赣南柑橘木虱体内黄龙病菌psy62株系原噬菌体类型多样性的研究
2018-01-04陈丽芬徐昭焕肖丽芳张丽卿王建国
陈丽芬,徐昭焕,肖丽芳,张丽卿,王建国*
(1.江西农业大学农学院, 南昌330045;2.崇义县农业与粮食局,江西崇义 341300)
赣南柑橘木虱体内黄龙病菌psy62株系原噬菌体类型多样性的研究
陈丽芬1,2,徐昭焕1*,肖丽芳1,张丽卿1,王建国1**
(1.江西农业大学农学院, 南昌330045;2.崇义县农业与粮食局,江西崇义 341300)
为探明柑橘木虱体内黄龙病菌psy62株系中原噬菌体遗传多样性及与柑橘株系的遗传差异,本研究将利用3对引物在psy62基因组中2个噬菌体FP1和FP2的同源序列区域内对赣南地区阳性样品进行PCR扩增和测序。结果显示,赣南地区阳性样品在此基因区域内有28头柑橘木虱检测到原噬菌体,共有5种类型,分别为A, A2, B, C, C1。其中A,B的检测率相对较高;C1检测率最低,为7.1%。由此可见,在此基因位点不同地理位置的柑橘木虱原噬菌体种群有较大的遗传差异,且与美国佛罗里达的“Ca.L. asiaticus”株系,中国柑橘叶片“Ca.L. asiaticus”株系均有显著的遗传差异,这有助于进一步研究分析原噬菌体的种类差异对柑橘木虱传毒机制的影响。
柑橘木虱;黄龙病菌;原噬菌体
柑橘黄龙病(HLB)是柑橘生产上的毁灭性病害,可危害各种柑橘类植物,在中国尤其对江西赣州脐橙造成了严重危害(陈丽芬等, 2017)。因此近些来年研究者对赣南地区黄龙病的发生、传播与防治进行了大量的研究,如刘登全等(2012)利用PCR扩增核苷酸序列测定技术在江西省大余县柑橘果园检测到了柑橘黄龙病菌。为了找到防治黄龙病的方法,其后利用椪柑、晚熟温州蜜柑、琯溪蜜柚三种柑橘品种对黄龙病的抗性情况进行了测试,结果表明晚熟温州蜜柑耐病性最强,而椪柑对柑橘黄龙病表现为高度感染(刘登全 等, 2014)。此外,陈慈相等(2015)对赣南地区柑橘黄龙病发生规律进行了研究。随后卢占军等(2016)基于SSR标记探讨了赣南柑橘黄龙病菌的遗传多样性,且同年金丹凤等(2016)、熊大维等(2016)分析了赣南脐橙柑橘黄龙病株内共生细菌的群落结构具有多样性。
原噬菌体为整合于宿主菌基因组中的噬菌体(Boydetal., 2002; 黎庶等, 2009),其区域已成为解析黄龙病病原遗传多样性及病害流行规律的重要靶标,且其多个高度变异区域已被应用于黄龙病菌种内多样性分析。如注释为噬菌体阻遏蛋白的基因位点包含多个重复单元序列,不同地理来源的黄龙病菌株系间的重复序列重复数差异显著(Chenetal., 2010; 谭锦, 2013);利用原噬菌体DNA聚合酶基因研究东南亚各国“Ca.L. asiaticus”的变异水平,将所有株系分为5簇(Tomimuraetal., 2009);利用原噬菌体末端酶基因位点研究发现黄龙病菌中国株系具有地区变异性(Liuetal., 2011)。此外,对黄龙病菌UF506株系中两种环状原噬菌体SC1和SC2基因组序列的测定,有助于在原噬菌体基因组上寻找更多的变异位点,这对病原菌遗传多样性的研究起到了重要的推动作用(Zhangetal., 2011; Zhouetal., 2011)。
