang Z, Fu M, He A, Zhang Z, Liao W.MiR-193b regulates early chondrogenesis by inhibiting the TGF-beta2 signaling pathway.FEBS Lett,2015,589(9):1040-1047.
[11] Laine SK, Alm JJ, Virtanen SP, Aro HT, Laitala-Leinonen TK.MicroRNAs miR-96, miR-124, and miR-199a regulate gene expression in human bone marrow-derived mesenchymal stem cells.J Cell Biochem,2012,113(8):2687-2695.
[12] Mondanizadeh M, Arefian E, Mosayebi G, Saidijam M, Khansarinejad B, Hashemi SM.MicroRNA-124 regulates neuronal differentiation of mesenchymal stem cells by targeting Sp1mRNA.J Cell Biochem,2015,116(6):943-953.
