黄淮海地区大豆主栽品种对8个大豆疫霉菌株的抗性评价
2018-01-22李晓那韩天富1
李晓那 孙 石 钟 超 韩天富1,,*
黄淮海地区大豆主栽品种对8个大豆疫霉菌株的抗性评价
李晓那1,2,**孙 石2,**钟 超2韩天富1,2,*
1东北农业大学农学院, 黑龙江哈尔滨 150030;2中国农业科学院作物科学研究所/ 农业部北京大豆生物学重点实验室, 北京 100081
随着麦茬免耕栽培技术的推广应用, 黄淮海地区麦后夏播大豆生产中疫霉根腐病呈加重趋势。了解该地区大豆主栽品种对疫霉根腐病的抗性和筛选抗病亲本, 对培育新的高产广适抗病品种具有重要意义。本研究利用8个具有不同毒力的大豆疫霉菌株, 采用下胚轴创伤接种法, 对20世纪50年代以来黄淮海地区审定、推广的140个大豆主栽品种进行接种鉴定。表明除6个品种对8个菌株均无抗性外, 其余134个品种分别抗1~8个大豆疫霉菌株, 占鉴定品种总数的95.7%, 其中抗6~8个以上菌株的品种有83个, 占鉴定品种总数的59.3%。以14个鉴别寄主的抗病反应型为参照, 发现134个品种对8个大豆疫霉菌株共产生65种反应型, 其中19个品种产生的5种反应型与已知单基因或2个单基因组合反应型相同; 115个品种产生的60种反应型与含有已知单基因或2个单基因组合的反应型不同, 推测可能含有新的抗病基因或基因组合。根据研究结果合理选择亲本, 可培育出聚合多个抗性基因且综合性状优良的大豆新品种。
大豆; 疫霉根腐病; 大豆疫霉; 抗性基因; 基因推导
近年来, 随着农村劳动力的转移和农村土地流转的加快, 黄淮海地区农业生产机械化水平逐步提高, 少耕免耕、秸秆还田等耕作栽培措施在生产上大面积推广应用[1], 这虽能起到保土、保水、增加土壤有机质的作用, 但也有利于病原菌的越冬、侵染和传播, 易造成作物根部病害流行和蔓延[2-3]。据国家大豆产业技术体系调查, 近年来黄淮海地区大豆根腐病加重发生, 其中, 危害最为严重的是大豆疫霉根腐病(Phytophthora root rot, PRR)[4]。该病是一种土传性病害, 可以在大豆的任何生育期侵染植株并造成危害。培育和种植抗病品种是防治该病害最经济、有效和环境友好的方法[5]。大豆对疫霉根腐病的完全抗性由单显性基因控制[6]。迄今, 国内外已在大豆的9条染色体(第2、第3、第7、第10、第13、第16、第17、第18、第19染色体)上定位了30个抗疫霉根腐病基因, 其中11个是由我国研究人员鉴定的[5,7-23]。大豆与大豆疫霉菌的互作为典型的基因对基因关系[24], 疫霉菌常因毒性基因的快速变异而导致高度的遗传分化, 特别是在抗病品种选择压力下, 新的毒力小种(毒力型)不断出现导致抗病品种抗性丧失[25-26]。因此, 需要不断培育和利用新的抗病品种。大面积种植品种(以下称为“主栽品种”)由于产量高、综合性状好、适应性强, 常被育种家作为选育抗病品种的亲本, 因此, 解析主栽品种对疫霉根腐病的抗性对培育新的抗病品种具有重要意义。本研究对黄淮海地区20世纪50年代以来审定、推广的140个大豆主栽品种进行鉴定, 比较不同年代和不同省份主栽品种对大豆疫霉的抗性表现, 了解其抗性多样性, 初步推导它们所蕴含的抗病基因, 为黄淮海地区大豆抗病品种的选育和疫霉根腐病的防治提供依据。
1 材料与方法
1.1 供试材料
140份材料来自黄淮海地区7省市(表1), 为20世纪50年代以来在该地区种植面积较大的大豆品种。此外, 选用14个含有已知抗大豆疫霉根腐病基因(基因)的大豆品种(系)及感病对照品种Williams作为抗病基因的鉴别品种(表2)。
表1 不同年代黄淮海地区主栽大豆品种
(续表1)
品种Cultivar来源Source审定年份Year of release品种Cultivar来源Source审定年份Year of release 五河大豆Wuhedadou安徽Anhui1977汾豆60 Fendou 60山西Shanxi2007 文丰5号Wenfeng 5山东Shandong1982邯豆7号Handou 7河北 Hebei2007 文丰7号Wenfeng 7山东Shandong1982石豆1号Shidou 1河北 Hebei2007 跃进5号Yuejin 5山东Shandong1982菏豆15 Hedou 15山东 Shandong2007 诱变30 Youbian 30北京Beijing1983冀豆18 Jidou 18河北Hebei2007 齐黄10 Qihuang 10山东Shandong1983石豆2号Shidou 2河北 Hebei2008 鲁豆3号Ludou 3山东Shandong1983淮豆9号Huaidou 9江苏 Jiangsu2008 鲁豆4号Ludou 4山东Shandong1985冀豆20 Jidou 20河北 Hebei2008 豫豆2号Yudou 2河南Henan1985皖豆28 Wandou 28安徽 Anhui2008 鲁豆5号Ludou 5山东Shandong1987山宁17 Shanning 17山东 Shandong2009 鲁豆6号Ludou 6山东Shandong1987徐豆16 Xudou 16江苏Jiangsu2009 豫豆8号Yudou 8河南Henan1988商豆6号Shangdou 6河南 Henan2009 中豆19 Zhongdou 19河南Henan1988石豆4号Shidou 4河北 Hebei2009 科丰6号Kefeng 