音乐增强大脑网络小世界特性
2018-01-10何裕嘉郑高兴于玉国
何裕嘉,张 玮,郑高兴,于玉国,2
(1.复旦大学 生命科学学院 生理学和生物物理学系 脑科学协同创新中心,上海 200438;2.复旦大学 计算系统生物学中心,上海 200433)
音乐增强大脑网络小世界特性
何裕嘉1,张 玮1,郑高兴1,于玉国1,2
(1.复旦大学 生命科学学院 生理学和生物物理学系 脑科学协同创新中心,上海 200438;2.复旦大学 计算系统生物学中心,上海 200433)
近年来实验发现音乐相比其他机械噪声对人脑感知系统更能增强大脑内部复杂网络特性.优美音乐在频谱上普遍具有1/f统计特征.在音乐增强脑电信号记录分析中,我们发现对比具有1/f特征的音乐信号,在仍保留1/f特征的随机乱乐刺激下,大部分脑皮层区域的脑功能网络连接密度普遍下降,并且增强的脑功能网络小世界特性在一定阈值范围内也会有显著下降.随机打乱的音乐虽然仍保留长程相关特征,但打乱后每分钟节拍数和节拍清晰度出现了明显降低.这两种音乐特性的降低与音乐打乱前后的大脑小世界网络统计指标的CMean/LMean降低有显著关联.说明音乐信号除了1/f长程相关统计特征之外的其他有效音乐信息在增强脑功能网络方面也起到重要作用.
音乐; 脑电信号; 1/f统计特性; 功能连接密度; 小世界网络特性
人类大脑是自然界通过漫长进化而产生的最复杂和最智能化的器官,具有计算和信息处理、意识和高级认知等功能[1].解析其工作原理已经成为近一个世纪以来脑科学、物理学、数学、计算机工程学和医学等众多科学领域的最前沿挑战课题之一.对脑功能的破译需要在多个层次上解析大脑网络系统的连接方式、规则和特性.阐明大脑多层次网络复杂结构及其功能的“线路设计图”,将为解析大脑工作原理开启一扇门[2-3].从单个脑神经元层次到皮层神经网络到跨皮层超级网络,是神经元的自发放电活动和受外界输入信号产生的兴奋性或抑制性电化学活动形成了网络之间的功能连接.有效的外界信号会驱动区域内和区域间的神经元集群产生放电活动.放电活动通过神经突触连接影响神经元之间的信息通讯,并形成对外界信号的编码和表征.这些区域同步或时空相关的脑电活动会将不同空间分布的脑皮层网络连接起来,形成随时空动态变化的功能连接网络,并产生复杂的跨时空尺度的复杂网络动力学现象.外界环境的输入信号可能拥有一些复杂特征,会增强大脑内部神经网络的放电活动,可以通过对输入信号和大脑响应输出信号的分析来研究大脑的功能网络[4-5].
长期以来,白噪声(具有平坦的功率谱密度,信号成分在各个频率具有近乎相同的功率,在时域上缺乏长程相关性)作为探针信号被广泛用来探测未知系统如大脑各个层次神经系统的输入输出函数[4].然而近年来的实验研究表明,大脑视觉、听觉和嗅觉等感知系统对自然界信号的响应更加敏感和强烈[6-8].实验结果表明,应用类自然界信号作为探针、结合沃泰拉(Volterra)核函数辨识方法可以更精确地反映神经系统的本征功能函数[9].自然界的信号在统计特性上普遍具有长程相关性(或1/f特征,其中f代表信号频率,1/f特征表明信号功率谱密度或功率随频率呈反幂律下降关系)[10-13].最近的实验和理论揭示,感知神经系统可能对具有1/f统计特征的类自然界信号都具有更强的敏感性[14-16].这使得应用类自然1/f信号来研究神经系统网络连接和功能特性成为必要.
