植物次生代谢产物-酚类化合物的研究进展
2018-01-08华晓雨陶爽孙盛楠郭娜阎秀峰蔺吉祥
华晓雨 陶爽 孙盛楠 郭娜 阎秀峰 蔺吉祥
(1. 东北林业大学盐碱地生物资源环境研究中心 东北盐碱植被恢复与重建教育部重点实验室,哈尔滨 150040;2. 扬州大学动物科技学院,扬州 225009;3. 黑龙江东方学院食品与环境工程学部,哈尔滨 150066)
植物次生代谢产物-酚类化合物的研究进展
华晓雨1陶爽1孙盛楠2郭娜3阎秀峰1蔺吉祥1
(1. 东北林业大学盐碱地生物资源环境研究中心 东北盐碱植被恢复与重建教育部重点实验室,哈尔滨 150040;2. 扬州大学动物科技学院,扬州 225009;3. 黑龙江东方学院食品与环境工程学部,哈尔滨 150066)
近年来,随着生物学理论与技术的快速发展,植物次生代谢产物愈发受到人们的关注,其中酚类物质是分布最广泛且具有重要生理功能的一类化合物,它不但具有天然的抗氧化活性,含多种药用价值,同时在植物生长发育以及逆境胁迫下的信号转导等多方面起着重要的调控作用,受到国内外学者的广泛重视。从酚类化合物的分类、合成途径、生理功能以及药用价值几方面对国内外相关研究进行了归纳与总结,并提出了今后该领域研究的方向及应该注意的问题,旨在为次生代谢产物的深入研究特别是酚类化合物的研究提供一定的参考。
酚类化合物;合成途径;生理功能;药用价值
植物的次生代谢是指由植物体内有机化合物的初生代谢途径衍生而来,最终合成一些具有种属特异性的有机化合物的代谢过程。在早期研究中,次生代谢及其产物曾被人们视为植物生命活动中无足轻重的机能与产物。但植物为何要耗费大量物质与能量来合成种类如此繁多的次生代谢产物,植物又为何在亿万年进化过程中保留了这一功能并演绎出如此多样化的次生代谢途径,这些问题随着生物学、医学、化学、生态学及食品工业科学等的发展逐渐被明确,人们也认识到植物的次生代谢产物可以为人类提供丰富的药物、香料和工业原料,而就植物本身而言,这些次生代谢产物与机体的防御、生长发育、信号转导等都有着紧密联系[1-2]。
酚类化合物是植物众多次生代谢产物中较为重要的一类化合物,它是指芳香烃中苯环上的氢原子被羟基取代所生成的一类化合物。酚类化合物一般都具有芳香气味,呈弱酸性,在环境中容易被氧化。越来越多的研究发现,酚类化合物显著影响植物与生态环境之间的关系,如酚酸、香豆素以及类黄酮对植物病原体有明显的抵抗作用[3],也有研究表明酚类化合物在蚕豆和根瘤菌之间的信号传导过程中起着关键的作用[4]。除此之外,酚类化合物具有的抗氧化活性对于防止食品氧化和促进人体健康也有着重要的作用[5]。因此,酚类化合物的研究也受到国内外学者的广泛关注。基于此,本文从酚类化合物的分类、合成途径、生理功能以及药用价值几方面对酚类化合物的相关研究进行了归纳与总结,以期为该领域的深入研究奠定一定的理论基础。
1 酚类化合物的分类
目前已经知道的酚类化合物多达8 000余种,并且在植物中有着广泛的分布。有报道称含酚类化合物较多的常见植物至少有600种以上。酚类化合物一般可以分为两大类:一类是多酚单体,即非聚合物,主要包括苯丙烷类和黄酮类化合物;另一类则是由单体聚合成低聚或多聚体,统称单宁类物质,包括缩合型单宁和水解型单宁[6]。
1.1 多酚单体
1.1.1 苯丙烷类 苯丙烷类化合物在植物界中普遍存在,对植物的生长调节和防御病毒的侵袭都有着重要作用。苯丙烷类化合物主要以羟基芳香环为共同特征,有多种不同的化学结构形式,它是以一个或多个C6-C3结构单元连在一起构成的天然产物[7]。根据其含有的C6-C3结构单元分子数又可以分为3类:香豆素类(含1分子C6-C3结构)、木脂素类(含2分子C6-C3结构)、木质素类(含多分子C6-C3结构)。香豆素是一种高活性化合物,具有化感作用,在植物与植物“通讯交流”过程中发挥着重要作用[8]。一般可以分为简单香豆素、呋喃香豆素、吡喃香豆素、异香豆素和其他香豆素。木脂素类化合物中比较常见的是四氢呋喃型木脂素,它是一类由2分子苯丙素衍生物通过侧链β碳原子聚合而成的且结构中具有C7-O-C7'、C9-O-C9'或C7-O-C9'等醚键形成的四氢呋喃环的木脂素[9]。木质素类化合物在植物体中占有很大的比重,仅次于纤维素,它以苯丙氨酸和酪氨酸为起点,经过相应的酸、醛,最后缩合而成,一般存在于裸子植物、被子植物和所有的维管植物中,蕴藏着丰富的化学能。