6北京Beijing1989山宁16 Shanning 16山东Shandong2009 齐黄20 Qihuang 20山东Shandong1990商951099 Shang 951099河南Henan2009 菏84-5 He 84-5山东Shandong1990合豆4号Hedou 4安徽Anhui2009 皖豆12 Wandou 12安徽Anhui1991周豆19 Zhoudou 19河南Henan2010 冀豆7号Jidou 7河北Hebei1992菏豆19 Hedou 19山东Shandong2010 鲁豆10 Ludou 10山东Shandong1993菏豆20 Hedou 20山东Shandong2010 晋豆19 Jindou 19山西Shanxi1994邯豆8号Handou 8河北Hebei2010 中品661 Zhongpin 661北京Beijing1994冀豆21 Jidou 21河北Hebei2010 鲁豆11 Ludou 11山东Shandong1995皖豆29 Wandou 29安徽 Anhui2010 徐豆8号Xudou 8江苏Jiangsu1996中黄39 Zhonghuang 39北京Beijing2010 鲁豆12 Ludou 12山东Shandong1996汾豆79 Fendou 79山西Shanxi2011 豫豆22 Yudou 22河南Henan1997沧豆10 Cangdou 10河北Hebei2011 晋豆21 Jindou 21山西Shanxi1997石豆6号Shidou 6河北Hebei2011 豫豆25 Yudou 25河南Henan1998淮豆11 Huaidou 11江苏Jiangsu2011 徐豆9号Xudou 9江苏Jiangsu1998邯豆9号Handou 9河北Hebei2011 邯豆3号Handou 3河北Hebei1999邯豆10 Handou 10河北Hebei2011 化诱542 Huayou 542河北Hebei1999冀豆19 Jidou 19河北Hebei2011 晋豆23 Jindou 23山西Shanxi1999商豆14 Shangdou 14河南Henan2011 晋豆25 Jindou 25山西Shanxi2000中黄74 Zhonghuang 74北京Beijing2011 合豆1号Hedou 1安徽Anhui2000冀豆22 Jidou 22河北 Hebei2012 五星1号Wuxing 1河北Hebei2001郑7051 Zheng 7051河南Henan2012 中黄13 Zhonghuang 13北京Beijing2001中黄66 Zhonghuang 66北京Beijing2012 科丰53 Kefeng 53北京Beijing2001齐黄34 Qihuang 34山东Shandong2012 郑92116 Zheng 92116河南Henan2001齐黄35 Qihuang 35山东Shandong2012 冀豆12 Jidou 12河北Hebei2001皖宿2156 Wansu 2156安徽Anhui2013 菏豆12 Hedou 12山东Shandong2002皖豆33 Wandou 33安徽Anhui2013 郑9007 Zheng 9007河南Henan2003中黄70 Zhonghuang 70北京Beijing2013 郑交107 Zhengjiao 107河南Henan2003石豆7号Shidou 7河北Hebei2013 皖豆24 Wandou 24安徽Anhui2003石豆8号Shidou 8河北Hebei2014 周豆11 Zhoudou 11河南Henan2003徐豆20 Xudou 20江苏Jiangsu2014 合豆2号Hedou 2安徽Anhui2003中黄75 Zhonghuang 75北京Beijing2014 徐豆12 Xudou 12江苏Jiangsu2003中黄79 Zhonghuang 79北京Beijing2015 邯豆5号Handou 5河北Hebei2004皖豆35 Wandou 35安徽Anhui2015 晋豆28 Jindou 28山西Shanxi2004中黄76 Zhonghuang 76北京Beijing2016
1.2 供试菌株
8个具有不同毒力的大豆疫霉菌株分别为PsUSAR2 (毒力型: 1b、2、3c、5)、Ps41-1 (毒力型: 1a、1b、1d、2、3a、3b、3c、4、5、6、7、8)、PsMC1 (毒力型: 1b、1c、1d、1k、2、3a、3b、3c、4、5、7)、PsRace1 (毒力型: 1a)、PsRace3 (毒力型: 1a、1d)、PsRace4 (毒力型: 1a、1d、3b)、PsRace5 (毒力型: 1a、7)和PsJS2 (毒力型: 1a、1b、1c、1d、1k、2、3a、3b、3c、4、5、6、7、8)。疫霉菌株由单个孢子纯化后保存在V8汁琼脂培养基上, 接种鉴定前将菌株转入含1.5%琼脂的胡萝卜(CA)培养基培养皿中, 在25℃黑暗条件下培养7~10 d, 接种到Williams幼苗以恢复毒力, 植株发病后再进行病原菌分离和纯化, 接种一套寄主矫正毒力, 保证接种的准确性。
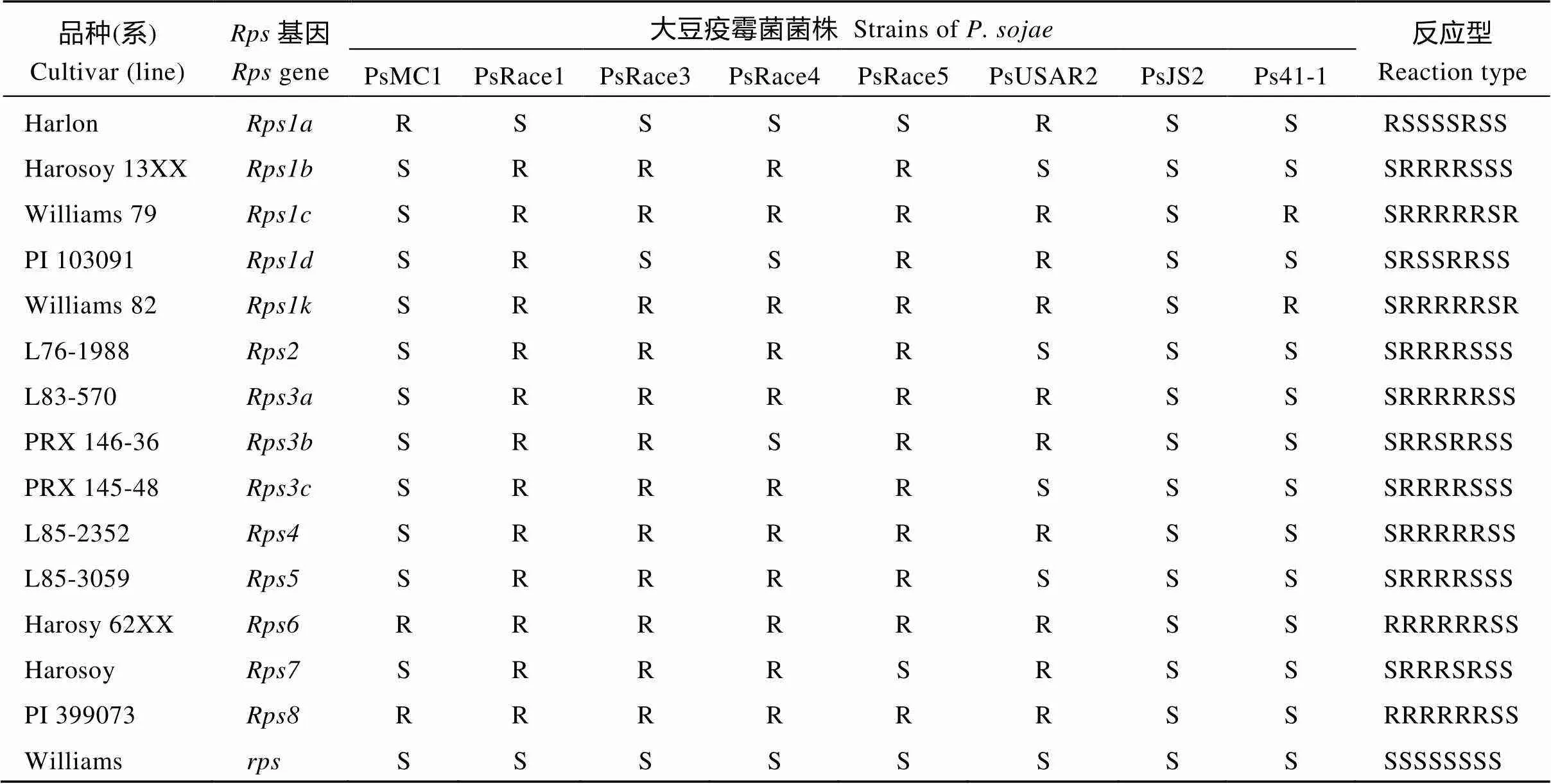
表2 含已知抗病基因的大豆品种(系)对8个大豆疫霉菌株的抗性反应
R: 抗病; S: 感病。R: resistant; S: susceptible.