音乐是否可以作为一个有效探针信号来充分增强脑跨皮层神经网络的复杂特性呢?近期已经有实验发现,优美的中国古典音乐可以诱导出复杂的脑网络连接,增强大脑的连通性,提升大脑跨皮层信息流传输,使大脑功能网络呈现出复杂的小世界网络特性[17-18].小世界网络理论(WS理论)最早由Watts和Strogatz在1998年提出[19],该理论描述了小世界网络(WS网络)物理结构介于规则网络和随机网络之间,但却具备了两种网络的优点.规则网络即网络节点以特定规则严格与近邻相连接,相距较远的节点要通过数条边才能建立连接.随机网络即网络节点完全随机建立连接.WS网络是在规则网络结构基础上,任意长程节点之间出现了可以随机连接概率,由此WS网络具有了较短的网络平均路径长度,以及和规则网络相似的网络集聚系数.现实世界中的生物网络、社会网络、交通网络、脑神经网络和传染疾病传播网络等都具有WS特性.由于WS网络通常具有较大的集聚系数和较小的平均最短路径长度,这使得信息流在WS网络中传输比在规则网络和随机网络都更加高效[20-21].已有数据分析发现大量的人工创作的优美音乐在频谱上都具有1/f统计特征[10],为了探究1/f特征以及其他音乐特征是否是音乐增强大脑小世界特征的关键因素,我们设计了实验,研究大脑对正序音乐和被随机打乱音乐(时间序列仍保留1/f特征)的响应,以及对增强大脑小世界网络特性的差异.我们使用64通道有线脑电图(Electroencephalogram, EEG)记录设备收集数据,考察了12名测试者对两首符合1/f频谱特征的优美音乐和对应随机乱序音乐的大脑EEG功能网络连通性,包括基于时间偏移的交叉相关函数分析,功能连接密度计算和小世界网络统计特性分析.我们改进了之前对脑电EEG信号的预处理方法和脑网络小世界特性的分析方法[22],希望能更有效地反映出任务态下的脑网络功能复杂连接线路和传输效率.
1 材料与方法
1.1 被试选取与实验准备
脑电实验被试为12名在校本科生与研究生,其中7名男生,5名女生,均为右利手.平均年龄21.75岁(SD=2.93,年龄范围从19岁到27岁).所有的被试都是与音乐无关专业的学生,其中仅有两位同学接受过音乐相关训练.被试均无神经系统疾病且矫正视力及听力均正常.
实验采用德国Brain Products公司的actiCHamp 64通道有线脑电设备,参考电极为顶叶中心位置Cz电极处.噪声声音播放采用漫步者(EDIFIER)R2000DB 5吋2.0音箱,左右音箱分别位于被试前方左右1m处.声音播放程序采用Eprime 2.0软件播放,并与actiCHamp脑电设备通过并口连接可以自动打标记.脑电刺激采集系统包括刺激呈现电脑,带有接收EEG信号的电极帽,BP放大器以及EEG信号记录电脑等.刺激呈现电脑用Eprime播放声音片段,并将每个播放片段的时间点标记通过放大器传输到脑电记录电脑.64通道电极帽所采集的脑电信号同样经过放大器传输到脑电记录电脑.
实验所采用的音乐文件均下载于网易云音乐软件,2首原始音乐片段分别为“Castle in the Sky”(天空之城)钢琴版,选择时长1min 56.892s和“The Blue Danube”(蓝色多瑙河),时长1min 42.263s.对应打乱音乐的片段由每段原始音乐片段先以1s为间隔分别切割成117段和103段(最后多出的0.892s和0.263s是单独的一段),再完全随机排列这些1s的小切片合成新的噪声音乐序列.对应打乱的音乐片段保证每个1s的小片段出现且仅出现1次,所以顺序打乱的音乐与相对应的原始音乐时长相等,且功率谱分析显示其高频成分(≥1Hz)基本保留了1/f特征.所有音频的截取、切割、随机排序等处理均用MATLAB完成.
1.2 实验流程
被试与刺激系统均会处于声音屏蔽房间中,被试穿戴有线64通道电极帽,并先确保接地电极和头顶Cz电极位置正确,接着确保其他电极位置正确.主试用平头针向电极注射导电膏优化EEG采集导联性,并观测检测软件界面,确保每个电极导联的阻抗最终稳定在10kΩ以下.
实验开始前要进一步测试脑电导联情况来预判数据是否可靠,被试会被要求分别闭眼一分钟和睁眼一分钟.主试根据屏幕显示的脑电波是否在闭眼时会出现明显呈锯齿状的α波(约10Hz),以及在睁眼时会出现明显的α波阻断来判断实验受试者和实验设备是否符合数据采集标准.未出现α波明显阻断的受试者数据会被抛弃不用.