1.1.2 黄酮类 黄酮类化合物又名生物类黄酮,是自然界中广泛存在的一类重要化合物。黄酮类化合物具有复杂的化学结构,是以C6-C3-C6结构为基本母核,2个苯环通过中央三碳链相互连接成的一系列C6-C3-C6化合物[10]。根据核的不同模式可以将黄酮类化合物分为:黄烷醇类(儿茶素、表儿茶素等)、黄酮醇(类槲皮素、杨梅酮和山柰酚等)、花青素(花青色素、飞燕草色素、天竺葵色素、矢车菊色素和牵牛花色素等)、黄酮(芹黄素,地奥司明等)和黄烷酮类等[11]。黄酮类化合物具有如抗氧化性、抗肿瘤、调节酶活、杀菌消炎等生理功能,但由于其结构的复杂、不同的作用位点以及溶解度等问题,限制了其生理药理方面功效的发挥[12]。因此,近几年国内外学者一直在研究如何对黄酮类化合物的结构进行优化和改造,以增强其生物活性。
1.1.3 其他 除以上两种类型外,从结构上划分多酚单体还分为倍酸型、鞣花酸型(如鞣料云实素,番石榴素B等)、羟基芪类(如丹酚酸A等)、羟基萘蒽类(如掌叶二蒽酮等)[13]。
1.2 单宁类物质
单宁类化合物又称鞣质或鞣酸,是一类结构较复杂的多元酚化合物,广泛存在于植物中。其含量丰富,仅次于纤维素和木质素。单宁类化合物能与蛋白质形成不溶于水的沉淀,因此可与生兽皮的蛋白质形成致密的难透水的皮革。根据其化学结构的不同大致可以分为缩合单宁和水解单宁两种类型。落叶松、黑荆树和坚木的树皮中所含单宁就属于缩合单宁,其结构是由黄烷-3-醇为基本单位构成的。而水解单宁(如五倍子、橡碗单宁等)的结构主要是由倍酸及其衍生物与多元醇通过酯键形成的[14]。植物中由于含有单宁类物质而具有独特的理化活性,能与蛋白质、多糖、生物碱结合,使其理化性质发生改变,能与不同金属离子发生络合反应,捕捉自由基并且具有还原性[14]。
2 酚类化合物的合成途径与关键酶功能
植物中的酚类化合物具有多种合成途径,其中最主要的是莽草酸途径。该途径是酚类合成的中心,最初是经糖酵解及磷酸戊糖途径形成底物磷酸烯醇丙酮酸(Phosphoenolpyruvic acid,PEP)和4-磷酸赤藓糖(Erythrose 4-phosphate,E4P),通过 7-磷酸 -3-脱氧阿拉伯庚酮糖酸(3-Deoxy-D-arabinoheptulosonate 7-phosphate,DAHP)合成酶的催化后进入莽草酸途径,经过几种酶促反应过程形成了重要中间产物莽草酸,莽草酸与PEP相互作用,生成此途径重要的枢纽物质分支酸,最终生成色氨酸、苯丙氨酸和酪氨酸,进而合成一系列酚类化合物(图1)。
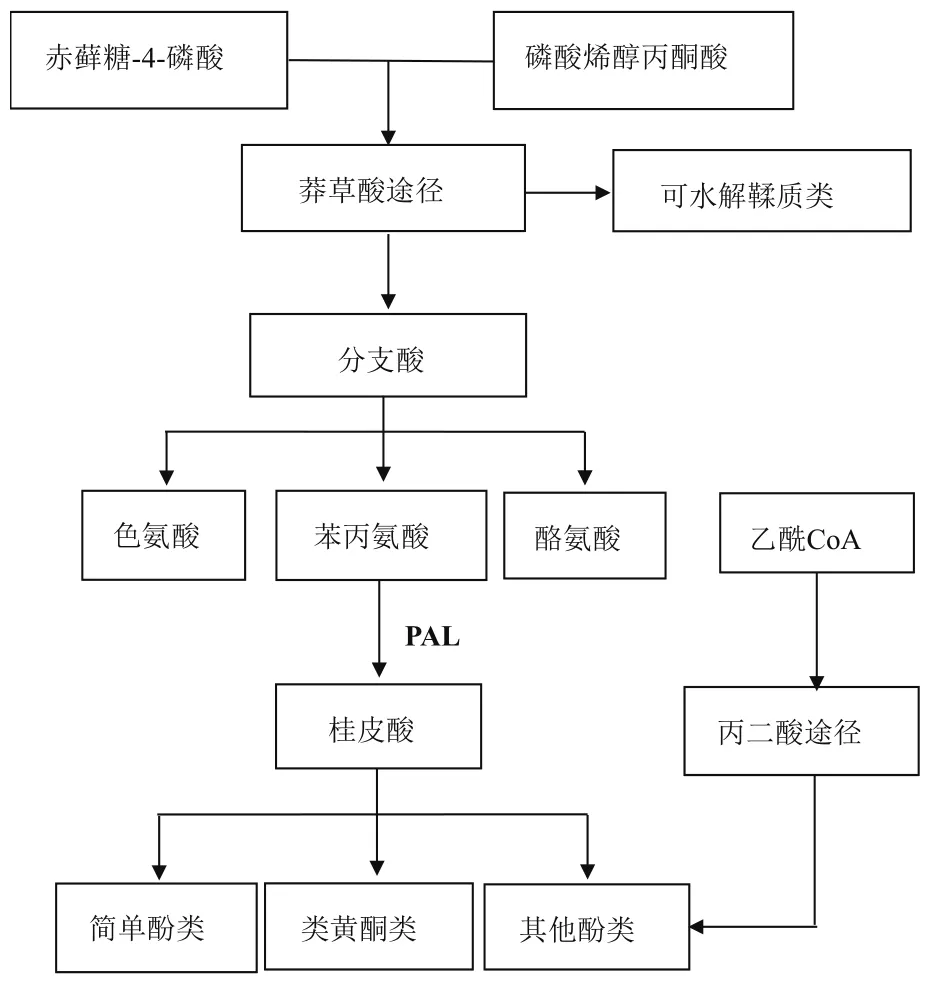
图1 酚类化合物的合成途径
在酚类化合物的合成代谢途径中有一些酶参与催化合成,其中苯丙氨酸解氨酶(Phenylalanineammonialyase,PAL)、肉桂酸羟化酶(Cinnamate 4-hydroxylase,C4H)、4-香豆酸-CoA连接酶(4-coumaryl:CoA ligase,4CL)、查尔酮合成酶(Chalcone synthase,CHS)为酚类化合物反应过程中的几种关键酶类,它们能够催化合成类黄酮、花青素、水杨酸和单宁等酚类化合物。类黄酮的生物合成途径是目前研究较为深入的一种,过程也较为清楚。首先,PAL作为植物生产酚类化合物的第一个关键酶,能够催化苯丙氨酸脱去氨基形成反式肉桂酸,再经C4H的作用下将肉桂酸与羟基催化形成P-香豆酸,然后P-香豆酸与辅酶A在4CL的作用下产生香豆酰-CoA,再进一步与丙二酰-CoA催化形成查尔酮,该产物进一步衍生构成一些类黄酮化合物[15]。