1.3 抗性鉴定
将140个供试大豆品种、14个鉴别寄主和感病品种Williams分别取15粒播种于以蛭石为基质、直径为10 cm的塑料花盆中, 出苗期温度控制在(25~29)°C, 出苗后保留10株长势一致的健康植株, 待第一对真叶(单叶)完全展开时(出土后7~10 d), 采用下胚轴创伤接种法接种[27]。用10 mL注射器上的针尖在大豆幼苗子叶下约0.5 cm处向下划开1 cm长的伤口, 深度不能超过茎粗的1/3, 然后将混匀的菌丝体注于伤口处。接种后在(18~25)℃条件下保湿48 h, 然后转入相同温度的温室继续培养。接种5~6 d后参照朱振东等[28]的抗性评价标准进行病情调查。植株死亡≥70%视为感病(susceptible, S),<30%则为抗病(resistant, R), 在30%~70%之间为中间类型(intermediate, I)。试验过程中进行3次重复鉴定。
1.4 抗病基因推导
将中间反应类型归为抗病反应[29], 比较待鉴定品种与14个鉴别寄主对8个大豆疫霉菌株的反应型。若待鉴定品种的反应型与某个鉴别寄主的反应型或某2个鉴别寄主组合的反应型一致, 则说明产生这种反应型的大豆品种很有可能含有和该鉴别寄主相同的抗病基因或2个鉴别寄主携带的抗病基因组合; 若待鉴定品种的反应型完全不同于14个鉴别寄主所含抗病基因或基因组合的反应型, 则这些品种可能携带未知的新基因或基因组合。
2 结果与分析
2.1 黄淮海地区不同年代大豆主栽品种对疫霉病的抗性表现
在黄淮海地区的140个大豆主栽品种中, 共有134个对1~8个菌株表现出抗性, 占鉴定总数的95.7%; 6个对8个菌株均无抗性。对不同菌株表现抗性的品种比例见图1, 其中, 对PsRace5菌株表现抗性的比例最高, 为82.1%; 抗PsRace1菌株的品种最少, 为54.3%。对不同数目疫霉菌株表现抗性的抗病品种所占比例见图2, 其中26个品种抗8个菌株, 占鉴定总数的18.6%; 22个品种抗7个菌株, 占15.7%; 35个品种抗6个菌株, 占25.0%。
黄淮海地区不同年代大豆主栽品种对疫霉菌不同菌株抗性的比例有一定变化。图3显示, 20世纪80年代以后审定的品种对PsMC1菌株的抗性比例相对较高; 20世纪80年代审定的主栽品种对PsRace1全部表现为抗病; 1980—2009年审定的主栽品种对PsJS2菌株抗病比例最高; 1970—1999年审定的主栽品种对PsRace3、PsRace4和PsRace5的抗性比例较高, 均达到80.0%以上。从主栽品种对疫霉菌抗性的变化趋势可以看出, 尽管不同年代大豆主栽品种抗性比例有差别, 但每个年代均存在抗大豆疫霉根腐病的品种。抗6个以上菌株的品种共83个, 占鉴定品种总数的59.3%, 其在不同年代抗病品种中的分布比例如图4。可以看出, 20世纪70年代多抗品种比例相对较低, 随后不断增多, 在2000—2009年间多抗品种比例高, 2010年后则有所下降。
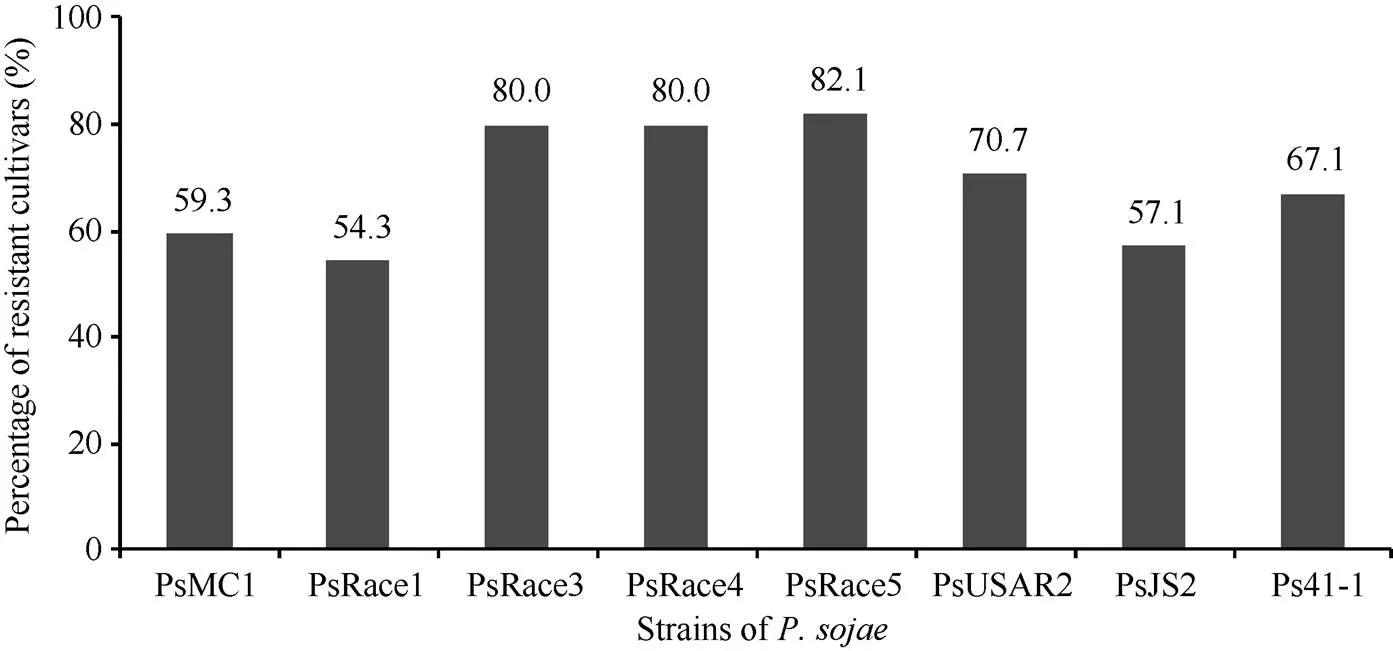
图1 抗不同大豆疫霉菌株的品种数量分布比例
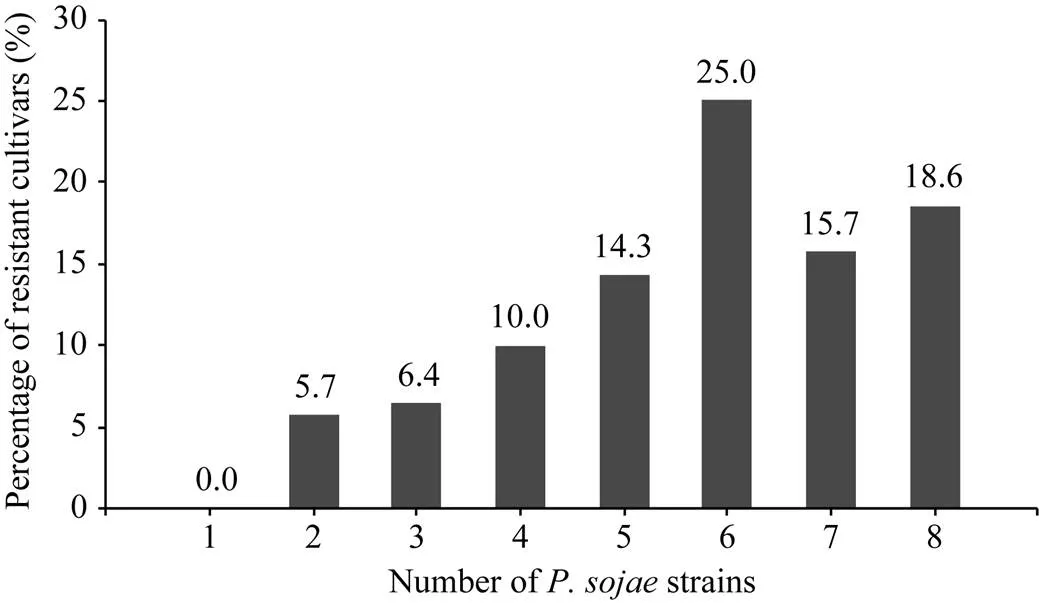
图2 供试大豆品种抗大豆疫霉菌株数目的分布
2.2 黄淮海地区不同省份大豆主栽品种对疫霉病的抗性表现
在所鉴定的大豆品种中, 来源于山东的品种所含抗病反应类型最丰富, 其次为来自河南、河北、北京、安徽、江苏和山西的品种。供试33个山东大豆品种表现出24种不同的反应型, 占总反应型(66种)的36.4%; 30个河南大豆品种表现出20种不同的反应型, 占30.3%; 25个河北大豆品种有18种不同的反应型, 占27.3%; 14个北京大豆品种有12种不同的反应型, 占18.2%; 16个安徽大豆品种有11种不同的反应型, 占16.7%; 11个江苏大豆品种有8种不同的反应型, 占12.1%; 11个山西大豆品种有8种不同的反应型, 占12.1% (图5)。对不同抗病反应型在各省(市)的分布情况比较后发现, 反应型为RRRRRRRR的大豆品种在山东、江苏、河南、安徽、山西和河北均有存在, 但部分反应型仅出现在某一个省份, 如RSRSSSSS反应型仅存在于山东大豆品种中; RRRRRSSR反应型仅存在于北京大豆品种中; SSRRSRSS反应型仅存在于河南大豆品种中; SSRRSRRR反应型仅存在于河北大豆品种中; RSRSRRRR反应型仅存在于山西大豆品种中; SSRRRRRR反应型仅存在于安徽大豆品种中; RRRRRSRR反应型仅存在于江苏的大豆品种中。由此可见, 黄淮海地区不同省(市)的大豆品种对疫霉菌的抗性水平存在明显差异, 在大豆抗病育种中, 应重视选用不同来源的亲本。由于本研究中所利用的品种数量有限, 对各省(市)品种抗病反应型的分析仍需要进一步研究。
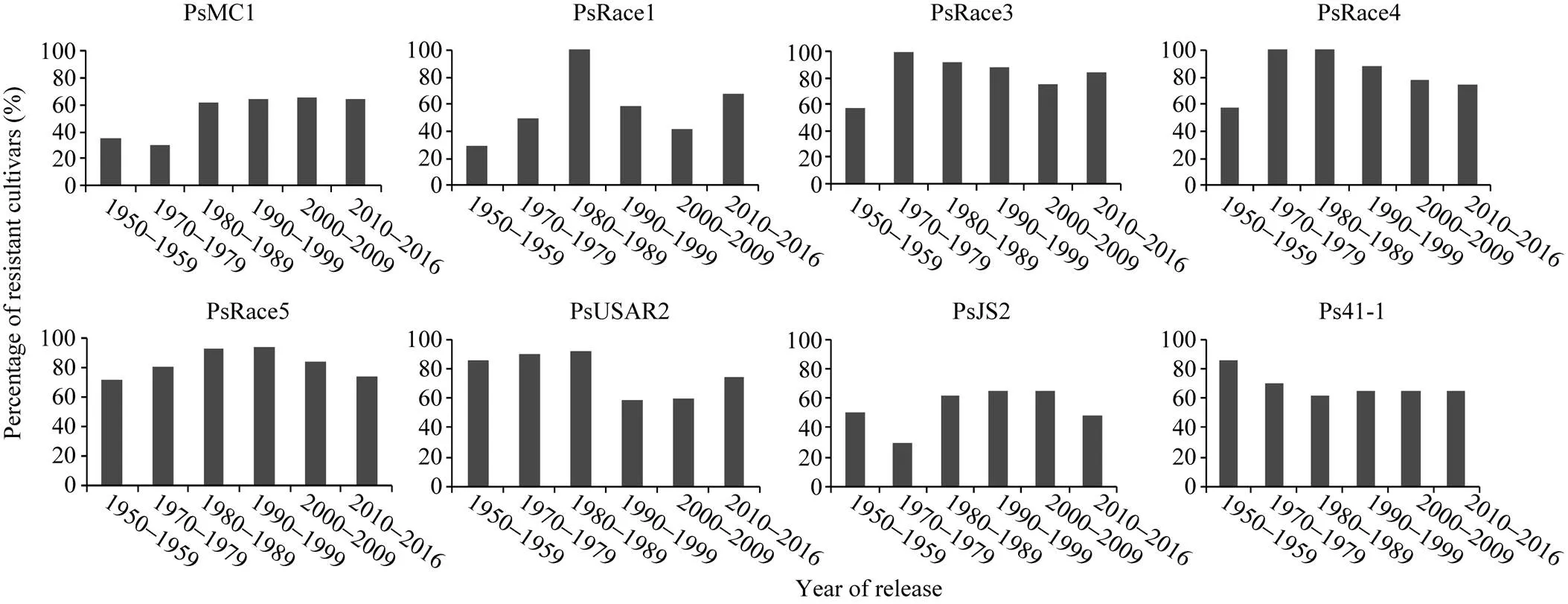
图3 不同年代大豆主栽品种对8个疫霉菌株的抗性比例
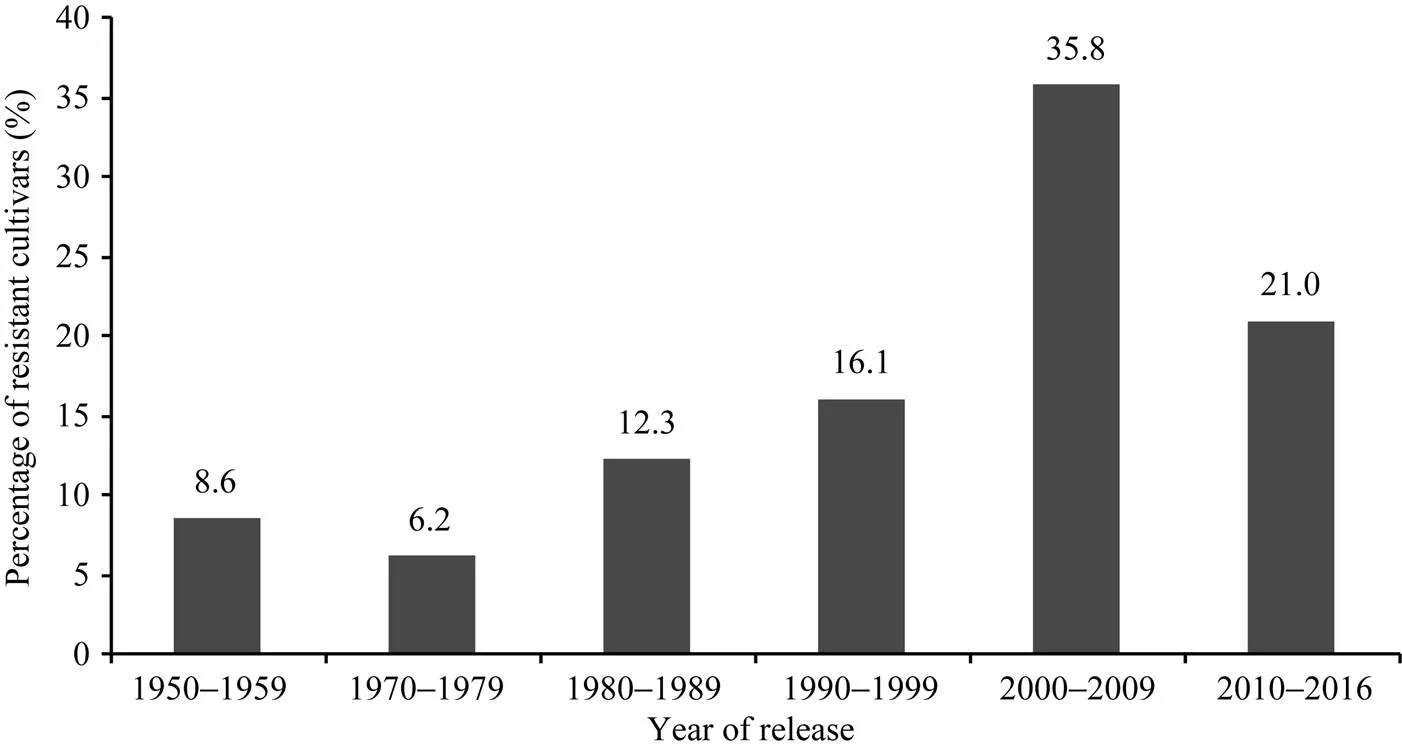
图4 抗6个以上菌株的品种在不同年代的分布比例
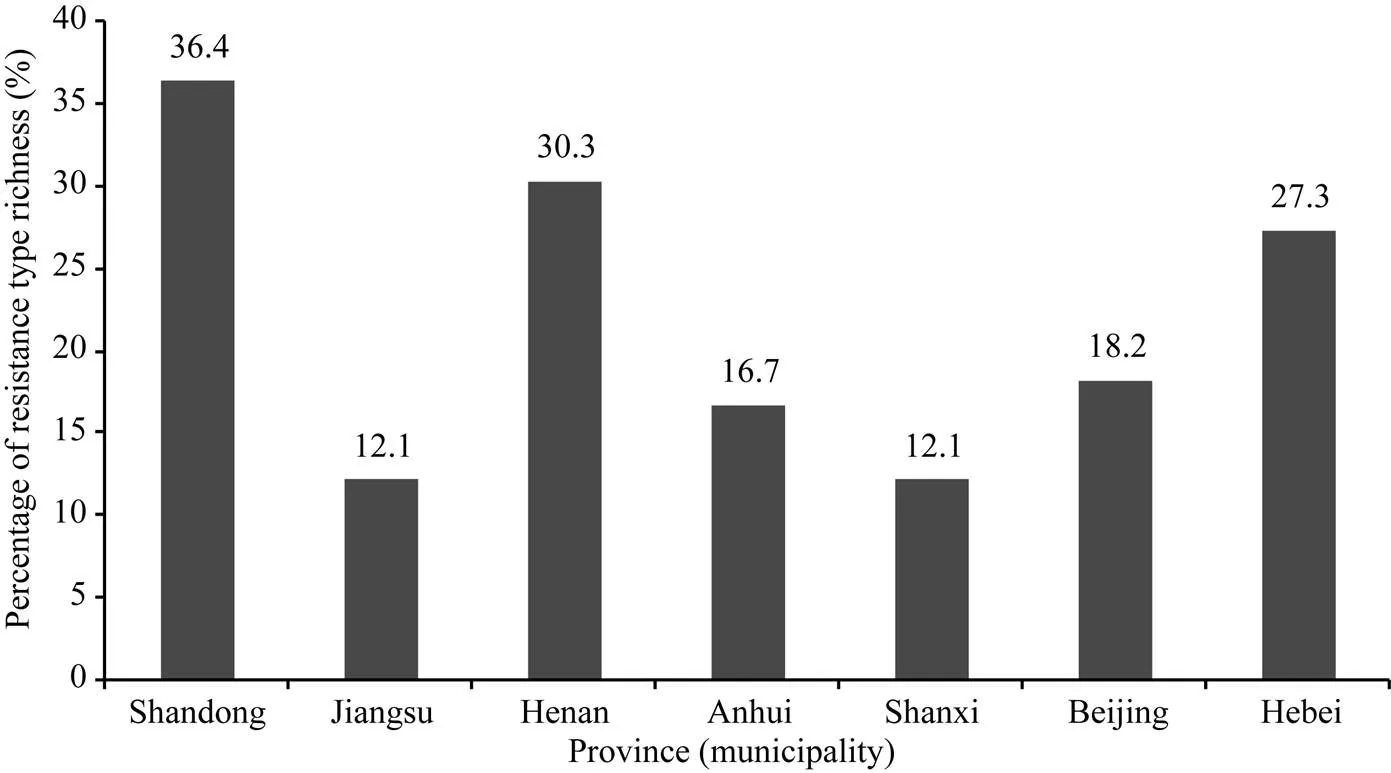
图5 7省(市)抗大豆疫霉根腐病的大豆品种分布情况
2.3 黄淮海地区大豆主栽品种抗病基因型的推导
14个分别抗不同大豆疫霉菌株的大豆品种及对照感病品种Williams对8个菌株的反应型见表2。在140个主栽品种中, 郑120、邳县软条枝、徐豆12、中黄70、中黄79和邯豆5号对8个疫霉菌株的反应型表现为SSSSSSSS, 与感病对照品种Williams ()的反应型一致, 说明这些品种不含已知抗疫霉根腐病基因。有134个品种分别抗1~8个菌株, 共产生65种不同的反应型, 其中有19个品种产生的5种反应型符合已知单基因、2个单基因组合的反应型, 115个品种产生的60种反应型与含有已知单基因或2个单基因组合的反应型不同, 说明这些品种可能含有新的抗病基因或基因组合, 其中大豆疫霉菌株PsJS2能够击败所有已知抗病基因, 抗PsJS2的品种应该含有新抗病基因。
