江苏宝华山红柴枝群落组成结构及多样性分析
2018-01-06张开文谢梦梦王鹏程李韦鸿伊贤贵
从 睿,张开文,谢梦梦,王鹏程,李韦鸿,伊贤贵*
(1.南方现代林业协同创新中心,南京林业大学生物与环境学院,南京 210037;2.深圳梧桐山风景管理处,广东深圳 518004)
江苏宝华山红柴枝群落组成结构及多样性分析
从 睿1,张开文2,谢梦梦1,王鹏程1,李韦鸿1,伊贤贵1*
(1.南方现代林业协同创新中心,南京林业大学生物与环境学院,南京 210037;2.深圳梧桐山风景管理处,广东深圳 518004)
【目的】为了更好地保护、开发和利用宝华山红柴枝这一珍稀植物资源,掌握其所处群落的特征,使该资源实现可持续化。【方法】对宝华山红柴枝植物群落样地调查和群落分析。【结果】在所调查的4 000 m2样地中,红柴枝植物群落中蕨类植物和种子植物共有70科135属162种;群落种类以榆科、蔷薇科、豆科、菊科和禾本科等为主。种子植物共64科129属156种,其中科有6种分布类型,热带分布科占65.79%;属有13种分布类型,北温带分布属占31.53%。重要值分布表明毛竹、红柴枝、紫楠、枫香树等为该群落的优势种群,其值分别为11.77%、8.24%、7.96%和7.35%。种群年龄结构的立木级分析表明,荚蒾、山胡椒、红柴枝等属于增长种群,而枫香树、朴树等属于衰退种群。在垂直结构上,草本层多样性最大、灌木层次之、乔木层则最小。【结论】红柴枝在群落中是较稳定的种群,低龄级苗木(Ⅰ级和Ⅱ级)较多,但该群落内种间竞争力强、郁闭度较高、受人为干扰等因素的影响,红柴枝生长受限,需适当开窗疏林。由于其优良性状并未进一步得到利用,因此,在加强保护红柴枝的基础上,可对其进行合理开发利用。
宝华山;红柴枝;群落组成;多样性
宝华山为国家级森林公园,森林类型为典型的常绿阔叶林、常绿-落叶阔叶混交林、落叶阔叶林及人工林演替的竹阔混交林[1],区内植被丰富、物种多样性高,先后有宝华玉兰(Magnolia zenii)[2]、野生早樱(Cerasus subhirtella var.ascendens)[3]、紫楠(Phoebe sheareri)[4]等群落学报道,前人对该地区具有代表性的植物群落结构及物种多样性研究进行了深入分析。
红柴枝(Meliosma oldhamii Maxim.)隶属清风藤科(Sabiaceae)泡花树属(Meliosma),又名南京珂楠树,主要分布于贵州、江西、浙江、江苏等地300~400 m之间的阳坡山谷、山坡、山地疏林中,或天然落叶阔叶林边缘地带[5-6]。该树树姿优美、花色淡雅、果实绚丽,极具观赏价值;木材坚硬,种子可制油,根皮茎叶可入药[7],极具经济、药用价值;加之其适应性与抗性强,在群落中具有一定的竞争力,因此开发利用前景广阔[8-9]。红柴枝已被列为江苏省珍稀保护植物之一[10],与同是省级保护植物的柞木(Xylosma racemosum)[11]、四照花(Dendrobenthamia japonica var.chinensis)[12]、南京椴(Tilia miqueliana)[13]等相比,上述物种均有群落学及相关性报道,而有关红柴枝的研究甚少,尤其是群落结构特征至今尚未见报道。宝华山自然分布的红柴枝小居群,受各种因素影响,使该种群生存面临一定的威胁。本研究拟通过对江苏宝华山红柴枝群落的调查,了解其所在群落的基本特征,分析资源状况,同时为开展红柴枝的保护、引种、驯化、繁殖及推广应用等提供科学理论依据。