2013年,Zhou等利用美国佛罗里达黄龙病菌psy62株系基因组中FP1和FP2噬菌体同源序列的基因区域对佛罗里达株系种群分化进行了进一步的研究,在此区域中共检测到9种噬菌体/原噬菌体类型,并且检测到在寄主植物和介体昆虫体内黄龙病菌含量情况受噬菌体/原噬菌体调节。且在之后有研究发现中国株系的原噬菌体与佛罗里达株系存在显著差异(Wangetal., 2014) 。但目前国内未有对各黄龙病发生区感病植物和柑橘木虱体内噬菌体/原噬菌体类型差异的详细研究(Morganetal., 2012),而这对于深入研究原噬菌体在介体传菌中的作用是至关重要的。本研究将利用黄龙病菌psy62株系基因组中FP1和FP2噬菌体同源序列区域内的基因位点,对中国赣南地区各柑橘产区柑橘木虱体内“Ca.L. asiaticus”株系中原噬菌体种群分化进行研究,进而探讨不同地理来源的黄龙病菌株系原噬菌体的遗传差异性;及分析柑橘木虱“Ca.L. asiaticus”株系原噬菌体类型与中国柑橘叶片株系是否存在遗传差异。对“Ca.L. asiaticus”株系中原噬菌体遗传差异性的研究,有助于加速找到原噬菌体在介体昆虫-柑橘木虱传菌过程中所起的作用,这将对揭秘柑橘木虱专性传播黄龙病有重大意义。
1 材料与方法
1.1 供试虫源
根据柑橘种植及黄龙病发生情况,选择赣南赣县、寻乌、于都、定南、瑞金、南康、崇义等多个柑橘产地为采集地点(采集信息见表1)。并于感染黄龙病的柑橘植株叶片上采集柑橘木虱,放入盛有无水乙醇的离心管中,带回实验室、鉴定并保存于-20℃冰箱备用。
1.2 试验方法
1.2.1总DNA提取
采用北京天根生化技术有限公司的基因组用DNA提取试剂盒(TIANamp Genomic DNA Kit)分别提取67头柑橘木虱样品的总DNA,利用引物OI1/OI2c对样品进行PCR检测以确定携带黄龙病菌。
1.2.2黄龙病菌阳性检测
利用引物OI1/OI2c对已采集到的67头柑橘木虱样品进行黄龙病菌阳性检测(Jagoueixetal., 1994)。PCR采用30 μL体系,包括15 μL Mix, 10.5 μL ddH2O, 5 μmol浓度引物各1.5 μL,1.5 μL DNA模版。反应程序见表2。
PCR扩增产物与6× Loading buffer 混匀后在1.2%琼脂糖凝胶中100 V电泳40 min,凝胶经EB染色后,使用UVI凝胶成像系统观察结果并拍照。
1.2.2柑橘木虱阳性样品原噬菌体目标位点扩增及检测
本次实验主要利用原噬菌体通用引物LJ753/LJ759和两个设计的特异性引物对柑橘木虱阳性样品进行PCR扩增。PCR采用30 μL体系,包括15 μL Mix, 10.5 μL ddH2O, 5 μmol浓度引物各1.5 μL,1.5 μL DNA模版。引物及反应程序见表3。