从不同年代大豆主栽品种抗病基因型的推导结果可知, 20世纪50年代审定的14个品种共产生12种反应型, 可能含有4个抗病基因、或; 1970—1979年审定的10个品种共产生10种反应型, 可能含有4个抗病基因、、或; 20世纪80年代审定的13个大豆品种共产生9种反应型, 可能含有6个抗病基因、、、、或; 20世纪90年代审定的17个品种共产生15种反应, 可能含有8个抗病基因、、、、、、或; 2000—2010年审定的55个品种共产生36种反应型, 推断可能含有6个抗病基因、、、、或; 2010—2016年审定的31个品种共产生17种反应型, 推断可能含有6个抗病基因、、、、或, 由此可知, 每个年代均有含抗病基因或的材料。从黄淮海地区7省(市)大豆品种所含抗病基因类型(表3)可知, 山东和河南大豆主栽品种含有已知抗病基因类型最多, 其次为江苏、河北、山西和北京的品种。
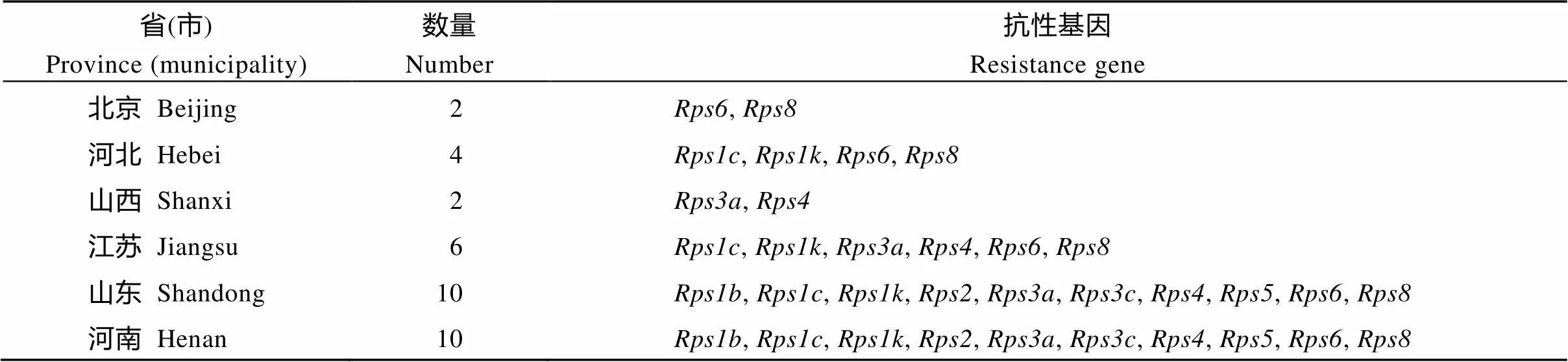
表3 7省(市)大豆主栽品种所含已知抗性基因的数量和类型
3 讨论
3.1 黄淮海地区大豆主栽品种对大豆疫霉根腐病抗性的多样性
我国是大豆起源地, 拥有丰富的大豆种质资源。国内外研究学者已对我国大豆种质进行了疫霉根腐病抗性鉴定。孙石等[30]利用7个具有不同毒力公式的大豆疫霉菌株, 对黄淮海大豆主产区的96个大豆品种(系)进行接种鉴定, 发现黄淮海地区大豆品种对大豆疫霉根腐病具有丰富的抗性多样性; Lohnes等[31]用4个不同毒力型的生理小种接种726份大豆资源, 发现安徽、江苏、山东等地大豆品种抗疫霉根腐病的比例较高; 朱振东等[32]利用10个具有不同毒力型的大豆疫霉菌株对120个栽培大豆品种(系)进行抗性鉴定, 发现河南、安徽和山西等地大豆品种(系)抗性资源丰富。任海龙等[33]利用大豆疫霉菌株Pm14接种鉴定江西、湖南和广西273份野生大豆资源的抗病性, 发现54份抗病或中间类型的资源, 占供试材料的19.8%; 程艳波等[34]利用7个具有不同毒力型的疫霉菌株接种鉴定华南地区67个大豆品种(系)及其亲本的抗病性, 发现抗疫霉菌株PGD1的比例最高, 占鉴定品种总数的9.0%; 申宏波等[35]利用大豆疫霉菌1号生理小种对黑龙江省346份大豆种质资源进行抗性鉴定显示, 表现为抗病和中间类型的品种有27个, 仅占供试材料总数的7.8%。本研究利用8个不同毒力型的疫霉菌株接种黄淮海地区自20世纪50年代以来审定的140个大豆主栽品种, 发现其中95.7%品种表现抗性, 结果再次证明黄淮海地区大豆品种对抗疫霉根腐病存在丰富的抗性多样性。
3.2 抗病基因的推导
基因推导是分析寄主抗病基因的一种简单、快速的方法。本研究通过与14个鉴别寄主品种对8个鉴别菌株的反应型相比较, 发现供试品种在对疫霉根腐病的抗性方面表现出66种不同的反应型, 其中有5种反应型符合已知单基因、2个单基因组合的反应型, 相关主栽品种可能含有8个抗疫霉根腐病基因、、、、、、和。在我国已定位和鉴定的11个抗病基因中就有5个是从黄淮海大豆主栽品种中发现的, 其中、、、和分别来源于豫豆25、鲁豆4号、皖豆15、早熟18和诱变30[10-15,17]。本研究对豫豆25、鲁豆4号和诱变30的抗病基因推导结果表明, 这3个品种可能含有新的抗病基因。基因推导方法虽然具有一定的局限性[36], 但是可利用此方法推导出品种可能含有的抗病基因, 然后进一步应用常规遗传学的方法和分子生物学技术进行基因鉴定和定位。事实也证明此种策略有效, 我国已定位和鉴定的11个大豆疫霉抗病基因都是采取这种策略获得的。所以基因推导法在大豆抗病材料筛选和和抗病育种中仍有重要价值。
3.3 主栽品种在大豆抗病育种上的应用
根据不同地区疫霉菌株基因型有针对性地选育和利用抗病品种是防控疫霉病害的有效措施。在本研究所用的8个疫霉菌株中, PsMC1和PsJS2是从黄淮海地区分离到的, 主栽品种对这2个菌株的抗性频率分别为59.3%和57.1%, 而14个鉴别品种对PsJS2均感病。PsRace3是分离自东北地区的大豆疫霉菌株, 河南省地区大豆品种抗PsRace3的比例高达90.0%。唐庆华等[37]对来自全国8省(市)的115个大豆疫霉分离物进行毒力测定发现, 含有抗病基因、和的鉴别寄主可以抵抗90.0%以上大豆疫霉分离物的侵染, 因此, 提出在中国种植含有、和的大豆品种将会有效地降低疫霉根腐病的发生。在本研究中, 推导出8个品种可能含有或, 其中兖黄1号、郑州135、文丰7号、豫豆8号、郑9007五个品种同时抗6个大豆疫霉菌株, 而海阳爬蔓青、徐豆9号和邯豆8号3个品种同时抗7个疫霉菌株, 可以利用这些品种和产量高、综合性状好的主栽品种配制杂交组合, 也可以利用这些品种与含有其他抗性基因的品种杂交, 进一步聚合多个抗性基因, 培育高产、多抗的大豆品种。
4 结论
在黄淮海地区不同年代审定的140个大豆主栽品种中, 6个品种对8个大豆疫霉菌株均无抗性, 134个品种分别抗1~8个大豆疫霉菌株。19个品种产生5种反应型, 可能含有单基因或2个单基因组合, 其中8个品种可能含有抗病基因或, 这些品种在大豆抗疫霉根腐病育种中具有重大价值, 是培育多抗品种的基因资源。
致谢:中国农业科学院作物科学研究所朱振东研究员提供大豆疫霉菌菌株, 并在试验和论文撰写中提供指导, 谨致谢意。
[1] 王幸, 吴存祥, 齐玉军, 徐泽俊, 王宗标, 韩天富. 麦秸处理和播种方式对夏大豆农艺性状及土壤物理性状的影响. 中国农业科学, 2016, 49: 1453–1465 Wang X, Wu C X, Qi Y J, Xu Z J, Wang Z B, Han T F. Effects of straw management and sowing methods on soybean agronomic traits and soil physical properties., 2016, 49: 1453–1465 (in Chinese with English abstract)