实验过程如下.在实验前,主试将调整室内灯光到适宜,让被试注意观看刺激呈现电脑屏幕显示的提示信息.被试将看到实验流程示意图.当被试认为调整到实验状态后,可按下Enter键开始实验.实验刺激程序按照次序分为闭眼静息1min,睁眼静息1min,播放第1段音乐原始片段,中间间隔1min,再播放第1段音乐打乱片段;然后闭眼静息1min,睁眼静息1min,播放第2段音乐原始片段,中间间隔1min,播放第2段音乐打乱片段,闭眼静息1min,睁眼静息1min.其中,每段音乐刺激时,被试是睁眼收听,但在实验开始前,被试将会被提示尽量减少眼球运动.每段刺激结束后刺激呈现电脑屏幕均有提示文字,另外,在闭眼静息1min结束后被试会听到‘滴’地一声结束提示音.每段刺激过程之间的间隙被试均可以做适当调整和放松,然后确认状态后再依据提示按下回车Enter键进行下一段实验.除此之外,被试无需做任何操作,只要求被试保持心态平静,身体尽可能保持不动,注意聆听所播放的音乐即可.每次被试按下回车键后EEG信号记录电脑将会自动打下标记,由于静息和听音乐的时间都是固定的,所以由开始的标记就可以推算出每段刺激结束的时间点.
1.3 数据分析
预处理采用MATLAB中的EEGLAB工具箱,依次进行: 基线校正;49~51Hz陷波,去除50Hz工频干扰;0.1~100Hz滤波;数据分段,按照标记时间信息,将数据切分出来;利用独立分量分析(Independent Component Analysis, ICA)算法配合Adjust 1.1插件[23],去除肌电、眼电和眼动等伪迹.
为评估EEG功能连接,我们首先计算EEG相关情况[24],即计算两两电极间EEG信号的互相关.对于信号长度为N的任意两段EEG信号xn和yn,互相关函数为一个2N-1的向量:
(1)
接着将互相关函数归一化,使得所有序列在0延时的自相关均为1.
网络分析方法部分.首先我们遍历计算出63个通道EEG信号所有两两互相关函数序列,并取各个最大值绘制成邻接矩阵.邻接矩阵中第i行表示第i号电极与其他电极的相关值,即为功能连接强度.那么我们将第i号电极与其他所有电极(除自身以外)功能连接强度的平均值记为第i号电极的功能连接密度.从而我们可以依次求出每一根电极所确定的功能连接密度.进一步,计算小世界网络[19]统计特性,即分析平均集聚系数(clustering coefficient)来评定网络的功能分隔性和分析平均最短路径(shortest path length)来评定网络的功能整合性[25],其中最短路径的计算采用Floyd算法[26],并将需要比较的系数对完全随机网络对应系数进行归一化.网络的平均最短路径可以较好的描述网络的连通性.网络中节点i和节点j之间的距离dij,即是连接两节点最短路径边数.平均最短路径长度L,为网络中任意两点间最短路径的平均值,即
(2)
其中N为网络的节点数.聚类系数是来表征网络节点聚类特性的,正常情况下,假设网络中的节点i与ki条边相关联,很明显,在这ki个节点之间,最多有ki(ki-1)/2条边,而这ki个节点间存在的边数实际为EI.则这ki个节点之间存在的实际边数EI与总的最大的可能边数ki(ki-1)/2之比,定义为节点i的聚类系数Ci,即
(3)
再对网络中所有节点的聚类系数Ci取平均值,得到的就是整个网络的聚类系数CMean,即
(4)
其中N为网络的节点数.由于对比随机网络,小世界网络的CMean较大,而LMean会较小,因此本文中将用CMean/LMean来表征小世界特性.
另外本文研究脑功能网络属于无权重网络,所记录任何两个电极之间的脑电信号之间的互相关系数的最大值在归一化后分布在0~1之间.网络中任意两节点之间有两种状态,一种是有连接,另一种是无连接,而我们定义阈值就是为了考察网络中有连接的节点之间的网络特性.为了区分有连接和无连接,我们需要定义阈值.当两节点之间的连接强度大于等于这个阈值时,划分为两节点有连接,小于这个阈值时,划分为两节点没有连接.在脑电中,63个电极两两之间的相关系数大于等于某个阈值时认为这两个电极之间存在连接,这样就能建立起脑网络,研究小世界特性.
2 结 果
2.1 预处理后脑电图示意

预处理过后的脑电图去除了肌电、眼电和眼动等伪迹和工频干扰,没有漂移和巨幅震荡等情况,通常较为稳定的处于-80~100μV之间,且清醒状态下,认知过程会呈现一定周期性的震荡,如(图1)所示,为第1位被试在接收第1段正序音乐刺激时AFz和O2两个电极的电压时间序列图节选(采样率: 1000Hz).