1 材料和方法
1.1 研究区自然概况
宝华山位于 N32°06'~32°10',E119°03′~119°08′,最高峰437.2 m,总面积17.67 km2。该地区属北亚热带湿润季风气候,四季分明,气候温和,雨水充沛,日照充足;年均温度15.4℃,年均降水量为1 018.6 mm;土壤为棕壤土,呈微酸性,土层较为深厚[14]。宝华山曾是北亚热带生物资源最为丰富的地区之一[15-16],拥有大量观赏价值高的野生植物资源[17],形成和保留了许多中国具地域代表性的物种和珍贵的古树名木[18]。宝华山现状植被类型主要是常绿落叶阔叶混交林和落叶阔叶林,其中优势种或群落建群种主要有朴树(Celtis sinensis)、建始槭(Acer henryi)、枫香树(Liquidambar formosana)、青冈(Cyclobalanopsis glauca)和紫楠等;同时还有一些油茶(Camelliaoleifera)林、黑松(Pinus thunbergii)林、毛竹(Phyllostachys edulis)林等人工植被。
1.2 研究方法
1.2.1 样地调查
在江苏宝华山红柴枝自然分布区内,对物种保存相对较好、代表性较强的地段设定10个20 m×20 m的样方,总面积4 000 m2。调查方法为“每木记账调查方法”。乔木起测高度为1.5 m,起测胸径为1.0 cm;乔木层测量胸径、高度、株数,灌木层测量高度、株数;草本层测量高度、盖度[19-20]。在每个所测样地内分别随机选择5 m×5 m的区域用来统计灌木、草本和层间植物的数据,总面积400 m2。
1.2.2 数据分析
①群落物种组成与区系分析[21-22]:统计样方内所有维管植物科、属及种的数量和比例;同时根据吴征镒对我国种子植物科、属分布区类型的划分分析该群落植物区系成分。
②重要值分析[23-24]:乔木层重要值(IV1)=(相对密度+相对优势度+相对频度)/3,灌木层重要值(IV2)=(相对密度相对高度+相对频度)/3,草本层重要值(IV3)=(相对盖度+相对高度+相对频度)/3。
③立木结构划分[25]:将重要值排前10位的乔木树种以立木级代替年龄级进行划分,设4个立木级(用DBH表示胸径):Ⅰ级苗木DBH<2.5 cm;Ⅱ级小树2.5cm≥DBH<7.5cm;Ⅲ级壮树7.5 cm≥DBH<22.5 cm;Ⅳ级大树DBH≥22.5 cm。
④物种多样性指数分析[26]:选用Shannon-Wiener多样性指数(S)、Pielou均匀度指数(E)和丰富度指数(R)来讨论群落的物种多样性特征。
2 结果分析
2.1 群落物种组成与区系分析
2.1.1 群落物种组成
经调查统计,样地内共有维管植物70科135属162种,其中蕨类植物6科6属6种,种子植物64科129属156种,无裸子植物。红柴枝群落以双子叶植物占优势,裸子植物缺失。