[2] 罗永藩. 我国少耕与免耕技术推广应用情况与发展前景. 耕作与栽培, 1991, (2): 1–7 Luo Y F. The popularization and application of less tillage and no tillage technology in China., 1991, (2): 1–7 (in Chinese)
[3] 田慧, 谭周进, 屠乃美, 李建国, 肖启明. 少免耕土壤生态学效应研究进展. 耕作与栽培, 2006, (5): 10–12 Tian H, Tan Z J, Tu N M, Li J G, Xiao Q M. Research progress on soil ecological effects of no tillage., 2006, (5): 10–12 (in Chinese)
[4] 朱振东, 王晓鸣, 戴法超. 大豆疫霉根腐病在我国的发生及防治对策. 植物保护, 1999, 25(3): 47–49 Zhu Z D, Wang X M, Dai F C. Occurrence and control of phytophthora root rot of soybean in China., 1999, 25(3): 47–49 (in Chinese)
[5] Sun J T, Li L H, Zhao J M, Huang J, Yan Q, Xing H, Guo N. Genetic analysis and fine mapping of, a novel resistance gene toin soybean [(L.) Merr.]., 2014, 127: 913–919
[6] Sugimoto T, Kato M, Yoshida S, Matsumoto I, Kobayashi T, Kaga A, Hajika M, Yamamoto R, Watanabe K, Aino M, Matoh T, Walker D R, Biggs A R, Ishimoto M. Pathogenic diversity ofand breeding strategies to develop-resistant soybeans., 2012, 61: 511–522
[7] Weng C, Yu K, Anderson T R, Poysa V. Mapping genes conferring resistance to phytophthora root rot of soybean,and., 2001, 92: 442–446
[8] Burnham K D, Dorrance A E, Francis D M, Fioritto R J, Martin S K St., a new locus in soybean for resistance to., 2003, 43: 101–105
[9] Gordon S G, Martin S K St, Dorrance A E.maps to a resistance gene rich region on soybean molecular linkage group F., 2006, 46: 168–173
[10] Wu X L, Zhang B Q, Sun S, Zhao J M, Yang F, Guo N, Gai J Y, Xing H. Identification, genetic analysis and mapping of resistance toof Pm28 in soybean., 2011, 10: 1506–1511
[11] Zhang J Q, Xia C J, Duan C X, Sun S L, Wang X M, Wu X F, Zhu Z D. Identification and candidate gene analysis of a novelresistance genein a Chinese soybean cultivar., 2013, 8: e69799
[12] 朱振东, 霍云龙, 王晓鸣, 黄俊斌, 武小菲. 一个抗大豆疫霉根腐病新基因的分子鉴定. 作物学报, 2007, 33: 154–157 Zhu Z D, Huo Y L, Wang X M, Huang J B, Wu X F. Molecular identification of a novelresistance gene in soybean., 2007, 33: 154–157 (in Chinese with English abstract)
[13] Sun S, Wu X L, Zhao J M, Wang Y C, Tang Q H, Yu D Y, Gai J Y, Xing H. Characterization and mapping of, a novel resistance gene to., 2011, 130: 139–143
[14] 范爱颖, 王晓鸣, 方小平, 武小菲, 朱振东. 大豆品种豫豆25抗疫霉根腐病基因的鉴定. 作物学报, 2009, 35: 1844–1850 Fan A Y, Wang X M, Fang X P, Wu X F, Zhu Z D. Molecular identification ofresistance gene in soybean cultivar Yudou25., 2009, 35: 1844–1850 (in Chinese with English abstract)
[15] 姚海燕, 王晓鸣, 武小菲, 肖炎农, 朱振东. 大豆品种早熟18抗疫霉根腐病基因的SSR分子标记. 植物遗传资源学报, 2010, 11: 213–217 Yao H Y, Wang X M, Wu X F, Xiao Y N, Zhu Z D. Molecular mapping ofresistance gene in soybean cultivar Zaoshu 18., 2010, 11: 213–217 (in Chinese with English abstract)