2.2 功能连接密度
接着,计算每位被试EEG在时域下两两电极间相关函数的最大值,并按照每根电极汇总制作不同音乐刺激条件下功能连接密度脑地形图(图2),整体范围为0.3~0.6,得到描述性统计(均值±均方误差)如下: 0.4345±0.0123(a1);0.4307±0.0112(b1);0.4357±0.0125(a2);0.4145±0.0106(b2),均为12名被试的叠加平均.这说明乱乐刺激下EEG的功能连接密度会下降,我们对两首音乐以及其对应乱乐刺激下叠加平均的功能连接密度脑地形图,分别做基于电极功能连接密度的配对t检验,得到两首音乐的乱序情况刺激下,功能连接密度均是显著下降的,对应P值分别为P=0.018和P=1.81×10-16.
2.3 小世界网络统计特性
然后,分析EEG信号对应的跨皮层脑区小世界网络统计特征,计算平均网络集聚系数(CMean)和平均最短路径(LMean),图3给出了不同阈值(0.01~0.99)下小世界特性的变化曲线CMean/LMean(图3).其中,为了区分两个节点之间是否存在网络连接和无连接,我们需要定义阈值.当两节点之间的交叉相关强度大于等于这个阈值时,定义为两节点有连接,小于这个阈值时,定义为两节点没有连接.我们使用双因素方差分析(two-way ANOVA),分别对在两首音乐以及其对应乱乐刺激下的EEG活动的小世界网络进行统计检验.我们发现在较广阔的阈值范围内,音乐刺激下EEG网络均比对应度数的随机网络拥有更好的小世界特性.其中音乐1与其对应随机音乐白谱序列在阈值范围(0.25~0.93)内有显著性差异P<0.05(b1),音乐2与其对应随机音乐白谱序列在阈值范围(0.28~0.93)内有显著性差异P<0.05(b2),这也侧面印证EEG本身具备一些小世界网络统计特性.另外我们发现,当音乐2被打乱后,EEG网络的小世界特性在一定阈值范围(0.34~0.39)内会显著(P<0.05)下降(b2).然而,对于被打乱的音乐1刺激,EEG网络的小世界网络特性在一定阈值范围内在均值层面有所下降(a1),尽管差值在统计上还不显著(b1).
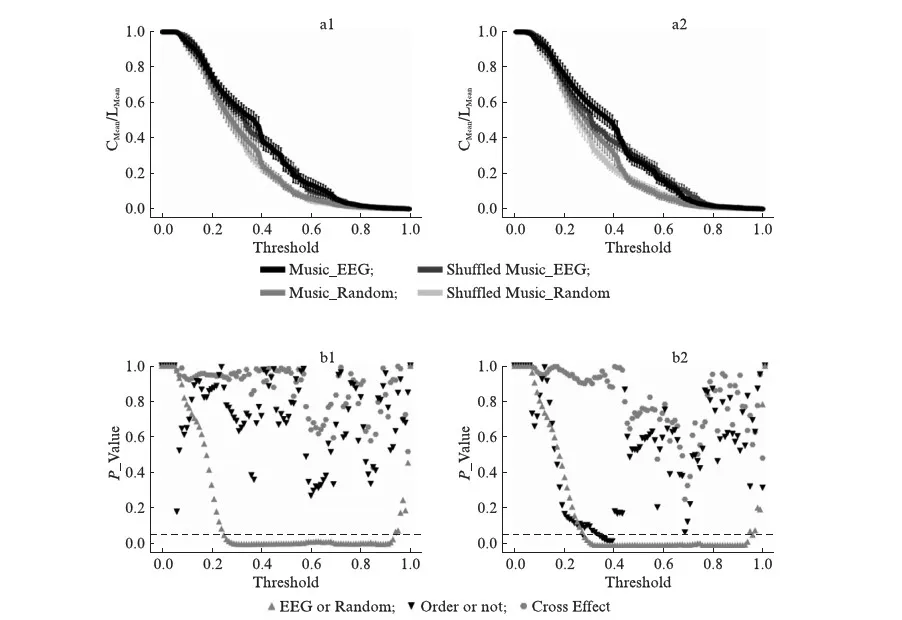
图3 脑电小世界网络统计Fig.3 EEG small-world network statisticsEEG网络小世界特性(定义为平均集聚系数除以平均最短路径)在不同阈值下的取值.a1: 原始音乐1以及其对应的乱序版本刺激;a2: 原始音乐2以及其对应的乱序版本刺激.图中还增加了EEG网络对应拓扑性质(度数相同,孤立点数相同)的随机网络作为参考.误差棒代表均方误差.使用双因素方差分析,对a1及a2做检验,分别得到b1和b2(依阈值对应分析,b1和b2中虚线代表P=0.05).

图4 脑电小世界网络的进一步统计验证Fig.4 Further statistical validation of EEG small-world network图中虚线代表P=0.05.