在样方中,种数为10种以上的仅有蔷薇科(Rosaceae),占所有种比例的6.79%,多数是悬钩子属(Rubus)植物。含6~10种的科主要有壳斗科(Fagaceae)和榆科(Ulmaceae),这两个科的植物构成了乔灌层的主要物种;豆科(Leguminosae)、菊科(Compositae)和禾本科(Gramineae)占据了主要草本层。含2~5种的科较多,物种数占所有种的39.51%,多为胡桃科(Juglandaceae)、毛茛科(Ranunculaceae)、十字花科(Cruciferae)等植物。另外,群落中寡属种比例较大,以草本植物居多,一方面说明了群落物种组成的分散性较高,物种丰富度较大;另一方面也反映了红柴枝群落具有较强的次生性。
2.1.2 群落区系分析
根据吴征镒对中国种子植物科分布类型的划分(表1),该群落的64科分属于6个分布类型;其中世界分布科有26个,反映出植被的次生性强的特点。热带性质分布的科占主导地位,其中泛热带分布占比为50.00%。从热带成分、温带成分和中国特有成分3个植物分布成分看,样地内热带分布比例最大,为65.79%,温带分布次之,占比34.21%,无中国特有科。热带分布科于温带分布科比例接近2∶1,说明在科层次的区系性质上真正的热带性质并不明显。
从属的层次上看[27],群落129属中,世界分布属18个,常见广泛分布属有蓼属(Polygonum)、毛茛属(Ranunculus)等。从地理成分看,热带分布属共有36个,占总属数的32.43%;其中泛热带分布属最多有21属,占总属数的18.92%,如千金藤属(Stephania)、细辛属(Asarum)等。温带分布属共有69个,占总属数的62.16%,其中北温带分布属最多为35属,占总属数的 31.53%,主要有胡桃属(Juglans)、桑属(Morus)等。中国特有分布属有6个,占5.41%,分别为青檀属(Pteroceltis)、牛鼻栓属(Fortunearia)、山拐枣属(Poliothyrsis)、明党参属(Changium)、秤锤树属(Sinojackia)、短穗竹属(Brachystachyum)。
属比科更能反映出该群落植物区系特点。通过对红柴枝群落植物科属的地理成分分析,温带分布属与热带分布属的比例约为3∶2,一方面说明该地区温带植物成分占有较大的优势,群落性质为温带分布;另一方面一定热带区系成分的存在也说明了该群落植物区系具有亚热带到北温带过渡的特点[13]。
2.2 重要值分析
红柴枝群落中共有乔木60种,表2中仅列出重要值大于2.00%的13种优势种;这些物种的相对密度、相对优势度和相对频度之和分别79.49%、77.49%、50.86%,占总重要值的近70%,因此能够代表该群落的主要特征。