[16] 于安亮, 徐鹏飞, 王金生, 张淑珍, 吴俊江, 李文滨, 陈维元, 李宁辉, 范素杰, 王欣, 姜良宇. 大豆品种绥农10抗疫霉根腐病遗传分析及抗病基因的SSR标记. 中国油料作物学报, 2010, 32: 462–466 Yu A L, Xu P F, Wang J S, Zhang S Z, Wu J J, Li W B, Chen W Y, Li N H, Fan S J, Wang X, Jiang L Y. Genetic analysis and SSR mapping of gene resistance torace 1 in soybean cv Suinong 10., 2010, 32: 462–466 (in Chinese with English abstract)
[17] 武晓玲, 周斌, 孙石, 赵晋铭, 陈受宜, 盖钧镒, 邢邯. 大豆对大豆疫霉菌株Pm14抗性的遗传分析及基因定位. 中国农业科学, 2011, 44: 456–460 Wu X L, Zhou B, Sun S, Zhao J M, Chen S Y, Gai J Y, Xing H. Genetic analysis and mapping of resistance toofin soybean., 2011, 44: 456–460 (in Chinese with English abstract)
[18] Zhang J Q, Xia C J, Wang X M, Duan C X, Sun S L, Wu X F, Zhu Z D. Genetic characterization and fine mapping of the novelresistance gene in a Chinese soybean cultivar., 2013, 126: 1555–1561
[19] Lin F, Zhao M X, Ping J Q, Johnson A, Zhang B, Abney T S, Hughes T J, Ma J X. Molecular mapping of two genes conferring resistance toin a soybean landrace PI 567139B., 2013, 126: 2177–2185
[20] Ping J Q, Fitzgerald J C, Zhang C B, Lin F, Bai Y H, Wang D C, Aggarwal R, Rehman M, Crasta O, Ma J X. Identification and molecular mapping of, a novel gene conferring resistance toin soybean., 2016, 129: 445–451
[21] Sahoo D K, Abeysekara N S, Cianzio S R, Robertson A E, Bhattacharyya M K. A novelresistancegene mapped to a genomic region that contains severalgenes,, 2017, 12: e0169950
[22] Li Y P, Sun S L, Zhong C, Wang X M, Wu X F, Zhu Z D. Genetic mapping and development of co-segregating markers of, which provides resistance toin soybean., 2017, 130: 1223-1233
[23] Cheng Y B, Ma Q B, Ren H L, Xia Q J, Song E L, Tan Z Y, Li S X, Zhang G Y, Nian H. Fine mapping of a- resistance genein soybean (L.) by high- throughput genome-wide sequencing., 2017, 130: 1041–1051
[24] Schmitthenner A F. Problems and progress in control of Phytophthora root rot of soybean., 1985, 69: 362–368
[25] Yang B, Wang Q Q, Jing M F, Guo B D, Wu J W, Wang H N, Wang Y, Lin L, Wang Y, Ye W W, Dong S M, Wang Y C. Distinct regions of theessential effector Avh238 determine its function in cell death activation and plant immunity suppression., 2017, 214: 361–375
[26] Lee S, Mian M A R, Sneller C H, Wang H H, Dorrance A E, McHale L K. Joint linkage QTL analyses for partial resistance toin soybean using six nested inbred populations with heterogeneous conditions., 2014, 127: 429–444
[27] 朱振东, 王化波, 王晓鸣, 武小菲. 黑龙江省主要栽培大豆品种(系)对大豆疫霉根腐病的多抗性评价. 植物遗传资源学报, 2004, 5: 22–25 Zhu Z D, Wang H B, Wang X M, Wu X F. Response of soybean cultivars or lines developed in Heilongjiang province to five strains of., 2004, 5: 22–25 (in Chinese with English abstract)