表1 柑橘木虱样本信息

表2 PCR引物及反应程序

表3 PCR引物及反应程序
PCR反应产物与-20℃保存备用。PCR产物的检测置于50×TAE的电泳缓冲液中,以1%琼脂糖凝胶电泳20 min,EB染色后,紫外凝胶成像仪观察并照相。
1.2.2PCR产物纯化、回收,克隆测序
采用生工生物工程(上海)有限公司的SanPrep柱式DNA胶回收试剂盒对PCR产物进行纯化、回收。回收的PCR产物直接与pGEM-T Easy载体(Promega)连接,转化大肠杆菌DH5α感受态细胞;选3个菌落PCR阳性样品送生工生物工程(上海)有限公司进行双向测序。
1.2.3序列分析
将测序返回的序列利用DNAStar软件包中的SeqMan功能进行拼接比对,并进行适当手工调整。利用Ediseq模块去除序列两端引物。用NCBI中的“BLAST”工具(http://blast.ncbi.nlm.nih.gov Blast.cgi)在GenBank中搜索关于佛罗里达黄龙病菌株系和中国地区柑橘叶片黄龙病菌株系psy62基因组中原噬菌体的同源序列。利用分子进化遗传分析软件MEGA进行系统进化分析,采用邻近法做Bootstrap验证(重复1000次)构建系统发育进化树。
2 结果与分析
2.1 柑橘木虱样品黄龙病菌检测
利用引物OI1/OI2c 对从赣南赣县、寻乌、于都、定南、瑞金、南康、崇义等7个柑橘产地所采集到的67头柑橘木虱样本进行黄龙病菌PCR扩增,共检测到42头柑橘木虱体内携带黄龙病菌(图1,表4)。

图1 引物OI1/OI2c对黄龙病的PCR扩增结果Fig.1 PCR amplification with primer sets OI1/OI2c against HLB注:M, DNA Marker;+CK, 阳性对照;-CK, 阴性对照;其他泳道, 不同来源地的样品。Note:M, DNA ladder marker;+CK, positive control; -CK, negative control;other lanes, samples from Diaphorina citri.

采集点Locality阳性样品Positivesamples 类型TypeTypeATypeA2TypeBTypeCTypeC1NoType阴性样品Negativesamples赣州市师范大学怡丰果园(GZ)GZ-YF1,4GZ-YF9GZ-YF2,3,6,7,8GZ-YF5,10,11寻乌县文峰乡(XW)XW-WF5,8,11XW-WF1,2XW-WF3,12XW-WF6XW-WF7,10XW-WF4,9定南县龙塘镇柏木村(DN)DN-BM2DN-BM7,10DN-BM5,9DN-BM1,3,4,6,8于都县银坑镇洋泾村(YD)YD-YK5,9YD-YK12YD-YK2,13YD-YK7YD-YK1,3,4,6,8,10,11瑞金县泽覃乡(RJ)RJ-ZT3RJ-ZT8RJ-ZT2,5,6RJ-ZT1,4,7南康县赤土畲族乡青塘村(NK)NK-QT1,4NK-QT3NK-QT6NK-QT2,5崇义县麟潭乡华山村CYCY-HS5,7CY-HS2CY-HS1CY-HS3,4,6
