朱砂叶螨危害对木薯幼苗光合作用及叶绿素荧光参数的影响
2018-01-16梁琼月董蒙蒙顾明华
梁琼月,董蒙蒙,何 冰,顾明华,陈 立,杨 娟,赖 艳,曾 玮
(1.广西大学农学院,广西 南宁 530004;
2.广西大学植物科学国家级实验教学示范中心,广西 南宁 530004)
木薯(Manihot esculentaCrantz)被称为世界三大薯类作物之一,除了作为粮食食用以外,其另一大用处为生物燃料——乙醇的生产原料。近年来,木薯栽培遭到以朱砂叶螨(Tetranychus cinnabarinus)为首的螨虫危害严重,其中华南等地区木薯朱砂叶螨危害成灾,严重时可致木薯产量降低50%~70%[1]。朱砂叶螨民间称之为红蜘蛛,是遍布全世界的害虫[2],木薯植株受取食后将出现生长发育不良,减产严重,甚至颗粒无收。国内外对木薯害螨的研究热点集中于木薯单爪螨(Mononychellus tanajoa)方面[3-6],国内偏向于防控检疫性害螨,如木薯单爪螨侵入危害方面[7-13],关于朱砂叶螨取食危害与寄主木薯的光合作用生理反应的研究鲜见报道。
植物生长过程中必须依靠光合作用来提供能量物质。光合作用作为植物生理学方面研究的关键性问题[14],这一生理过程极易受外界环境变化的影响[15]。叶绿素荧光参数是植物光合作用和环境的关系的内在反映,从更深程度解析植物进行光合作用时光系统对光能的吸收到利用的整个过程深入反映其内在本质特征[16-17]。因此,我们研究了朱砂叶螨危害对木薯4个品种(华南205、南植199、桂热891、华南8号)幼苗光合作用和叶绿素荧光参数的影响,以期为木薯朱砂叶螨的防治及抗性品种的筛选提供理论依据。
1 材料与方法
1.1 试验材料
供试植物为盆栽木薯4个品种:华南8号、南植199、桂热891、华南205。各品种木薯分别种植于塑料盆(上口径 15 cm)内,每盆2株,以无菌泥炭土为种植基质,用防虫罩各盆隔离栽培于玻璃温室内,温度28(±2)℃,相对湿度70(±5)%,水分、肥料管理按常规栽培进行,植株生长至7~9片完全展开叶时进行试验处理。
供试昆虫为朱砂叶螨,于2016年5月在武鸣采集,隔离饲养于室内。试验虫龄为2~3 龄虫,从饲养室内继代种群中随机挑取,用毛笔将其移接至试验木薯正4~8叶片上。
1.2 试验方法
2016年4月20日栽种木薯,6月15日于每个木薯品种中选取未受螨虫或其他病虫危害的3盆(每盆1株)长势相当的木薯健康植株,接种虫量10头/株,随机释放,让其自由取食叶片,接种后的各木薯植株分别采用防虫罩隔离栽培。每个处理3次重复。分别在接种螨虫后5、10 d对木薯叶片进行光合作用相关指标和叶绿素荧光参数的测定,以健康植株为对照。试验地点为广西大学农学院教学科研基地玻璃网室内。
光合作用的测定采用便携式光合作用测定仪(LI-6400XT,LI-COR,美国)分别对木薯叶片(正5叶,功能叶)光合相关指标进行测定,测定时,设定光强PAR为1 500 umol/m2·s,温湿度采用自然状态。试验采用不同处理交替进行测量避免环境因素变化影响测量结果。试验时间在6月1~7日上午9:00~11:00,天气晴朗无云。
叶绿素荧光参数测定采用叶绿素荧光分析仪(PAM-2500,WALZ,德国)分别对木薯叶片(正5叶,功能叶)叶绿素荧光参数进行测定。试验时间在6月1~7日凌晨。
试验结果采用Excel、SPSS软件进行处理分析。
2 结果与分析
2.1 朱砂叶螨危害对木薯叶片气体交换参数的影响
由图1可知,供试木薯品种受朱砂叶螨危害后叶片净光合速率(Pn)、气孔导度(Gs)和蒸腾速率(Tr)均低于健康植株,差异显著,且随着受害时间的延长呈下降趋势;而胞间CO2浓度(Ci)则反之,受害植株均高于正常健康植株,且受害时间越长其差异也越明显。4个不同基因型木薯在受害10 d后,气体交换参数的变化趋势一致,其中受害叶片Pn、Gs和Tr比健康植株降低最为显著的是桂热891,分别降低91.3%、95.0%和80.1%,而胞间Ci则以华南205上升最显著,达45.2%。
2.2 朱砂叶螨危害对木薯叶片叶绿素荧光参数的影响