[28] 朱振东, 王晓鸣, 常汝镇, 马淑梅, 武小菲, 田玉兰. 黑龙江省大豆疫霉菌生理小种鉴定及大豆种质的抗性评价. 中国农业科学, 2000, 33: 62–67 Zhu Z D, Wang X M, Chang R Z, Ma S M, Wu X F, Tian Y L. Identification of race ofand reaction of soybean germplasm resources in Heilongjiang province., 2000, 33: 62–67 (in Chinese with English abstract)
[29] Kyle D E, Nickell C D, Nelson R L, Pedersen W L. Response of soybean accessions from provinces in southern China to., 1998, 82: 555–559
[30] 孙石, 赵晋铭, 武晓玲, 郭娜, 王源超, 盖钧镒, 邢邯. 黄淮海地区大豆种质对疫霉根腐病的抗性分析. 大豆科学, 2008, 27: 465–470 Sun S, Zhao J M, Wu X L, Guo N, Wang Y C, Gai J Y, Xing H. Resistance of soybean gemplasm to phytophthora in Huanghuai Valley., 2008, 27: 465–470 (in Chinese with English abstract)
[31] Lohnes D G, Nickell C D, Schmitthenner A F. Origin of soybean alleles forresistance in China., 1996, 36: 1689–1692
[32] 朱振东, 霍云龙, 王晓鸣, 黄俊斌, 武小菲. 大豆疫霉根腐病抗源筛选. 植物遗传资源学报, 2006, 7: 24–30 Zhu Z D, Huo Y L, Wang X M, Huang J B, Wu X F. Screening for resistance sources to Phytophthora root rot in soybean., 2006, 7: 24–30 (in Chinese with English abstract)
[33] 任海龙, 宋恩亮, 马启彬, 杨存义, 王瑞鹏, 马天翔, 唐玉娟, 年海. 南方三省(区)抗大豆疫霉根腐病野生大豆资源的筛选. 大豆科学, 2010, 29: 1012–1015 Ren H L, Song E L, Ma Q B, Yang C Y, Wang R P, Ma T X, Tang Y J, Nian H. Screening for resistance sources to Phytophthora root rot infrom three provinces of southern China., 2010, 29: 1012–1015 (in Chinese with English abstract)
[34] 程艳波, 马启彬, 牟英辉, 谭志远, 吴鸿, 年海. 华南地区推广应用大豆品种对疫霉根腐病的抗性评价. 华南农业大学学报, 2015, 36: 69–75 Cheng Y B, Ma Q B, Mu Y H, Tan Z Y, Wu H, Nian H. Resistance evaluation of soybean varieties to phytophthora root rot in South China., 2015, 36: 69–75 (in Chinese with English abstract)
[35] 申宏波, 丁俊杰, 于永梅, 姚文秋, 顾鑫, 杨晓贺, 郑天琪, 赵海红. 2009年黑龙江省大豆新品系抗疫霉根腐病鉴定与评价. 大豆科学, 2010, 29: 1087–1090 Shen H B, Ding J J, Yu Y M, Yao W Q, Gu X, Yang X H, Zheng T Q, Zhao H H. Identification and evaluation on soybean lines resistant to phytophthora root rot from Heilongjiang province in 2009., 2010, 29: 1087–1090 (in Chinese with English abstract)
[36] 徐鹏飞, 姜良宇, 李文滨, 张淑珍, 陈维元, 吴俊江, 李岑, 邱丽娟, 常汝镇, 王金生. 黑龙江省大豆品种对大豆疫霉根腐病抗性评价及抗性基因推导. 中国油料作物学报, 2011, 33: 521–526 Xu P F, Jiang L Y, Li W B, Zhang S Z, Chen W Y, Wu J J, Li C, Qiu L J, Chang R Z, Wang J S. Screening on soybean cultivars resistance toand genes postulation in Heilongjiang province., 2011, 33: 521–526 (in Chinese with English abstract)
[37] 唐庆华, 崔林开, 李德龙, 戴婷婷, 阴伟晓, 董莎萌, 邢邯, 郑小波, 王源超. 黄淮地区大豆种质资源对疫霉根腐病的抗病性评价. 中国农业科学, 2010, 43: 2246–2252Tang Q H, Cui L K, Li D L, Dai T T, Yin W X, Dong S M, Xing H, Zheng X B, Wang Y C. Resistance evaluation of soybean germplasm from Huanghuai valley to Phytophthora root rot., 2010, 43: 2246–2252 (in Chinese with English abstract)