进一步,我们再使用双因素方差分析(two-way ANOVA),重点考察两首音乐间差异,以及音乐与对应乱乐的差异这2个因素的统计显著性.发现接受乱乐刺激这个因素后,EEG网络的小世界网络特性在前述(图3,b2)相同阈值范围(0.34~0.39)内会显著(P<0.05)下降(图4).从而可以得出结论,接收乱序音乐刺激相比正序音乐刺激,会弱化EEG网络小世界特性形成,宏观上也会降低人脑信息传递与整合效率.
事实上,正常播放音乐增强到较高水平的小世界网络特性是通过增强的网络集聚系数和降低了网络最短路径来实现的(图5,图6).我们使用双因素方差分析(two-way ANOVA),分别对在两首音乐以及其对应乱乐刺激下的EEG活动的小世界网络进行统计检验.我们发现在较广阔的阈值范围内,音乐刺激下EEG网络均比对应度数的随机网络拥有更大的平均集聚系数(图5(b),图6(b)).其中音乐1与其对应乱乐在阈值范围(0.2~0.88)内对应的脑电小世界网络有显著性差异P<0.05(图5(d)),音乐2与其对应乱乐在阈值范围(0.2~0.91)内有显著性差异P<0.05(图6(d)),这也侧面印证信号增强的脑电呈现出较突出的小世界网络特性,同已有报道相符[2].由于我们的构造保证了对应随机网络的度数和拓扑性质(孤立点数)与实际EEG网络相同,所以平均最短路径无差异(图5(a),图6(a)).另外我们发现,当音乐1被打乱后,EEG网络的平均最短路径在一定阈值范围(0.36~0.37)内会显著(P<0.05)上升(图5(c)).当音乐2被打乱后,EEG网络的平均最短路径在一定阈值范围(0.31~0.39)内会显著(P<0.05)上升(图6(c)).然而,对于被打乱的音乐刺激,EEG网络的平均集聚系数在一定阈值范围内在均值层面有所下降(图5(b),图6(b)),虽然差异并不显著(图5(d),图6(d)).我们计划在接下来的实验中研究闭眼听音乐和白噪声乱序音乐的差异,以及测试更多的乐曲来检验音乐所包含的对增强人脑功能网络的关键因素.

图5 音乐1脑电小世界网络统计Fig.5 EEG small-world network statistics of music 1原始音乐1以及其对应的乱序版本刺激下,不同阈值内EEG网络小世界特性统计图.a1: 平均最短路径;a2: 平均集聚系数.图中还增加了EEG网络对应拓扑性质(度数相同,孤立点数相同)的随机网络作为参考.误差棒代表均方误差.使用双因素方差分析,对a1及a2做检验,分别得到b1和b2(依阈值对应分析,b1和b2中虚线代表P=0.05).
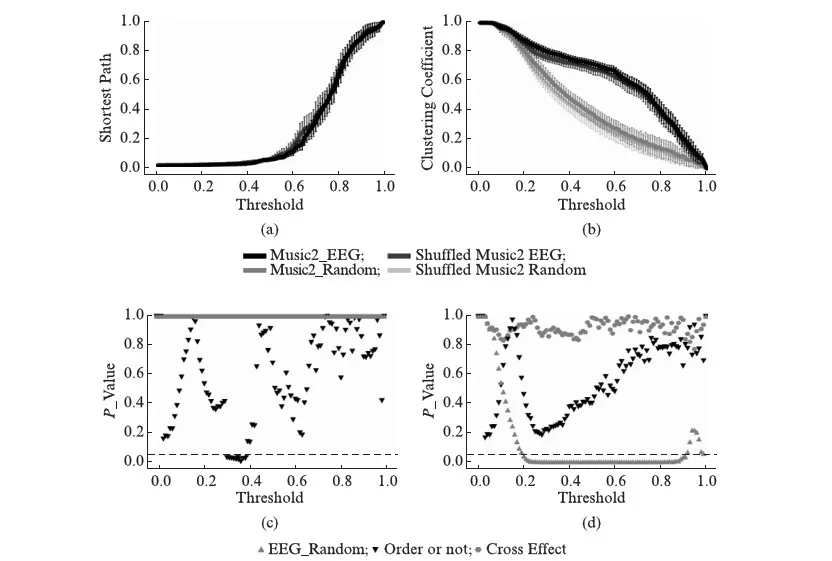
图6 音乐2脑电小世界网络统计Fig.6 EEG small-world network statistics of music 2原始音乐2以及其对应的乱序版本刺激下,不同阈值内EEG网络小世界特性统计图.a1: 平均最短路径;a2: 平均集聚系数.图中还增加了EEG网络对应拓扑性质(度数相同,孤立点数相同)的随机网络作为参考.误差棒代表均方误差.使用双因素方差分析,对a1及a2做检验,分别得到b1和b2(依阈值对应分析,b1和b2中虚线代表P=0.05).