苯丙氨酸解氨酶作为植物次生代谢的关键酶,在植物的生长发育过程中起着重要的作用。该酶自首次在大麦幼苗中发现并进行分离纯化以来,一直备受关注[16]。人们对PAL提取并进行了活性测定,其中在植物生长逆境胁迫条件下,PAL常作为一个重要的响应指标[17]。有研究发现,许多植物在逆境胁迫条件下,苯丙烷类代谢被激活,使PAL活性迅速上升。如水稻幼苗在受到2.5 kJ·m-2和5.0 kJ·m-2UV-B辐射条件下,植物组织内苯丙氨酸解氨酶活性增加,促进类黄酮的合成,从而能够抵御外界胁迫[18],这也证实了PAL在次生代谢途径中发挥了重要的作用。
除了苯丙氨酸解氨酶外,查尔酮合成酶同样是植物次生代谢途径中的关键酶。它是植物黄酮类物质合成途径中的第一个酶。CHS能催化丙二酰-CoA和香豆酰-CoA结合形成查尔酮,之后进一步衍生转化成类黄酮化合物,这些化合物不仅能够赋予植物颜色,而且还参与了诸如抗病、生长素运输及抗逆境胁迫等多种生理过程[19]。目前,研究最多的是查尔酮合成酶分子生物学和基因工程方面,通过导入基因来提高植物中查尔酮合成酶的活性,利用调控途径从而改良物种抗逆等生理功能。有研究表明,结合TAIL-PCR技术从小麦叶片中克隆查尔酮合成酶基因,命名为TaCHS。以未侵染的小麦根部作为对照,实时定量PCR分析在全蚀菌侵染小麦根部的TaCHS的表达,结果显示经侵染的小麦根部中TaCHS基因表达开始上调,并在4 d时达到最大值,与未侵染的小麦根为对照相比有显著差异,这说明了TaCHS的表达可能对小麦的全蚀菌侵染有一定的抗性作用[20]。另有研究显示,高粱幼苗受玉米小斑病病菌侵染后,CHS的表达量明显提高,并且抗性高的品种表达持续时间更长,这也说明了CHS参与了高粱幼苗玉米小斑病的抗性作用[21]。
3 酚类化合物的生理功能
植物中的酚类化合物作为次生代谢产物在自然界普遍存在,并且具有广泛的生理功能。酚类化合物不仅能够作为植物器官的天然色素,还对植物的生长发育、逆境防御、信号转导以及抗氧化机理等方面有着重要的作用,这些生理功能对植物适应环境起着重要调节作用(图2)。
3.1 植物器官的天然色素
植物花会呈现不同颜色主要来源于花瓣细胞液中的花色素,其中最普遍的就是类黄酮物质,如花青素和黄酮醇。花青素是花和果实中的主要色素,植物呈现不同颜色是植物体内各种花青素、辅色剂以及液泡中的H+相互作用的结果[22]。薛德艳等[23]对钝裂银莲花花色素成分及其稳定性进行研究,结果表明,主要色素为黄酮类化合物(花色素苷、黄酮、黄酮醇和异黄酮)。此外,也有研究表明,花青素与辅色剂之间的相互作用可以使花青素更为稳定,辅色剂加入到花青素溶液中,可以使溶液的色泽加深[24]。黄酮醇合成酶(Flavonol synthase,FLS)可通过影响辅色剂的合成来改变花色,Holton等[25]从矮牵牛中克隆并过表达FLS,改变了花青素苷与黄酮醇的比例,导致矮牵牛花的颜色发生变化。另外,通过改变液泡pH可以使花青素苷颜色转变,有研究显示矮牵牛中PH1自身没有H+转运活性,但可以与PH5共同控制花瓣液泡的酸化,从而决定花朵的颜色,达到观赏目的[26]。鲜艳的花色和果实能够吸引昆虫和鸟类,有助于传粉和种子的传播,因此这也是酚类化合物生理功能的重要体现[27]。
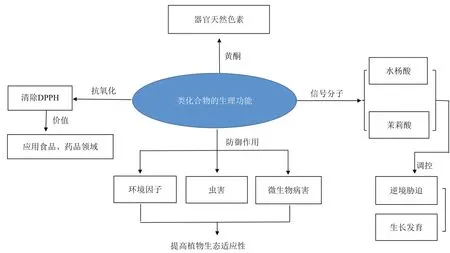
图2 酚类化合物的生理功能
3.2 防御作用
植物中的一些酚类化合物可对一些诸如侵染、辐射等逆境胁迫起到防御作用[28]。花青素能够有效的消除光合电子传递产生的活性氧自由基来减缓紫外光引起的损害,达到保护植物的作用[29]。有研究表明,将生长在高纬度地区的植物移植到低纬度地区,或者利用人工模拟的紫外辐射环境培养,发现植物叶片中类黄酮化合物的含量成倍增加[30]。上述研究均证明酚类化合物对紫外辐射有一定的防御作用。除此之外,大部分关于酚类物质与植物抗病性的相关报道中指出,植物在受到侵染时,会积累大量的酚类化合物,对病原菌起到抑制作用,从而达到抗病目的[31],其中木质素木质化可以为植物抵御病原菌的入侵。韩树全等[32]发现香蕉根部木质部导管中的穿孔板将其纵向连接的部分隔开,进一步阻止分生孢子的扩展使其形成堆积,阻止了分生孢子生长的菌丝体穿过孔板继续侵染,从而为抗病反应留出时间。冷平等[33]利用柿树枝条来检测黄酮类物质与植物抗寒性间的相互关联,结果表明,秋冬季节的低温胁迫造成活性氧在柿树枝条内的积累,而黄酮类物质上的羟基具有很强的供电子能力能够清除氧离子或其他自由基,从而控制低温逆境所产生的机体损伤。另外,植物中的单宁类物质也能够有效的抵御动物、微生物和病毒的伤害,它能够与蛋白质分子结合,是有效的化学防御物质[34]。有研究表明,缩合单宁作为一种慢性毒剂,抑制了棉铃虫幼虫的生长,起到抗虫病的作用,保护了棉花植株的生长繁殖[35]。上述研究都说明了植物中酚类化合物对外界逆境胁迫具有有较好的防御作用。