由表2可见,除了毛竹和紫楠以外,红柴枝相对密度、相对优势度和相对频度均具有最大值,这说明红柴枝在群落内分布较均匀、幼龄树种较多、环境适合其繁殖生长,是群落内仅次于毛竹的主要建群种之一。群落中毛竹重要值最大为11.77%,在该群落中处于较大的优势地位,占据较大的生存空间。但是,从目前群落的演化趋势来看,毛竹极有可能取代红柴枝、紫楠、枫香树等优势种成为该群落的绝对优势种,而其他树种也会因为没有足够的资源丧失种间竞争力,从而逐渐退出群落的竞争。因此,急需对毛竹进行适当疏理和砍伐利用,否则群落内的植物多样性会受到一定的影响,尤其是一些幼小的乔灌木。毛竹、紫楠、油茶和青冈等常绿树种占有更多的光热和水分资源,由此导致落叶树种在这方面的劣势,这在一定程度上制约了红柴枝的生长,尤其是对幼株生长的影响。
群落中常见灌木树种(重要值大于2.00%)有17种,它们的相对密度、相对高度、相对频度之和分别为63.37%、61.03%、53.68%,重要值之和近60%,这些物种基本能够代表群落灌木层的主要结构特征。优势物种中建始槭、紫楠、化香树(Platycarya strobilacea)等为乔木树种,在将来可能会增加其乔木的重要值,与红柴枝形成一定的竞争局面。
红柴枝群落中常见草本植物(重要值大于2.00%)有17种,它们的相对高度、相对盖度、相对频度之和分别为51.74%、57.89%、43.37%,重要值之和为50.99%。优势物种中百合科(Liliaceae)、蓼科(Polygonaceae)及禾本科的重要值占了一半,说明这几个科在草本层中占较大的优势;而寡属种、单属种的物种虽然在科属比例上占一定优势,却在数量上不占优势。
2.3 优势种群年龄结构分析
种群的年龄结构可以反映出种群数量的动态变化,同时也反映了群落发展的变化趋势。根据立木级模型可将样地内物种的年龄结构分为2种类型(表 3):1)金字塔型(稳定或增长种群),包括荚蒾(Viburnum dilatatum)、山胡椒(Lindera glauca)等;2)倒金字塔型或纺锤型(衰退或不稳定种群),包括枫香树、朴树等。群落内既有落叶优势树种,如枫香树、朴树等,又有常绿优势树种,如紫楠、青冈、油茶等。由此可推断该群落是在落叶树种基础上发育而来的常绿-落叶阔叶混交林,如果外界干扰较少,加之紫楠、油茶、青冈等常绿树种的生长,该群落将进一步向北亚热带常绿落叶阔叶混交林发展。此外,群落中不同物种各优势种群的年龄结构变化不一,反映出群落演替的不稳定性,造成红柴枝生长环境的复杂性更高。
红柴枝种群年龄结构基本接近金字塔型(Ⅰ、Ⅱ龄级树较多),年长的红柴枝得到更多的光热与水分,结实率增加,后代量大,所以种群中有一定的新生幼树和幼苗;成年树数量也占有一定比例,说明其种群具有增长性。但由于群落朝着常绿落叶阔叶混交林发展,次生林林分郁闭度增加,使得红柴枝所接受光热和水分的时间与空间比例减少,它与其他物种之间的相对竞争力会逐渐减弱。需要注意的是,红柴枝幼苗由于受光热及人为干扰等因素的存在,种群数量过小,后期更应注意其幼苗种群的管理与维护。
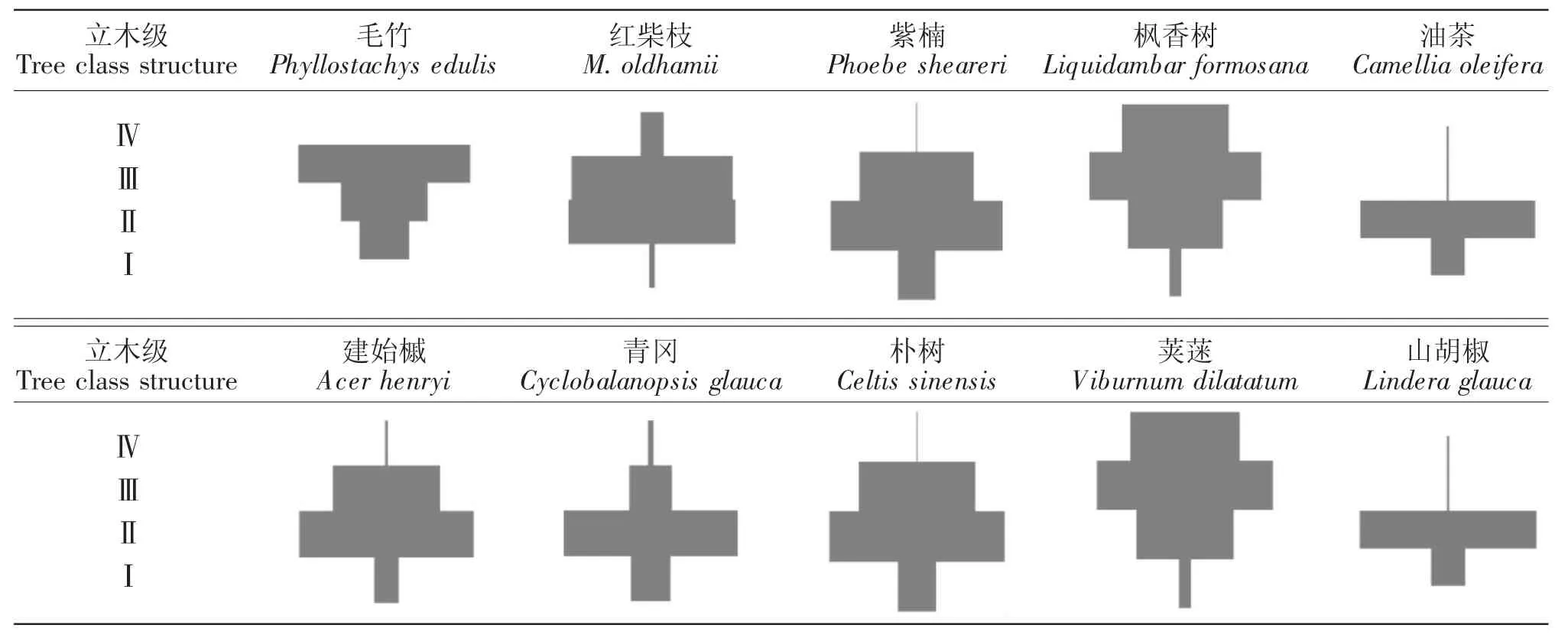
表3 宝华山红柴枝群落优势种群立木径级Table 3 Structure diagrams of standing trees in the Meliosma oldhamii communities in Baohua Mountain
2.4 群落物种多样性分析