我们分析了两首音乐在打乱前和打乱后的音乐属性,我们发现有2个音乐指标: 每分钟节拍数(通常范围为40~250)和节拍清晰度(取值范围0~1)出现了明显降低.音乐1的每分钟节拍数从136降低到了打乱后的128,降低了5.88%(以未打乱前100%为标准);音乐2的每分钟节拍数从112降低到了打乱后的107,降低了4.46%(以未打乱前100%为标准),见图7(a).音乐1的节拍清晰度则从正常的0.3887降低到了0.316(降低18.7%),音乐2的节拍清晰度则从正常的0.2595降低到了0.231(降低10.09%),见图7(b).我们进而将这两种音乐特性的降低与音乐打乱前后的大脑小世界网络统计指标的CMean/LMean降低做了关联性分析(图7).图中可以看出,对于2首音乐,打乱前后两个音乐节奏属性(每分钟节拍数和节拍清晰度)的降低与小世界特性的降低是直接相关的.

图7 音乐节奏属性与脑电小世界统计指标比例图Fig.7 Scale map of music rhythm property with EEG small-world network statistics index(a) 两首音乐及其乱乐下每分钟节拍数与脑电小世界网络指标CMean/LMean的比例图;(b) 两首音乐及其乱乐下节拍清晰度与脑电小世界网络指标CMean/LMean的比例图;对于任意一首音乐,以其音乐节奏属性与脑电小世界统计指标为基准(100%),计算出其对应乱序音乐下的音乐节奏属性与脑电小世界统计指标所占的比例.
3 讨 论
音乐是一种非常优美和特殊的人工创作的信号,它的音频、节拍和有规律的声波振动,能产生一种物理能量作用于人的听觉系统,产生美妙的共鸣,并驱使身体生理、心理和大脑神经系统产生轻松、愉悦(或伤感)和安宁(或激动)等情感[27].这使得古今中外的医学家们开创了音乐治疗这个方向,尝试使用音乐对那些有心理抑郁、情绪低落、生理疼痛、甚至精神紊乱的患者进行心理和生理调节、甚至达到治疗的目的.近年来著名的医学期刊柳叶刀杂志综述了大量临床实验研究表明[28-29],音乐的确能有效改善受试者的疼痛、消极心态、睡眠质量和生活态度.因此研究音乐对大脑神经网络的作用效果和增强复杂网络的特性可能会提供音乐治疗方面的机制,尤其值得深入和系统的研究.
在过去几十年中,通过脑功能成像等技术临床研究音乐感知对其神经相关性机制已经引起了极大关注.这些研究中的大多数主要基于音乐处理模块化的概念[30],其中模块对应于特定脑区,包括音乐特征分析,听觉记忆,听觉场景分析,以及音乐语法和语义的处理[31].在本研究中,我们尝试从另外的角度来观察音乐对大脑网络的作用.音乐因其节律和旋律具备了时间上的长时程关联性质,在频域空间具有和自然信号相似的高阶统计特性.我们之前的研究发现自然界信号中的1/f特征可能是增强脑认知反应的关键因素.优美的音乐不仅仅具备了同自然信号相似的1/f特征,可能还包含了非常丰富的文化、节拍、旋律等关键因素,会增强人脑的反应.因此,本研究选择了在我们日常生活中听到的音乐作为听觉刺激以重建实验室环境中的类自然信号.通过被试者聆听音乐增强跨皮层区域的功能信号活动,来研究不同脑皮层区域的时空有序相关动力学行为,尤其研究是否除1/f特征之外,还有其他对增强脑功能网络重要的因素.我们采用标准64通道EEG记录技术、独立分析变量和复杂网络方法分析了EEG数据,在实验中通过间隙和静息弱化了累积效应,在分析时通过截去每段音乐的前后2秒规避了过度效应.