3.3 信号分子
酚类化合物同时还能够充当信号分子。近年来研究较多的水杨酸(Salicylic acid,SA)就是植物体内最常见的、有效的内源信号分子。它能够诱导植物抗旱、抗盐碱、耐低温等抗逆反应,同时也对植物的生长发育、成熟以及衰老等生理过程有显著的调控作用[36]。许多研究已经证实了水杨酸作为信号分子具有一定的生理功能。White等[37]利用外源SA作为信号分子来提高烟草对烟草花叶病毒的抗性作用,当烟草受到病毒侵染后,体内的SA合成急剧增加,激活产生抗性反应的细胞信号响应,从而提高烟草的抗性。近年来,又有研究发现,经浓度为5 mmol/L水杨酸喷雾处理后的黄瓜植株,其体内与抗病相关的酶,如苯丙氨酸解氨酶、多酚氧化酶(Polyphenol oxidase,PPO)、过氧化物酶(Peroxidase,POD)的活性迅速升高,对白粉病起到抑制作用[38]。另外,许多学者还研究了水杨酸介导病理相关蛋白,利用水杨酸类似物苯并噻二唑-S-甲基酯(Benzothiodiazole,BTH)筛选鉴定并克隆拟南芥抗病相关基因,揭示了拟南芥水杨酸通路蛋白参与植物抗生物胁迫的作用机理[39]。除水杨酸外,茉莉酸也是重要的信号分子。有研究表明,人参的根部含有三萜皂苷类化合物,而茉莉酸甲酯能够诱导鲨烯环氧酶和香树素合成酶的大量表达,进一步提高人参皂苷的含量[40]。另外,类黄酮可以作为根瘤菌结瘤基因的诱导物质,根瘤菌在类黄酮的作用下产生结瘤因子,通过Ca2+介导或磷酸类脂信号转导途径,诱导宿主植物的一系列反应[41]。
3.4 抗氧化作用机理
国内外学者经过大量的研究发现,酚类化合物具有很强的抗氧化机理,是很好的还原剂、金属离子络合剂、单线态氧猝死、灭剂和供氢体。它能够清除自由基、杀菌、抗癌、抑制脂质过氧化,还能缓解自动氧化链式反应的进行,是潜在的高效天然氧化剂[42]。范金波等[43]通过测定不同浓度的咖啡酸对1,1-二苯基-2-苯肼基(DPPH)自由基的清除能力及对咖啡酸的抗氧化性进行评价发现,当咖啡酸浓度在0-0.035 mg/mL时,DPPH的清除率呈线性增长,说明咖啡酸具有清除DPPH自由基的能力与较强的抗氧化活性。另有研究同样采用DPPH自由基清除法来测定山奈酚的抗氧化活性,结果表明,在山奈酚与羟乙基-β-环糊精的包合物浓度范围在0.5-2.5 mg/mL时,DPPH自由基的清除率随浓度升高而增强,这也证实了山奈酚具有一定的抗氧化活性[44]。Brown 等[45]研究了槲皮素与 Cu2+的相互作用,结果证明B环邻3’,4’二羟基能以氧负离子的形式与金属离子形成稳定的螯合物,从而影响了抗氧化活性。施伟梅等[46]利用超声协同酶法提取了紫花苜蓿多酚,证明其具有很好的抗氧化性,清除DPPH的半数抑制浓度IC50达到了10.78 g/mL。上述研究都证实了酚类化合物具有很强的抗氧化活性。酚类化合物的抗氧化活性也一直是研究的热点,并且作为抗氧化剂已经被广泛的应用到了食品、药品等领域。
4 酚类化合物的药用价值
酚类化合物具有抗心肌缺血、保肝、抗肿瘤、抗炎症等多种功效,人们将其广泛应用到医药研究中,不断发挥其药用价值。
4.1 抗心肌缺血作用