由图1可知,随着乔木层、灌木层、草本层物种数量增多,多样性指数逐渐增大,丰富度指数随之减小,而均匀度指数则无明显变化关系;总体看来,物种数量越多,多样性指数和均匀度指数越大,丰富度指数越小。由此可知,群落物种数量决定其多样性,多样性指数越大,丰富度指数越小,则群落的复杂程度越高,群落中所含的信息量越大。
从数据中可以看出,乔木植物多样性以样方6最高,样方3次之;灌木植物多样性以样方9最高,样方6次之;草本植物多样性以样方5最高,样方3次之。各样方表现出不同的多样性差异,说明该群落生境的差异性是影响多样性的一个重要因素。
3 讨论与结论
宝华山红柴枝群落地处北亚热带北缘,是典型的落叶阔叶林。群落中蔷薇科、壳斗科、榆科植物在数量上占明显优势,其他科成分则较为简单。由于地处亚热带和暖温带过渡地带,受地理成分和气候因素的影响,该区系特征表现为热带性分布和温带性分布兼具的特点,但后者大于前者,说明该区系有北亚热带向温带过渡的特征,较好地反映出红柴枝群落种子植物的区系特征。由于群落中物种分布不均匀,群落存在分化与演替的趋势。就目前空间格局看,群落中红柴枝、枫香树等高大乔木占据优势层,未来一段时间群落仍趋于稳定。从垂直结构上来看,乔、灌、草层分层明显,高大乔木占据顶层优质空间,矮灌及其他幼苗分散林下。各层多样性表现为草本层最大、灌木层次之、乔木层最小。

图1 宝华山红柴枝群落物种数量、物种多样性、物种均匀度、物种丰富度特征Figure 1 The characteristics of number, diversity,evenness and richness of species in the Meliosma oldhamii community in Baohua Mountain
红柴枝在群落中属于的种群结构近金字塔型(将幼树幼苗合计考虑),虽有一定的小树存在,但新生苗木较少。但该群落郁闭度较大,存在很多常绿物种,如果适当开窗疏林,则有利于红柴枝种群增长。另外由于毛竹的入侵,其种间竞争具有较大的优势,在群落演替中极有可能发展为毛竹林。毛竹高大且常绿,林分郁闭度很高,竹鞭纵横交错,对其他树种的吸收养分与扎根支柱产生一定的遏制作用,一定程度上毛竹已经成为该群落物种多样性的破坏者。为保护红柴枝的生境,需适当控制毛竹的生长速度,例如可砍伐一些壮年毛竹用、挖取部分笋苗、设置适当隔离带等,从而减弱对红柴枝的抑制干扰。此外,由于人为干扰及自然环境的破坏,使得现存的红柴枝种群不断变小,如不加以重视保护,该种群则有可能不断减小甚至消失的可能。因此,可以采取以下策略:①强化保护树种的管理制度,增强森林巡护,防止乱砍滥伐等破坏林分行为。②加强森林抚育,对林内枯树、枯枝等进行清理扫除,开窗疏林,增强光照。③进行种质保护,研究其生态学和生物学特性[28-29]。④建立生态保护红线,对红柴枝重点监控,合理地对其进行引种驯化、人工栽培等加强种苗培育的工作,并引导其相关研究进展。加强开发利用与投入实际应用,是目前亟待解决的问题。
[1]张明娟,刘茂松,王磊,等.宝华山典型群落物种多样性的差异分析[J].南京林业大学学报(自然科学版),2003,27(6):35-39.