由图2可知,木薯对朱砂叶螨的危害的反应并非因品种而异,受危害后F0含量均呈现上升趋势,两个不同时间段含量均显著高于健康植株;而Fm值则有所下降,受危害后5 d以华南205下降最为显著,其他各品种与健康植株未达差异显著水平,受害后10 d 4个品种受害植株与健康植株间Fm值未达差异显著水平;同时Fv/Fm也下降,变化趋势与Fm一致;随着木薯的生长发育,F0和Fm有所降低。同时本试验4个不同基因型木薯在受害后10 d,各受害植株间、各健康植株间叶绿素荧光参数F0,Fm和Fv/ Fm均未达差异显著水平。有研究认为F0值增大说明光系统Ⅱ中RC受损坏,或永久失活。通常植株受到胁迫,F0值增大,本研究结果也显示木薯植株受危害后F0值有所上升。可见F0、Fm和Fv/Fm值在植物生长发育过程中并非简单的增加或降低,其变化趋势因植物种类而异。
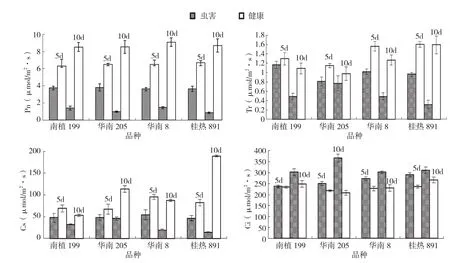
图1 朱砂叶螨危害对木薯叶片气体交换参数的影响

图2 朱砂叶螨危害对木薯叶片叶绿素荧光参数的影响
3 结论与讨论
植物生长发育过程中,光合作用在生理代谢进程中发挥极其重要的作用,光合水平直接影响植物的生长发育及其抗逆性[18]。植物光合作用过程中对二氧化碳和水的运输及调控是通过气孔完成,气孔限制或非气孔限制均直接致使光合能力降低。植物叶片Ci是评估其光合作用是受气孔或非气孔限制的关键参数,Ci降低为气孔限制,Ci 上升为非气孔限制[19]。同时前人发现,植物受病虫侵害后,致使叶片Gs值上升,Pn减弱,如昆虫危害云杉[20]、叶甲危害红荆[21];同时也有研究表明植株受虫害导致叶片Gs降低,如鳞翅目取食苹果树[22]、瘿螨危害糖枫树[23]和缨翅目危害梨树[24]。而二斑叶螨对棉花危害初期Gs降低,后期Gs上升[25]。可见,病虫危害对植物光合指标的影响因植物种类、危害时期和强度而异。本研究中,不同木薯品种受朱砂叶螨取食危害后,叶片的Pn、Gs均降低,而Ci增加,可见Pn降低可能是由于非气孔限制引起的叶肉细胞同化能力降低。
植物长期处于较严重的胁迫状态,光合强度受限多为非气孔限制所致,光合代谢过程受抑制和损伤,如叶片光合放氧能力、叶绿体Hill反应和叶绿素荧光强度显著降低等[19]。叶绿素荧光参数反映了光合系统PSⅡ的电子传递吸收光化学量子效率。光系统Ⅱ的光能转化效率是反映植物潜在光合效率高低的指标,常被用于评价植物受环境胁迫程度的指标。有研究表明不同植物种类受病虫危害后,叶片Fv/Fm值因植物种类而异,如绿斑病藻侵染夏橙[26]、黄瓜花叶病毒侵染烟草[27]均降低了Fv/Fm值;而鳞翅类幼虫取食冬青[28]、茶尺蠖取食茶树[29]和大蜗牛危害黄瓜[30]后Fv/Fm值则升高。Fo和Fm表示光系统Ⅱ反应中心处于完全打开或完全关闭状态的荧光产量,PS反应中心受损坏坏或可逆失活致使F0值上升,而PS热能耗散增多F0值则降低。本研究结果表明,供试木薯品种受朱砂叶螨危害后Fv/Fm值降低,且随着危害时间的延长而下降,说明在朱砂叶螨的危害下木薯叶片发生了光抑制或PSⅡ损害,同时F0水平呈上升的趋势,说明光抑制或PSⅡ损害程度较严重。
从本研究可见,朱砂叶螨危害对木薯光合机制产生一定的影响,部分气体交换参数和叶绿素荧光参数值降低,净光合速率降低主要是由于非气孔限制所致,叶绿素荧光参数值的变化可能是木薯叶片发生光抑制或PSⅡ损害导致。
[1]陈青,卢芙萍,黄贵修,等.木薯害虫普查及其安全性评估[J].热带作物学,2010,31(5):819-827.
[2]李隆术,李云瑞.蜱螨学[M].重庆:重庆出版社,1988:4-129.
[3]Henrik Skovgård,Jonna Tomkiewicz,Gösta Nachman,et al.The effect of the cassave green miteMononychellus tanajoaon the growth and yeild of cassavaManihot esculentain a seasonally dry area in Kenya[J].Experimental & Applied Acarology,1993(17):41-58.
[4]Akinlosotu T A.Seasonal trend of green spider mite,Mononychellus tanajoapopulation on cassava,Manihot esculentaand its relationship with weather factors at moor plantation[J].Insect Sci.Application,1982,3(4):251-254.