酚类化合物在抗心肌缺血方面有显著的药用价值。研究表明,天山花楸叶黄酮类物质能够通过灌注损伤后左心室(Left ventricular developed pressure,LVDP),以最大速率清除自由基,从而抑制脂质的过氧化反应来保护心肌细胞,达到改善人体心肌缺血的症状[47]。陈健康[48]等通过口服给药的方法观察银杏黄酮对家兔心肌缺血症状是否有所改善。结果表明,家兔左心室内压微分在心肌缺血和缺陷/再灌注过程中,与对照组相比,银杏黄酮组对家兔的心肌组织的梗死有明显的减缓作用,从而改善心肌缺血症状。此外,也有研究发现,以大鼠为实验对象,当淡竹叶总黄酮浓度为50和100 mg/kg时,能够明显抑制缺血/再灌注所致血清和心肌组织中LDH、CK活性的升高,同时增加心肌细胞以及血清中超氧化物歧化酶(Superoxide dismutase,SOD)和谷胱甘肽过氧化物酶(Glutathione peroxidase,GSH-PX)的活性,明显提高血清中NO含量,加速清除ROS,从而增强心肌细胞的抗氧化能力,抑制心肌脂质过氧化,达到保护心肌细胞的目的[49]。
4.2 抗肿瘤作用
除了抗心肌缺血作用,酚类化合物在抗肿瘤方面也有很高的药用价值。近年来,人们对其抗肿瘤作用进行了广泛的研究和应用。李辉敏等[50]在刺头复叶耳蕨总黄酮体外抗肿瘤活性的研究发现,体外培养的肝癌HepG2细胞在加入不同浓度的复叶耳蕨提取物和总黄酮,1 d后发现肝癌细胞生长出现抑制,2 d后多数细胞出现裂解、凋亡,进一步说明总黄酮能够抑制肝癌细胞HepG2的生长与扩散。除此之外,咖啡酸和其酯类物质,如咖啡酸苯乙酯也具有抗肿瘤活性,能抑制癌细胞的生长,咖啡酸苯乙酯(Caffeic acid phenethylester,CAPE) 作 为 NF-κB抑制剂,NF-κB参与调控一些肿瘤坏死因子受体相关因子,能够阻止Tax蛋白与IκBα结合,防止其降解,抑制了NF-κB的活性,从而阻断人类T淋巴细胞白血病病毒1型的致癌作用[51]。另有研究表明,木犀草素对人的胃癌细胞BGC-823的增殖起到抑制作用。通过在体外利用浓度为5、10、20和40 μmol/L的木犀草素对胃癌细胞BGC-823进行处理,发现在处理48 h后BGC-823细胞的生长受到明显抑制,且木犀草素的浓度越高,抑制效果越明显[52],这也进一步证实了木犀草素在体外具有较好的抗肿瘤活性。此外,苜蓿中的黄酮类化合物可以降低乳腺癌的发病率,黄酮中的刺芒柄花黄素能够降低甲状腺癌的发病率[53]。
4.3 抗炎症作用
酚类化合物利用自身的抗氧化活性,能够起到很好的抗炎作用。有研究表明,茶蜂花粉提取物(Bovine pituitary extract,BPE)中富含黄酮等酚类物质,平舜等[54]以BPE为材料,采用脂多糖(Lipopolysaccharides,LPS)诱导小鼠Raw254.7细胞体外炎症模型。结果显示,BPE能显著的抑制Raw264.7细胞NO的释放以及炎症相关基因的表达,这说明了黄酮类物质具有良好的抗炎症效果。有研究人员利用角叉菜胶导致大鼠足肿胀来创建一个急性炎症模型,用来测定人工培植雪莲中黄酮的抗炎性。结果表明,急性足肿胀大鼠外涂雪莲黄酮组与对照组相比,肿胀程度和肿胀率均呈现显著性降低,说明雪莲总黄酮具有明显的抗炎作用[55]。另外,AP-1是细胞内的一个重要转录激活因子,它能够应对多种刺激和细胞进程,包括控制炎症、细胞分化、发育的激活启动蛋白等作用。Maggi-Capeyron等[56]研究发现低浓度下用葡萄糖酚酸类化合物可以抑制AP-1的转录活性,这也间接说明了酚酸具有抑制炎症、抗菌的作用。
5 展望
具有代表性的次生代谢产物——酚类化合物,不仅在植物中具有重要的生理功能,还在医学、食品、化妆品及保健品等领域有着广泛的研发前景。纵观国内外对酚类化合物的分类、合成途径、生理功能及药用价值等领域的研究现状,以下几点需要进一步研究。
首先应该加大对植物资源中酚类化合物的研发与利用。随着分子生物学技术的不断发展,大量新的次生代谢产物被报道,特别对是一些生长在极端环境下的植物体内次生代谢物质的研究需进一步深入,而这一类植物中通常会含有一些酚类化合物且具有独特的生理功能。另外,随着基因组学、蛋白组学及代谢组学技术的更新,对各种类型植物中的酚类化合物的结构、代谢途径及基因调控模式等研究应进一步加强,应进一步明晰酚类化合物的生理功能,重点利用分子生物学手段从分子水平阐述重要酚类化合物活性成分合成过程中的关键酶基因,这也为药物研发等领域提供重要的科学依据。