2.2 柑橘木虱阳性样品原噬菌体目的基因PCR扩增
利用通用引物LJ753/LJ759对42头柑橘木虱阳性样品(表4)进行原噬菌体目的基因PCR扩增,实验结果显示,引物LJ753/LJ759最优PCR反应体系为:15 μL PCR Mix,1.5 μL DNA模板,5 μmol/L 引物各2 μL,9.5 μL ddH2O;且最佳退火温度为54℃(表3)。电泳后能检测到目的条带(见图2)但未能得到相对应的完整序列。因此利用Primer Premier 5.0并以佛罗里达黄龙病菌psy62株系已有的序列为序列样本重新设计引物,新引物为LJ753/LJ49、LJ27/LJ759,运用这两对引物对柑橘木虱阳性样品重新PCR扩增,反复试验结果得出两对引物原噬菌体目的基因的最佳退火温度分别是45℃,54℃,电泳后不仅能检测到目的条带且测序得到了相对应的完整序列。

图2 柑橘木虱原噬菌体PCR扩增的效果电泳图Fig.2 PCR amplification for prophage of Diaphorina citri
2.3 柑橘木虱黄龙病菌psy62株系中原噬菌体的扩增类型
利用两对设计的新引物对已检测到的柑橘木虱阳性样品重新PCR扩增检测及测序,结果显示共28头柑橘木虱阳性样品(66.7%)可进行扩增测序,测序结果见表4,表5。测序结果表明,在江西赣南不同柑橘产地的柑橘木虱样品在黄龙病菌psy62株系FP1和FP2噬菌体同源序列基因组中共发现5种噬菌体/原噬菌体类型,分别为A, A2,B, C, C1;且A的检测率最高,为35.7%(10/28),其中赣县2头、寻乌3头、定南1头、于都2头、南康2头。B检测率次之,为25%(7/28),其中赣县1头、寻乌2头、于都2头、瑞金1头、崇义1头。C1检测率最低,为7.1%(2/28),且只在寻乌和瑞金各一头柑橘木虱阳性样品中检测到。A2样品中的检测率为17.9%(5/28),其中寻乌2头、于都1头、崇义2头。且在定南2头,南康1头,崇义1头等柑橘木虱样品中检测到C,检出率为14.3%(4/28)。

表5 柑橘木虱原噬菌体基于高度变异的FP1和FP2噬菌体基因内的不同类型及其检出率
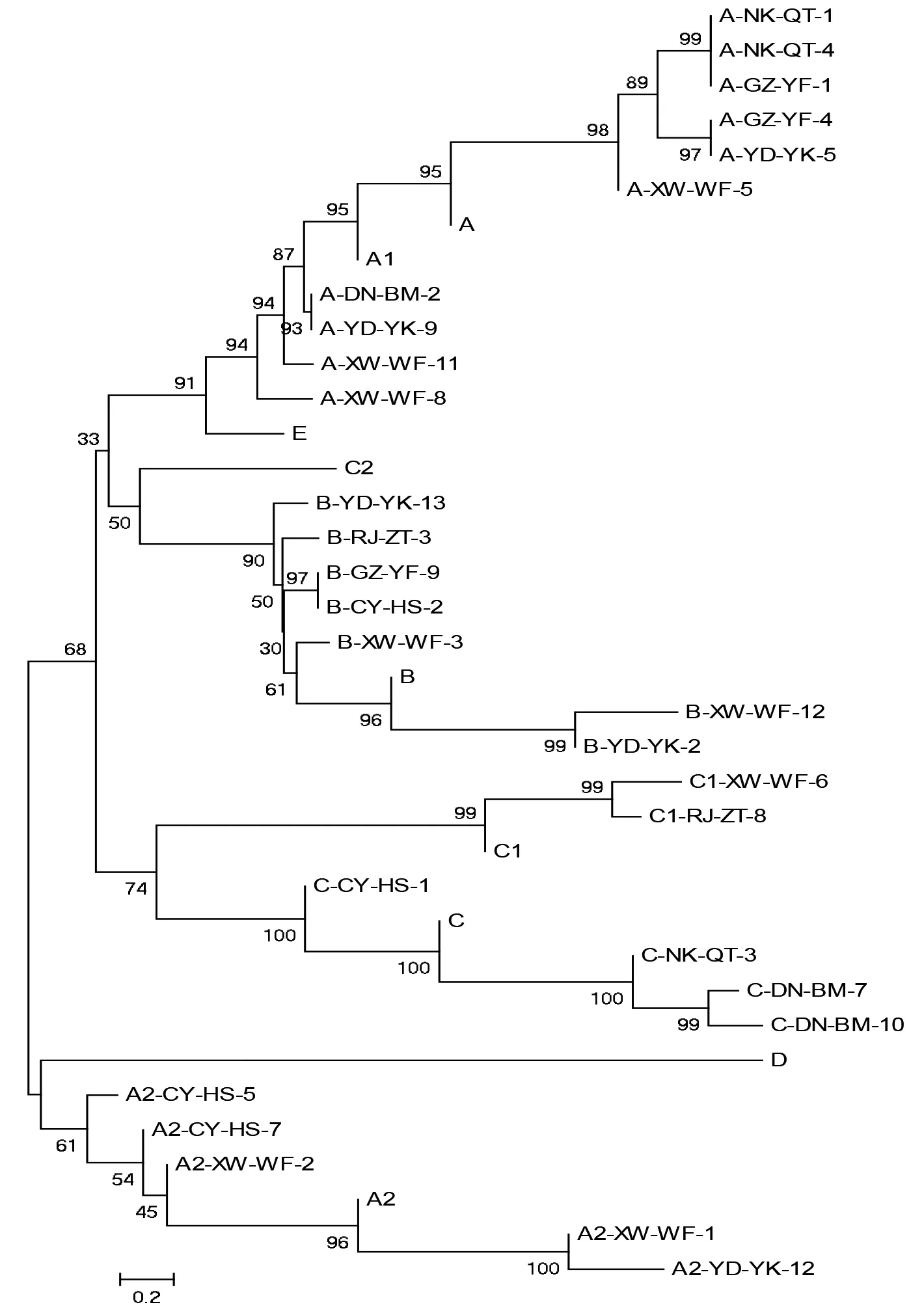
图3 柑橘木虱与弗罗里达佛罗里达州株系原噬菌体类型基因序列NJ系统发育树Fig.3 Neighbor-joining tree for prophage gene sequences of Diaphorina citri and Florida strains

图4 柑橘木虱与中国柑橘原噬菌体类型基因序列NJ系统发育树Fig.4 Neighbor-joining tree for prophage gene sequences of Diaphorina citri and China citrus