Resistance Evaluation to EightIsolates for Major Soybean Cultivars in Huang-Huai-Hai Rivers Valley
LI Xiao-Na1,2,**, SUN Shi2,**, ZHONG Chao2, and HAN Tian-Fu1,2,*
1College of Agriculture, Northeast Agricultural University, Harbin 150030, China;2Key Laboratory of Soybean Biology (Beijing) / Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China
With the application of wheat stubble no-tillage cultivation technology, the occurrence rate of phytophthora root rot of soybean is getting higher in the Huang-Huai-Hai Rivers Valley. Therefore, it is crucial to investigate the resistance of the widely-grown soybean cultivars to phytophthora root rot and to screen disease-resistant resources for soybean improvement. In the current study, a total of 140 soybean cultivars, widely-grown in Huang-Huai-Hai Rivers Valley since the 1950s, were investigated for their response to eight strains ofthrough the hypocotyl inoculation method. Except that six cultivars were susceptible to all strains, 134 cultivars (95.7%) were resistant to 1–8 strains, including 83 cultivars (59.3%) resistant to 6–8 strains. A total of 65 reaction types in response to the eight strains were identified from the 134 resistant cultivars by means of 14 reference genotypes. Five reaction types produced by 19 cultivars fit the reaction-type profile of a known resistance gene or the combination of two genes. The remaining 60 reaction types by 115 cultivars were different from all reaction types of a single resistance gene or the combinations of two genes, indicating that theses cultivars might carry novel resistance gene(s)/gene combination(s) against. These results provide a guide in parent selection for pyramiding resistance genes in soybean cultivars with good agronomic backgrounds.
Soybean; Phytophthora root rot;; Resistance gene; Gene postulation
10.3724/SP.J.1006.2017.01774
本研究由国家自然科学基金项目(31471521), 国家现代农业产业技术体系建设专项(CARS-04), 国家公益性行业(农业)科研专项经费项目(201303018)和中国农业科学院农业科技创新工程资助。
This study was supported by the National Natural Science Foundation of China (31471521), the China Agriculture Research System (CARS-04), the Special Fund for Agro-Scientific Research in the Public Interest (201303018), and the Agricultural Science and Technology Innovation Program of the Chinese Academy of Agricultural Sciences.
韩天富, E-mail: hantianfu@caas.cn**同等贡献(Contributed equally to this work)
李晓那, E-mail: 913714351@qq.com; 孙石, E-mail: sunshi@cass.cn
2017-04-12; Accepted(接受日期): 2017-07-23; Published online(网络出版日期): 2017-08-10.
http://kns.cnki.net/kcms/detail/11.1809.S.20170810.1616.012.html