[2]王剑伟,张光富,陈会艳.特有珍稀植物宝华玉兰种群分布格局和群落特征[J].广西植物,2008,28(4):489-494.
[3]谢春平,张金鹤,王贤荣,等.武夷山和宝华山野生早樱群落植物区系研究[J].长江大学学报(自然科学版),2007,4(4):81-85,108.
[4]邓送求,闫家峰,关庆伟.宝华山紫楠风景林林分空间结构分析[J].东北林业大学学报,2010,38(4):29-32.
[5]凌云.宝华山国家森林公园植物区系及其生物多样性流研究[D].南京:南京师范大学,2011.
[6]张帆航.句容宝华山国家森林公园维管植物区系研究[D].南京:南京农业大学,2008.
[7]ABE F,YAMAUCHI T,SHIBUYA H.Triterpenoid Glycosides from Bark of Meliosma lanceolata[J].Phytochemistry,1996,42(3):809-814.
[8]刘易蓉,梁光义,张永萍.清风藤属药用植物的研究概况[J].贵阳中医学院学报,2006,28(1):50-52.
[9]赵洁,姚默,贾旭,等.清风藤属药学研究进展[J].安徽农业科学,2011,39(32):19804-19805.
[10]陈瑞冰,张光富,刘娟,等.江苏宝华山国家森林公园珍稀植物的濒危等级及优先保护[J].生态与农村环境学报,2015,31(2):174-179.
[11]陈祥伟,丁宝勇,石福臣,等.天然柞木林不同群落结构林木生理指标与小气候因子的关系[J].东北林业大学学报,1990,18(S3):47-55.
[12]易咏梅,艾训儒,彭诚,等.四照花群落结构及多样性研究[J].湖北民族学院学报:自然科学版,2010,28(2):205-207,229.
[13]汤诗杰,彭志,汤庚国.宝华山南京椴群落的特征分析[J].扬州大学学报:农业与生命科学版,2008,29(1):90-94.
[14]李玉双,陈洁,庞莉莉,等.宝华山常绿落叶阔叶混交林下蕨类植物研究[J].生态科学,2016,35(1):10-16.
[15]张月新.句容宝华山自然保护区简介[J].江苏林业科技,1984,1(1):58.
[16]宋红涛,郭水良,沈蕾,等.江苏宝华山自然保护区苔藓植物多样性及其分布与环境关系研究[J].山地农业生物学报,2010,29(6):482-489.
[17]邱国金,杨士虎.宝华山野生观花观果树木资源的开发利用[J].中国林副特产,2004,2(2):57,59.
[18]张明,唐明茂,杜晓祥.句容宝华山古树名木资源调查[J].江苏林业科技,2005,32(2):28-29.
[19]方精云,王襄平,沈泽昊,等.植物群落清查的主要内容、方法和技术规范[J].生物多样性,2009,17(6):533-548.
[20]黄健.南京老山地区植物区系与植物资源研究[D].南京:南京林业大学,2013.
[21]吴征镒,周浙昆,李德铢,等.世界种子植物科的分布区类型系统[J].云南植物研究,2003,25(3):245-257.