2.4 系统进化分析
运用MEGA 5.0软件,基于Kimura 2-parameter 模型,采用邻接法,对本研究得到的28条赣南柑橘木虱体内黄龙病菌株系中原噬菌体基因序列和在Genbank中下载的佛罗里达州黄龙病菌株系中原噬菌体参考序列(见表6)构建相对应的系统发育进化树(见图3)。
此次实验在柑橘木虱体内黄龙病菌中未检测到原噬菌体A1,C2,D,E,这与佛罗里达州株系存在显著差异。从系统发育树中可以看出赣南地区柑橘木虱体内不同原噬菌体类型之间存在一定的差异性,且同一原噬菌体类型在不同地区株系间存在较小的差异。但在赣县、于都、瑞金、崇义、寻乌等不同地区株系原噬菌体B的同源性存在较大的差异,原噬菌体A2在崇义和寻乌地区株系间差异性较大。并且进化树显示赣南地区柑橘木虱体内原噬菌体A,A2,B,C,C1分别与佛罗里达州株系原噬菌体A,A2,B,C,C1同源性差异不大,其中原噬菌体A和A1、E的同源性也存在较小差异。
在NCBI中Blast搜索中国地区柑橘叶片“Ca.L. asiaticus”原噬菌体相关序列,如云南、贵州、四川、广东、广西、福建、江西、浙江等地,并下载同源序列,通过邻接法(NJ)对所有赣南柑橘木虱“Ca.L. asiaticus”原噬菌体序列和下载的同源序列构建系统发育树(图4)。结果显示下载的中国等地柑橘叶片“Ca.L. asiaticus”株系原噬菌体类型均聚在一大枝上,表明不同中国地区的柑橘叶片原噬菌体的同源性差异不大,但在这一大枝内有些小枝的置信度较低,说明中国不同地理来源的柑橘叶片“Ca.L. asiaticus”株系间的原噬菌体还是存在一定的同源性差异。

表6 佛罗里达原噬菌体同源序列信息
续上表

序号No.类型Typename序列号AccessionNo.长度(bps)Length地点、寄主、菌株Place,host,strain参考文献References6A1JX2754942747佛罗里达州-小长春花,柑橘,柑橘木虱Zhou等,20137A2JX2754952728佛罗里达州-小长春花,柑橘木虱Zhou等,20138C1JX2754962682佛罗里达州-小长春花,柑橘,柑橘木虱Zhou等,20139C2JX2754972703佛罗里达州-小长春花,柑橘,柑橘木虱Zhou等,2013
从进化树中还可以看出赣南地区的柑橘木虱阳性样品中不同原噬菌体类型与中国不同地区柑橘叶片“Ca.L. asiaticus”株系的同源性均存在差异,且差异性显著。
3 结论与讨论
原噬菌体遗传变异程度的研究可为Ca.L. asiaticus 遗传多样性提供更多信息(Zhengetal, 2016)。本试验先利用通用引物LJ753/LJ759对42头阳性样本进行PCR扩增,检测到3000 bp左右的目的条带。但由于其片段过长且存在二级结构,导致无法得到完整的测序序列,分析也可能是原噬菌体超变异基因区域的高度变异性所致。于是利用设计的两对特异性引物对42头阳性样品重新PCR扩增,其中有14头柑橘木虱阳性样品未检测到目的条带,尝试调整设计引物的PCR扩增程序却仍未获得目的条带,表明这部分柑橘木虱阳性样品psy62株系中可能不存在FP1和FP2噬菌体相关的基因序列,亦或者是原噬菌体序列的缺失导致未能检测到目的条带。