其次,加大对酚类物质提取与利用效率的研发工作。次生代谢产物通常在植物体内的含量很少,因此一旦有可利用的资源如酚类化合物,如何高效利用则显得尤为重要。依托现有的溶剂提取法、超声提取法及微波辅助提取法等进一步优化,提高提取率并且利用高效液相色谱(HPLC)、液相色谱与质谱联用技术(LC-MS)、高效液相色谱电喷雾电离质谱-质谱(HPLC-ESI-MS/MS)及核磁共振技术(NMR)等对酚类化合物精准的鉴定工作需要不断探索,以求最大效率的认识与利用酚类化合物。
最后,基于酚类化合物强大的药理活性,加大其在药物研发领域的研究同样尤为重要。尽管随着人们的不断研究,一系列酚类化合物的药物与食品被开发利用,但由于结构复杂,对于具有重要活性的酚类化合物研究应深入挖掘具有显著生物活性的先导化合物,为药物开发提供重要的科学依据。
[1] Wink M. Secondary Metabolites, the Role in Plant Diversification of[J]. Encyclopedia of Evolutionary Biology, 2016 :1-9.
[2] Bahmani M, Golshahi H, Saki K, et al. Medicinal plants and secondary metabolites for diabetes mellitus control[J]. Asian Pacific Journal of Tropical Disease, 2014, 4(5):S687-S692.
[3] Maddox CE, Laur LM, Li T. Antibacterial activity of phenolic compounds against the phytopathogen Xylella fastidiosa[J].Current Microbiology, 2010, 60(1):53-58.
[4] Bekkara F, Jay M, Viricel MR, et al. Distribution of phenolic compounds within seed and seedlings of two Vicia faba cvs differing in their seed tannin content, and study of their seed and root phenolic exudations[J]. Plant & Soil, 1998, 203(1):27-36.
[5] 陈守江, 姜松. 果蔬中的酚类化合物及其抗氧化作用(综述)[J].安徽技术师范学院学报, 2003, 17(2):144-148.
[6] 左玉 . 多酚类化合物研究进展[J]. 粮食与油脂 , 2013(4):6-10.
[7] 穆红梅, 杜秀菊, 张秀省, 等. 植物MYB转录因子调控苯丙烷类生物合成研究[J]. 北方园艺, 2015(24):171-174.
[8] Lupini A, Araniti F, Sunseri F, et al. Coumarin interacts with auxin polar transport to modify root system architecture in Arabidopsis thaliana[J]. Plant Growth Regulation, 2014, 74(1):23-31.
[9] 孙彦君, 王雪, 陈辉, 等. 四氢呋喃型木脂素类化合物研究进展[J]. 中草药 , 2013, 44(21):3067-3079.
[10] 龙春, 高志强, 陈凤鸣, 等. 黄酮类化合物的结构-抗氧化活性关系研究进展[J]. 重庆高教研究, 2006, 5(2):13-17.
[11] Grassi D, Aggio A, Onori L, et al. Tea, flavonoids, and nitric oxidemediated vascular reactivity[J]. J Nutr, 2008, 138(8):1554S.