在网络分析中,我们发现音乐信号被随机打乱后,大部分脑皮层区域的脑功能网络连接密度普遍下降(图2),并且增强的脑小世界网络特性CMean/LMean在一定阈值范围内也会有显著下降(图3,图4).虽然不能简单地认为其是网络组织中的结构变化,至少我们从功能连接层面印证了在1/f特征之外还有其他音乐信息,如音乐序列中的节拍数和节拍清晰度等信息会增强大脑网络的小世界特性.图7指出两首音乐在打乱前和打乱后每分钟节拍数和节拍清晰度出现了明显降低.这两种音乐特性的降低与音乐打乱前后的大脑小世界网络统计指标的CMean/LMean降低有显著关联.因此音乐作为输入信号,在以1s为单位进行全部碎片化打乱,即使保留了大于1Hz频率的1/f长程相关高级统计特性,但由于其他音乐有效信息的降低,这里尤其是节拍和节拍清晰度两个变量,人脑所感知的有效信息发生了变化,有可能也导致了人脑跨脑区小世界网络特性的降低.事实上,早期研究也指出[17-18],即使都是同种类型的音乐,钢琴演奏比起古琴演奏会更加增强受试者的小世界特性.因此音乐对人脑带来的影响可能是多个变量的贡献,值得更加深入的研究.当随机打乱之后,节拍等有效音乐信息降低,可能是导致脑功能连接密度下降、小世界特性降低的一个因素.大脑组织响应音乐的变化类似于工作记忆任务的表现[32].这种相似性表明,对具有丰富内涵信息的音乐信号收听任务可能会激活更高效的信息处理和认知能力,而失去节拍等有效信息的乱序音乐也在增强大脑网络小世界特性方面的能力会明显降低.本研究的另一目的是寻找在音乐片段之间的网络组织中可能的差异.我们的结果分析显示在音乐刺激之间脑网络特性没有发现差异,但这可能是所选音乐片段的风格相似的缘故.很有可能选择风格迥异的音乐会增强不同性质的小世界网络特性,值得我们将来设计实验进行深入研究.
总之,我们的研究提供了人类大脑功能网络在听正序与乱序音乐时重新配置的宏观证据.我们发现正序音乐认知活动引起皮层区域更能增强大脑跨皮层活动呈现小世界网络特性,说明皮层网络在大脑倾听音乐状态具有优化的功能连接特性,可能会有益于高效的认知信息传输.
[1] KANDEL E R, SCHWARTZ J H, JESSELL T M. Principles of neural science [M]. New York: McGraw-Hill, Health Professions Division, 2000.
[2] FAN L, LI H, ZHUO J,etal. The human brainnetome atlas: A new brain atlas based on connectional architecture [J].CerebralCortex, 2016,26(8) : 3508.
[3] PARK H-J, FRISTON K. Structural and functional brain networks: From connections to cognition [J].Science, 2013,342(6158): 1238411.
[4] MARMARELIS P Z, MARMARELIS V Z. Analysis of physiological systems: The white-noise approach [M].KluwerAcademicPub, 1978.
[5] VAN HATEREN J H. A theory of maximizing sensory information [J].BiolCybern, 1992,68(1): 23-29.
[6] RIEKE F, BODNAR D A, BIALEK W. Naturalistic stimuli increase the rate and efficiency of information transmission by primary auditory afferents [J].ProcBiolSci, 1995,262(1365): 259-265.
[7] LEWEN G D, BIALEK W, DE RUYTER VAN STEVENINCK R R. Neural coding of naturalistic motion stimuli [J].Network, 2001,12(3): 317-329.
[8] GARCIA-LAZARO J, AHMED B, SCHNUPP J. Tuning to natural stimulus dynamics in primary auditory cortex [J].CurrentBiology, 2006,16(3): 264-271.
[9] THEUNISSEN F E, DAVID S V, SINGH N C,etal. Estimating spatio-temporal receptive fields of auditory and visual neurons from their responses to natural stimuli [J].Network, 2001,12(3): 289-316.
[10] VOSS R F, CLARKE J. “1/fnoise” in music: Music from 1/fnoise [J].TheJournaloftheAcousticalSocietyofAmerica, 1978,63(1): 258-263.
[11] GILDEN D L, THORNTON T, MALLON M W. 1/fnoise in human cognition [J].Science, 1995,267(5205): 1837-1839.
[12] MUSHA T, YAMAMOTO M. 1/ffluctuations in biological systems [C]∥Proceedings of the 19th Annual International Conference of the IEEE. Chicago, USA: IEEE Press, 1997,6: 2692-2697.
[13] DE COENSEL B, BOTTELDOOREN D, DE MUER T. 1/fnoise in rural and urban soundscapes [J].Actaacusticaunitedwithacustica, 2003,89(2): 287-295.
[14] YU Y, ROMERO R, LEE T S. Preference of sensory neural coding for 1/fsignals [J].PhysRevLett, 2005,94(10): 108-103.
[15] SOBIE C, BABUL A, DE SOUSA R. Neuron dynamics in the presence of 1/fnoise [J].PhysicalReviewE, 2011,83(5): 051912.