且PCR扩增测序结果显示,在赣南地区柑橘木虱阳性样品体内黄龙病菌psy62株系基因组中FP1和FP2噬菌体同源序列区域内共发现5种噬菌体/原噬菌体类型,且不同类型的原噬菌体检出率略有差异:A检测率最高,为35.7%,C1检测率最低,为7.1%。而在佛罗里达州株系共被检测出9种,且这9种原噬菌体在柑橘叶片株系、柑橘木虱株系中的分布情况与在赣南地区柑橘木虱体内均有所不同(见表6)。进一步的系统发育树显示在赣南地区柑橘木虱阳性样品中不同原噬菌体类型之间存在一定的差异性,不同地区株系间的同一原噬菌体类型存在较小的差异。同时发现美国佛罗里达的“Ca.L. asiaticus”株系,中国各地区柑橘叶片“Ca.L. asiaticus”株系与中国赣南地区柑橘木虱“Ca.L. asiaticus”株系也均具有显著差异。
从以上实验结论中可以看出不同地理来源的柑橘木虱体内“Ca.L. asiaticus”原噬菌体种群有较大的遗传差异。但由于本实验中缺乏中国其他地点的柑橘木虱样品,缺乏一定的说服力,因此很有必要继续补充中国其他地点的数据及样品,在丰富数据后再进一步比较中国柑橘木虱体内“Ca.L. asiaticus”株系中原噬菌体的多样性与地理环境条件的关系,进而更加详细的探明柑橘木虱体内“Ca.L. asiaticus”噬菌体/原噬菌体的遗传差异性。当然对赣南地区黄龙病菌中原噬菌体遗传差异性的研究更加证实了 “美国黄龙病由中国传入” 的传言没有科学根据。
此研究结果也表明了原噬菌体在介体昆虫体内与柑橘叶片株系确实存在差异。如在中国赣南地区和佛罗里达柑橘木虱阳性样品中均未检测到原噬菌体类型D,且发现原噬菌体D与噬菌体iFP3有关,而噬菌体iFP3可能就是由FP1和FP2噬菌体重组产生的(Zhouetal., 2013)。由此推测包含类型D的噬菌体iFP3是否是阻止或减少病原菌传播的关键:如在传播过程中类型D在介体昆虫体内杀死病菌终止传播,或是影响昆虫传播病菌重要的表达基因、亦或降低黄龙病菌浓度等方式。因此接下来的任务是要研究黄龙病菌中D原噬菌体为何在传播介体柑橘木虱体内不存在?且D原噬菌体在柑橘木虱传播病菌过程中到底担任了什么角色?这将有助于更进一步的了解柑橘木虱传播病菌的真正机制。
)
Boyd EF, Brussow H. Common themes among bacteriophage-encoded virulence factors and diversity among the bacteriophages involved[J].TrendsMicrobiol., 2002, 10: 521-529.