[5]Costa E C,Teodoro A V,Rêgo A S,et.al.Population structure and dynamics of the cassava green miteMononychellus tanajoa(Bondar)and the predatorEuseius ho(DeLeon)(Acari:Tetranychidae,Phytoseiidae)[J].Arthropods,2012,1(2):55-62.
[6]Omorusi V I,Ayanru D K G.Effect of NPK fertilizer on diseases,pests and mycorrhizal symbiosis in cassava[J].Int.J.Agric.Biol,2011(13):391-395.
[7]卢辉,陈青,卢芙萍,等.基于Maxent模型的木薯单爪螨在云南的适生性评价(英文)[J].Agricultural Science & Technology,2011,12(12):1905-1908.
[8]卢辉,陈青,卢芙萍,等.木薯单爪螨全球潜在地理分布的MaxEnt预测[J].植物检疫,2012,26(1):1-6.
[9]经福林,李迁,卢芙萍,等.木薯单爪螨的寄主选择性研究[J].热带作物学,2013,34(2):344-347.
[10]卢芙萍,符悦冠,黄贵修,等.温度对木薯单爪螨生长发育与繁殖的影响[J].热带作物学报,2011,32(9):1720-1724.
[11]卢芙萍,符悦冠,经福林,等.卵高温胁迫对木薯单爪螨发育与繁殖的影响研究[J].中国农学通报,2012,28(21):229-236.
[12]黄洁,范伟峰,刘茂,等.531份木薯种质的朱砂时螨抗性评价[J].广东农业科学,2011,38(2):27-28.
[13]韦威旭,覃维治,韦民政,等.木薯品种对二点红蜘蛛的抗性观察[J].广西农业科学,2009,40(5):504-506.
[14]Nabity P D,Zavala J A,Lawlor D W.Musings about the effects of environment on photosynthesis[J].Annals of Botany,2009,103:543-549.
[15]Samsone I,Andersone U,Ievinsh G.Variable effect of arthropod-induced galls on photochemistry of photosynthesis,oxidative enzyme activity and ethylene production in tree leaf tissues[J].Environmental and Experimental Biology,2012,10:15 - 26.
[16]李晓,冯伟,曾晓春.叶绿素荧光分析技术及应用进展[J].西北植物学报,2006,26(10):2186-2196.
[17]陈建明,俞晓平,程家安.叶绿素荧光动力学及其在植物抗逆生理研究中的应用[J].浙江农业学报,2006,18 (1):51-55.
[18]惠红霞,许兴,李前荣.外源甜菜碱对盐胁迫下枸杞光合功能的改善[J].西北植物学报,2003,23(12):2137 -2422.
[19]FarquharG D,SharkeyTD.Stomatal conductance and photosynthesis[J].AnnualReview PlantPhysiology and PlantMolecular,1982(33):317.
[20]Ozaki K,Saito H,Yamamuro K.Compensatory photosynthesis as a response to partial debudding in ezo spruce,Picea jezoensis seedlings[J].Ecol Res,2004,19(2):225-231.
[21]Pattison R R,D'Antonio C M,Dudley T L.Biological control reduces growth,and alters water relations of the saltcedar tree(Tamarixspp.)in western Nevada,USA[J].J Arid Environ,2011,75(4):346-352.
[22]Pincebourde S,Frak E,Sinoquet H,et al.Herbivory mitigation through increased wateruse efficiency in a leaf-mining moth-apple tree relationship[J].Plant Cell Environment,2006,29(12):2238-2247.
[23]Patankar R,Thomas S C,Smith S M.A gallinducing arthropod drives declines in canopy tree photosynthesis[J].Oecologia Nov,2011,167(3):701-709.
[24]Ellsworth D S,Tyree M T,Parker B L,et al.Photosynthesis and water-use efficiency of sugar maple(Acer saccharum)in relation to pear thrips defoliation[J].Tree Physiol,1994,14(6):619-632.
[25]Reddall A,Sadras V O,Wilson L J,et al.Physiological responses of cotton to two-spotted spider mite damage[J].Crop Sci,2004,44(3):835-846.
[26]王大平,曾明,朱钧,等.绿斑病藻寄生对夏橙叶片光合作用特性的影响[J].应用生态学报,2006,17(6):1141-1144.
[27]王春梅,施定基,朱水芳,等.黄瓜花叶病毒对烟草叶片和叶绿体光合活性的影响[J].植物学报,2000,42(4):388-392.
[28]Zhang B,Zhou X,Zhou L,et al.A global synthesis of below-ground arbon responses to biotic disturbance:a meta-analysis[J].GlobalEcology and Biogeography,2014,24(2):126-138.
[29]韦朝领,童鑫,高香凤,等.茶树对茶尺蠖取食危害的补偿光合生理反应研究[J].安徽农业大学学报,2007,34(3):355-359.
[30]Thomson V,Cunningham S,Ball M,et al.Compensation for herbivory by Cucumis sativus through increased photosynthetic capacityand efficiency[J].Oecologia,2003,134(2):167-175.