[22]吴征镒.《世界种子植物科的分布区类型系统》的修订[J].云南植物研究,2003,25(3):535-538.
[23]谢春平,王华辰,陈林,等.木林子自然保护区河谷樱花群落特征分析[J].南京林业大学(自然科学版),2014,38(S1):81-88.
[24]王育松,上官铁梁.关于重要值计算方法的若干问题[J].山西大学学报(自然科学版),2010,33(2):312-316.
[25]谢春平,方彦,伊贤贵,等.宝华山野生早樱群落特征的初步研究[J].广东农业科学,2011,38(3):56-59.
[26]王凤娟,丁福波.植物群落物种多样性研究综述[J].现代园艺,2015(4):155.
[27]李锡文.中国种子植物区系统计分析[J].云南植物研究,1996,18(4):363-384.
[28]RONSE DE CRAENE L P,WANNTORP L.Morphology and Anatomy of the Flower of Meliosma(Sabiaceae): Implications for Pollination Biology[J].Plant Systematics and Evolution,2007,271(1):79-91.
[29]WANNTORP L,RONSE DE CRAENE L P.Flower Development of Meliosma(Sabiaceae): Evidence for Multiple Origins of Pentamery in the Eudicots[J].American Journal of Botany,2007,94(11):1828-1836.
Community Composition Structure and Diversity of Meliosma oldhamii Maxim.in the Baohua Mountain of Jiangsu Province
CONG Rui1,ZHANG Kai-wen2,XIE Meng-meng1,WANG Peng-cheng1,LI Wei-hong1,YI Xian-gui1*
(1.Co-Innovation Center for Sustainable Forestry in Southern China,College of Biology and the Environment,Nanjing Forestry University,Nanjing 210037,China;2.Administration Division of Wutongshan National Park,Shenzhen 518040,Guangdong,China)
【Objective】In order to protect,exploit and utilize Meliosma oldhamii better,one of valuable plant resources in the Baohua Mountain of Jiangsu province,we investigated its community characteristic and geographical distribution and put forward some protection measures in this study.【Method】Field plot of M.oldhamii community was investigated and analyzed in theBaohua Mountain.【Results】There were 162 species(varieties),which belonged to 135 genera and 70 families in the 4 000 m2quadrats.The major families were Ulmaceae,Rosaceae,Leguminosae,Composite and Gramineae.For spermatophyte,there were 156 species,belonging to 64 families and 129 genera.There were 6 family types and 65.79%of them was distributed in tropical zone.There were 13 genera areal-types and north temperate distribution accounted for 31.53%of total type.P.edulis,M.oldhamii,P.sheareri and L.formosana were the dominant populations in the community,whose important values were 11.77%,8.24%,7.96%and 7.35%,respectively.The analysis of population age structure indicated that V.dilatatum,L.glauca and M.oldhamii were rising in the communitiy,while L.Formosana and C.sinensis were descending.For the vertical structure,the diversity of the herb layer was the largest,followed by the shrub layer and tree layer.【Conclusion】M.oldhamii was a stable populatioin in the communityand which mainly consist of young seedlings(Ⅰ andⅡclasses).Theseseedlingscouldnotgrowwellduetospeciescompetition,canopydensity and human disturbances.M.oldhamii can be exploited and utilized based on sustainable protection.
Baohua Mountain;Meliosma oldhamii Maxim.;community composition;diversity
S718.54+2;Q949.755.6;Q178.1+4 文献标志码:A 文章编号:1000-2650(2017)02-0213-07
10.16036/j.issn.1000-2650.2017.02.012
2016-11-10
绿色江苏专项资金(2130205);第二次全国重点保护野生植物资源调查专项资金。
从睿,硕士研究生。*责任作者:伊贤贵,讲师,博士研究生,主要从事樱属和木犀属植物研究,E-mail:354067272@qq.com。
(本文审稿:谢春平;责任编辑:巩艳红;英文编辑:徐振锋)