Chen CX, Zhang J, Xie JZ,etal. Reseach the occurrence regularity of huanglongbing in Gannan[J].SouthChinaFruits, 2015, 44(6): 43-45. [陈慈相, 张倩, 谢金招, 等. 赣南地区柑桔黄龙病发生规律研究[J]. 中国南方果树, 2015, 44(6): 43-45]
Chen J, Deng X, Sun X,etal. Guangdong and Florida populations of “Candidatusliberibacter asiaticus” distinguished by a genomic locus with short tandem repeats[J].Phytopathology, 2010, 100(6): 567-572.
Chen LF, Xu ZH, Xiao LF,etal. A study on the diversity of prophage types from “Candidatusliberibacter” inDiaphorinacitricollected from southern Jiangxi Province[J]. ActaAgriculturaeUniversitatisJiangxiensis, 2017, 39(2): 278-285. [陈丽芬, 徐昭焕, 肖丽芳, 等. 赣南柑橘木虱体内黄龙病菌原噬菌体多样性的研究[J]. 江西农业大学学报, 2017, 39(2): 278-285]
Jin DF, Xiong DW, Huang GC,etal. Comparison of cultured and uncultured endophytic bacterial community in huanglongbing affected Gannan navel oranges[J].JiangxiScience, 2016, 34(4): 434-437. [金丹凤, 熊大维, 黄国昌, 等. 赣南脐橙柑橘黄龙病株内生细菌群落多样性研究[J]. 江西科学, 2016, 34(4): 434-437]
Li S, Hu FH. Prophages[J].InstituteofMicrobiology, 2009, 36(3): 432-438. [黎庶, 胡福泉. 前噬菌体[J]. 微生物学通报, 2009, 36(3): 432-438]
Liu DQ, Tu HM, Li L,etal. PCR detection of citrus huanglongbing pathogen in Dayu County of Jiangxi Province[J].BiologicalDisasterScience, 2012, 35(3): 261-263. [刘登全, 涂怀妹, 李霖, 等. 江西省大余县柑橘黄龙病菌PCR检测[J]. 生物灾害科学, 2012, 35(3): 261-263]
Liu DQ, Cui CY, Jiang JX,etal. Resistance identification of different citrus cultivars against huanglongbing[J].ActaAgriculturaeUniversitatisJiangxiensis, 2014, 36(1): 97-101. [刘登全, 崔朝宇, 蒋军喜, 等. 不同柑橘品种对黄龙病的抗性鉴定[J]. 江西农业大学学报, 2014, 36(1): 97-101]
Liu R, Zhang P, Pu X,etal. Analysis of a prophage gene frequency revealed population variation of “Candidatusliberibacter asiaticus” from two citrus-growing provinces in China[J].PlantDisease, 2011, 95(4): 431-435.
Lu ZJ, Liu YX, Ding P,etal. Genetic diversity of “Candidatusliberibacter asiaticus” in Gannan using SSR markers[J].SouthChinaFruits, 2016, 45(5): 1-6. [卢占军, 刘映雪, 丁鹏, 等. 基于SSR标记的赣南柑桔黄龙病菌遗传多样性分析[J]. 中国南方果树, 2016, 45(5): 1-6]
Morgan JK, Zhou L, Li W,etal. Improved real-time PCR detection of “Candidatusliberibacter asiaticus” from citrus and psyllid hosts by targeting the intragenic tandem-repeats of its prophage genes[J].MolecularandCellularProbes, 2012, 26(2): 90-98.
Tan J. Genetic Diversity of Prophage Region from “CandidatusLiberibacter asiaticus” in China[D]. Chongqing: Master’s thesis of Southwest University, 2013, 1-90. [谭锦. 中国柑橘黄龙病病原菌原噬菌体遗传多样性研究[D]. 重庆:西南大学硕士论文, 2013, 1-90]
Tomimura KS, Miyata N, Furuya K,etal. Evaluation of genetic diversity among “Candidatusliberibacter asiaticus” isolates collected in Southeast Asia[J].Phytopathology, 2009, 99(9): 1062-1069.
Wang XC, Zhou X, Deng H,etal. Molecular characterization of a mosaic locus in the genome of “Candidatus liberibacter asiaticus” [J].BMCMicrobiology, 2012, 12(1): 1-7.