[16] GAL A, MAROM S. Entrainment of the intrinsic dynamics of single isolated neurons by natural-like input [J].JournalofNeuroscience, 2013,33(18): 7912-7918.
[17] WU J, ZHANG J, DING X,etal. The effects of music on brain functional networks: A network analysis [J].Neuroscience, 2013,250: 49-59.
[18] WU J, ZHANG J, LIU C,etal. Graph theoretical analysis of EEG functional connectivity during music perception [J].BrainRes, 2012,1483: 71-81.
[19] WATTS D J, STROGATZ S H. Collective dynamics of ‘small-world’ networks [J].Nature, 1998,393(6684): 440-442.
[20] HONEY C J, SPORNS O, CAMMOUN L,etal. Predicting human resting-state functional connectivity from structural connectivity [J].ProcNatlAcadSciUSA, 2009, 106(6): 2035-2040.
[21] YU S, HUANG D, SINGER W,etal. A small world of neuronal synchrony [J].CerebralCortex, 2008,18(12): 2891-2901.
[22] CHEN Z, HE Y, YU Y. Enhanced functional connectivity properties of human brains during in-situ nature experience [J].PeerJ, 2016,4: e2210.
[23] MOGNON A, JOVICICH J, BRUZZONE L,etal. ADJUST: An automatic EEG artifact detector based on the joint use of spatial and temporal features [J].Psychophysiology, 2011,48(2): 229-240.
[24] GUEVARA M A, CORSI-CABRERA M. EEG coherence or EEG correlation? [J].IntJPsychophysiol, 1996,23(3): 145-153.
[25] RUBINOV M, SPORNS O. Complex network measures of brain connectivity: Uses and interpretations [J].Neuroimage, 2010,52(3): 1059-1069.
[26] FLOYD R W. ALGORITHM-97-SHORTEST PATH [J].CommunicationsoftheACM, 1962,5(6): 345-345.
[27] PATEL A D. Music, language, and the brain [M]. Oxford, UK: Oxford University Press, 2010.
[28] HOLE J, HIRSCH M, BALL E,etal. Music as an aid for postoperative recovery in adults: A systematic review and meta-analysis [J].TheLancet, 2015,386(10004): 1659-1671.
[29] GLASZIOU P. Music in hospital [J]. The Lancet, 2015, 386(10004): 1609-1610.
[30] PERETZ I, COLTHEART M. Modularity of music processing [J].NatNeurosci, 2003,6(7): 688-691.
[31] BASAR E, BASAR-EROGLU C, KARAKAS S,etal. Gamma, alpha, delta, and theta oscillations govern cognitive processes [J].IntJPsychophysiol, 2001,39(2-3): 241-248.
[32] ADRIAN R H, CHANDLER W K, HODGKIN A L. Voltage clamp experiments in striated muscle fibres [J].TheJournalofPhysiology, 1970,208(3): 607-644.
MusicEnhancesSmallWorldCharacteristicsofHumanBrainFunctionalNetwork
HEYujia,ZHANGWei,ZHENGGaoxing,YUYuguo
(1.TheCollaborativeInnovationCenterforBrainScience,DepartmentofPhysiologyandBiophysics,SchoolofLifeSciences,FudanUniversity,Shanghai200438,China;2.CenterforComputationalSystemsBiology,FudanUniversity,Shanghai200433,China)
In recent years, experiments have found that music compared with other mechanical noise on the human brain perception system can enhance a more complex network characteristics. Beautiful music generally have 1/fstatistical characteristics in the spectrum. Here we designed experiment to compare the human brain EEG response to normal music (NM) (has 1/fproperty) and randomly shuffled music sequences (RSMS) (1/fproperty is kept). In the music-induced EEG signal analysis, we found that the brain network displays a degraded small-world network property and lower brain functional connectivity density to RSMS signal than to NM. RSMS signal although retains long-term correlations, but the beat rate per min and clarity of the pulsation decrease. The degeneration of the two music parameter is correlated with the small network property, suggesting that the additional music parameters may contain rich information which also plays an important role in enhancing the brain function network.
music; EEG signal; 1/fstatistical characteristics; functional connectivity density; small world network characteristics
0427-7104(2017)06-0692-09
2017-03-07
国家自然科学基金(31571070),国家高级术研究发展计划(2015AA020508),上海市“东方学者”和国家癫痫精准医学(2016YFC0904400)项目
何裕嘉(1991—),男,硕士研究生;张 玮(1986—),男,科研助理;郑高兴(1994—),男,硕士研究生;于玉国,男,研究员,通信联系人;E-mail: yuyuguo@fudan.edu.cn.
Q42
A