[12] 朱勍, 贾威阵, 章银军. 黄酮类化合物结构修饰研究进展[J].发酵科技通讯, 2016, 45(1):50-54.
[13] 陈迪华, 常琪, 斯建勇, 等. 天然多酚成分研究进展[J]. 现代药物与临床, 1997(1):9-15.
[14] 邹婷, 彭志远. 单宁在高分子材料中的应用研究进展[J]. 应用化工, 2015, 44(12):2308-2311.
[15] 夏涛, 高丽萍. 类黄酮及茶儿茶素生物合成途径及其调控研究进展[J]. 中国农业科学, 2009, 42(8):2899-2908.
[16] Koukol J, Conn EE. Metabolism of aromatic compounds in higher plants. IV. Purification and properties of phenylalanine deaminase of Horden vulgare[J]. J Biol Chem, 1961, 10:2692-2698.
[17] 高雪. 植物苯丙氨酸解氨酶研究进展[J]. 现代农业科技,2009(1):30-33.
[18] 姚银安, 祖艳群, 李元. 紫外线B辐射与植物体内酚类次生代谢的关系[J]. 植物生理学报, 2003, 39(2):179-184.
[19] 许锋, 程水源. 植物查尔酮合成酶分子生物学研究进展及展望[C]. 泰安:中国园艺学会青年学术讨论会. 2006.
[20] 薛保国, 谢丽华, 杨丽荣, 等. 小麦查尔酮合成酶基因及其启动子序列的克隆与分析[J]. 植物病理学报, 2017, 47(1):50-60.
[21] Cui Y, Frederiksen R, Magill C. Chalcone synthase and phenylalanine ammonia-lyase mRNA levels following exposure of sorghum seedlings to three fungal pathogens[J]. Physiological &Molecular Plant Pathology, 1996, 49(3):187-199.
[22] 顾林, 朱洪梅, 顾振新. 花青素的生物合成和成色机理及提高其稳定性的途径[J]. 食品工业科技, 2007(11):240-244.
[23] 薛德艳, 刘左军, 高望, 等. 钝裂银莲花花色素成分及其稳定性[J]. 草业科学 , 2015, 32(10):1569-1575.
[24] Bakowska A, Kucharska AZ, Oszmianski J. The effects of heating,UV irradiation, and storage on stability of the anthocyaninpolyphenol copigment complex[J]. Food Chemistry, 2003, 81(3):349-355.
[25] Holton TA, Brugliera F, Tanaka Y. Cloning and expression of flavonol synthase from Petunia hybrida[J]. Plant Journal for Cell& Molecular Biology, 1993, 4(6):1003-1010.
[26] Faraco M, Spelt C, Bliek M, et al. Hyperacidification of vacuoles by the combined action of two different P-ATPases in the tonoplast determines flower color[J]. Cell Reports, 2014, 6(1):32-43.
[27] Albert NW, Lewis DH, et al. Light-induced vegetative anthocyanin pigmentation in Petunia[J]. J Exp Bot, 2009, 60(7):2191.
[28] 王玲平, 周生茂, 戴丹丽, 等. 植物酚类物质研究进展[J].浙江农业学报, 2010, 22(5):696-701.
[29] 王良再, 胡彦波, 张会慧, 等. 植物叶片花青素的光破坏防御机制研究进展[J]. 应用生态学报, 2012, 23(3):835-841.
[30] 罗丽琼, 陈宗瑜, 丁金玲, 等. 低纬高原地区UV-B辐射对报春花类黄酮含量的影响[J]. 云南农业大学学报, 2007, 22(2):229-233.
[31] Boudet AM. Evolution and current status of research in phenolic compounds[J]. Phytochemistry, 2007, 68(22-24):2722-2735.
[32] 韩树全. 酚类物质抗香蕉枯萎病的作用机理[D]. 武汉:华中农业大学, 2014.
[33] 冷平, 张国军, 吴晓云, 等. 秋冬季节柿属植物树体内酚类物质含量的变化[J]. 中国农业大学学报, 2001, 6(1):63-67.
[34] 石蕾, 严善春, 金虎. 杂种落叶松子代针叶内单宁质量分数及防御性酶活性的变化[J]. 东北林业大学学报, 2013, 41(1):100-105.
[35] 张永军, 郭予元. 棉花缩合单宁和Bt杀虫蛋白的交互关系[J]. 棉花学报 , 2000, 12(6):294-297.
[36] Metwally A, Finkemeier I, Georgi M, et al. Salicylic acid alleviates the cadmium toxicity in barley seedlings[J]. Plant Physiology,2003, 132(1):272-281.
[37] White R F. Acetylsalicylic acid(aspirin)induces resistance to tobacco mosaic virus in tobacco[J]. Virology, 1979, 99(2):410-412.
[38] 岳东霞, 张要武, 庄勇, 等. 水杨酸对黄瓜植株抗病酶系和白粉病抗性的诱导作用[J]. 河北农业大学学报, 2003, 26(4):14-17.
[39] 朱杉. 拟南芥水杨酸通路蛋白参与植物抗生物胁迫的作用机理研究[D]. 兰州:兰州大学, 2012.