Wang XH, Su L, Huang X,etal. Identification of a novel 1033-nucleotide deletion polymorphism in the prophage region of “liberibacter asiaticus”: Potential applications for bacterial epidemiology[J].JournalofPhytopathology, 2014, 163(7-8): 681-685.
Xiong DW, Jin DF, Huang GC,etal. Endophytic bacterial community in Gannan navel orange affected by huanglong disease[J].MicrobiologyChina, 2016: 1-12. [熊大维, 金丹凤, 黄国昌, 等. 柑橘黄龙病赣南脐橙内生菌种群结构分析[J]. 微生物学通报, 2016: 1-12]
Zhang SZ, Flores CL, Zhou BH,etal. “Candidatusliberibacter asiaticus” carries an excision plasmid prophage and a chromosomally integrated prophage that becomes lytic in plant infections[J].MolecularPlant-MicrobeInteractions, 2011, 24(4): 458-468.
Zheng Z, Bao M, Wu F,etal. Predominance of single prophage carrying a CRISPR/cas system in “Candidatusliberibacter asiaticus” strains in Southern China[J].PLoSONE, 2016, 11(1): e0146422.
Zhou LCA, Powell MT, Hoffman W,etal. Diversity and plasticity of the intracellular plant pathogen and insect symbiont “liberibacter asiaticus” as revealed by hypervariable prophage genes with intragenic tandem repeats[J].AppliedandEnvironmentalMicrobiology, 2011, 77(18): 6663-6673.
Zhou L, Powell CA, Wenbin L,etal. Prophage-mediated dynamics of “Candidatusliberibacter asiaticus” populations, the destructive bacterial pathogens of citrus huanglongbing[J].PLoSONE, 2013, 8(12): e82248.
AstudyonthediversityofprophagetypesfromCandidatusliberibacterstrainpsy62inDiaphorinacitricollectedfromSouthernJiangxiProvince
CHEN Li-Fen1,2, XU Zhao-Huan1*, XIAO Li-Fang1, ZHANG Li-Qing1,WANG Jian-Guo1**
(1.School of Agriculture Science, Jiangxi Agricultural University, Nanchang 330045, China;2.Agriculture and Grain Bureau of Chongyi, Chongyi 341300, Jiangxi Province,China)
In order to research on the genetic diversity of prophage region fromCandidatusliberibacter strain psy62 inDiaphorinacitri, and the genetic differenty betweenD.citriand citrus strains. For this experiment, using there PCR primers to detect positive samples ofD.citriwere collected from seven locality in Gannan. Initial result indicated that the percentage of type A and type B was relatively high, the percentage of type C was lowest. The study result suggested that different location of prophage inD.citripopulations have larger genetic differences. And “Ca. L. asiaticus” strains of Florida in the United States, Chinese “Ca. L. asiaticus” strains of citrus are all have significant genetic differences to “Ca.L. asiaticus” strains ofD.citriin Gannan. It will further the research and analysis of the influences of the difference prophage species to the toxic mechanism ofD.citri.
Diaphorinacitri;Candidatusliberibacter asiaticus; prophage
陈丽芬,徐昭焕,肖丽芳,等.赣南柑橘木虱体内黄龙病菌psy62株系原噬菌体类型多样性的研究[J].环境昆虫学报,2017,39(6):1198-1206.
Q968.1;S433.39
A
1674-0858(2017)06-1198-09
国家自然科学基金(31360457) ; 江西省教育厅科技计划项目(GJJ150408)
陈丽芬, 女, 1991年生,研究方向为昆虫学, E-mail: chenlfjx@126.com
*共同第一作者, 徐昭焕, 女, 1984年生, 博士, 讲师, 研究方向为昆虫学, E-mail: hzzhaohuan@163.com
**通讯作者Author for correspondence, E-mail: jgwangjxau@qq.com
Received: 2017-05-15; 接受日期Accepted: 2017-10-22