[40] 张琴, 张磊. 豆科植物根瘤菌结瘤因子的感知与信号转导[J].中国农学通报, 2005, 21(7):233-238.
[41] 任昂. 茉莉酸甲酯对灵芝三萜生物合成的影响及其灵芝应答基因的差异表达研究[D]. 南京:南京农业大学, 2012.
[42] Popa VI, Mariana D, Irina V, et al. Lignin and polyphenols as allelochemicals[J]. Industrial Crops & Products, 2008, 27(2):144-149.
[43] 范金波, 蔡茜彤, 冯叙桥, 等. 咖啡酸体外抗氧化活性的研究[J]. 中国食品学报, 2015, 15(3):65-73.
[44] 李云波, 陈玉珍, 马汉军. 山奈酚/HE-β-CD包合物的结构表征及其抗氧化活性研究[J]. 食品科技, 2016(5):253-257.
[45] Brown JE, Khodr H, Hider RC, et al. Structural dependence of flavonoid interactions with Cu2+ions:implications for their antioxidant properties[J]. Biochem J, 1998, 330(3):1173-1178.
[46] 施伟梅, 吴龙火, 杨翔, 等. 超声协同酶法提取紫花苜蓿多酚及其抗氧化性质[J]. 草业科学, 2016, 33(3):519-526.
[47] 耿玮峥, 吕铭洋, 崔新明, 等. 天山花楸叶总黄酮对心肌缺血再灌注损伤大鼠心肌酶和心肌超微结构的影响[J]. 中国实验诊断学, 2015(6):877-879.
[48] 陈健康, 卫张蕊, 刘水冰, 等. 银杏黄酮对急性心肌缺血/再灌注的药理作用[J]. 心脏杂志, 2007, 19(3):293-294.
[49] 邵莹, 吴启南, 周婧, 等. 淡竹叶黄酮对大鼠心肌缺血/再灌注损伤的保护作用[J]. 中国药理学通报, 2013, 29(2):241-247.
[50] 李辉敏, 姜登钊, 殷嫦嫦. 刺头复叶耳蕨总提取物与总黄酮体外抗肿瘤活性的比较研究[J]. 中国现代应用药学, 2015, 32(9):1037-1041.
[51] Shvarzbeyn J, Huleihel M. Effect of propolis and caffeic acid phenethyl ester(CAPE)on NFκB activation by HTLV-1 Tax[J]. Antiviral Research, 2011, 90(3):108-115.
[52] 陆雪莹, 李艳红, 肖向文, 等. 木犀草素抑制人胃癌BGC-823细胞增殖作用的研究[J]. 现代中西医结合杂志, 2012, 21(3):246-249.
[53] 左晓昕, 姚丹丹, 占今舜, 等. 苜蓿生物活性物质在动物上的应用[J]. 草业科学 , 2015, 32(7):1185-1191.
[54] 平舜, 王凯, 张江临, 等. 茶蜂花粉提取物BPE对LPS诱导的Raw264. 7细胞的体外抗炎症作用[J]. 食品与生物技术学报,2015, 34(12):1302-1307.
[55] 陈伟强, 杨泽民, 胡太平, 等. 人工培植雪莲黄酮含量测定及其抗炎作用的研究[J]. 中国实用医药, 2010, 5(4):12-14.
[56]Maggi-capeyron MF, Ceballos P, Cristol JP, et al. Wine phenolic antioxidants inhibit AP-1 transcriptional activity[J]. Journal of Agricultural and Food Chemistry, 2001, 49(11):5646-5652.
Research Progress on Phenolic Compounds of Plant Secondary Metabolites
HUA Xiao-yu1TAO Shuang1SUN Sheng-nan2GUO Na3YAN Xiu-feng1LIN Ji-xiang1
(1. Alkali Soil Natural Environmental Science Center,Northeast Forestry University,Key Laboratory of Saline-alkali Vegetation Ecology Restoration,Ministry of Education,Harbin 150040 ;2. Colleges of Animal Science and Technology,Yangzhou University,Yangzhou 225009;3. Department of Food and Environmental Engineering,East University of Heilongjiang,Harbin 150066)
With the rapid development of biological theories and technologies in recent years,plant secondary metabolites are growingly concerned. Among them,the phenolic compounds are the most widely distributed and have critical physiological functions. They not only have natural antioxidant activity,but also contain a variety of medicinal values,and play an important regulating role in plant development and signal transduction under stress conditions,thus have become a hot spot of research. Based on this,we summarized the classification,synthesis pathways,physiological functions and medicinal values of the phenolic compounds from domestic and foreign researches,and put forward the future research direction and the problems that should be paid attentions to,aimed at providing some references for the in-depth study of secondary metabolites,especially phenolic compounds.
phenolic compounds;synthetic pathway;physiological function;medicinal value
10.13560/j.cnki.biotech.bull.1985.2017-0546
2017-07-01
国家自然科学基金项目(31502013)
华晓雨,女,硕士研究生,研究方向:植物生理生态学;E-mail:huaxiaoyu0828@126.com
蔺吉祥,男,副教授,研究方向:植物逆境生理生态学;E-mail:linjixiang@nefu.edu.cn
(责任编辑 朱琳峰)
