Constitutive expression of feedback-insensitive cystathionine γ-synthase increases methionine levels in soybean leaves and seeds
2018-01-04YUYangHOUWenshengYaelHachamSUNShiWUCunxiangIfatMatityahuSONGShikuiRachelAmirHANTianfu
YU Yang, HOU Wen-sheng, Yael Hacham, SUN Shi, WU Cun-xiang, Ifat Matityahu SONG Shikui, Rachel Amir, HAN Tian-fu
1 Key Laboratory of Soybean Biology (Beijing), Ministry of Agriculture/Institute of Crop Sciences, Chinese Academy of Agricultural Sciences, Beijing 100081, P.R.China
2 Laboratory of Plant Science, Migal Galilee Technology Center, Kiryat Shmona 12100, Israel
3 Tel-Hai College, Upper Galilee 11016, Israel
RESEARCH ARTICLE
Constitutive expression of feedback-insensitive cystathionine γ-synthase increases methionine levels in soybean leaves and seeds
YU Yang1*, HOU Wen-sheng1*, Yael Hacham2,3*, SUN Shi1, WU Cun-xiang1, Ifat Matityahu2, SONG Shikui1, Rachel Amir2,3, HAN Tian-fu1
1 Key Laboratory of Soybean Biology (Beijing), Ministry of Agriculture/Institute of Crop Sciences, Chinese Academy of Agricultural Sciences, Beijing 100081, P.R.China
2 Laboratory of Plant Science, Migal Galilee Technology Center, Kiryat Shmona 12100, Israel
3 Tel-Hai College, Upper Galilee 11016, Israel
Soybean (Glycine max (L.) Merr.) is a major crop that provides plant-origin protein and oil for humans and livestock. Although the soybean vegetative tissues and seeds provide a major source of high-quality protein, they suffer from low concentration of an essential sulfur-containing amino acid, methionine, which signi ficantly limits their nutritional quality. The level of methionine is mainly controlled by the first unique enzyme of methionine synthesis, cystathione γ-synthase (CGS).Aiming to elevate methionine level in vegetative tissues and seeds, we constitutively over-expressed a feedback-insensitive Arabidopsis CGS (AtD-CGS) in soybean cultivars, Zigongdongdou (ZD) and Jilinxiaoli 1 (JX). The levels of soluble methionine increased remarkably in leaves of transgenic soybeans compared to wild-type plants (6.6- and 7.3-fold in two transgenic ZD lines, and 3.7-fold in one transgenic JX line). Furthermore, the total methionine contents were signi ficantly increased in seeds of the transgenic ZD lines (1.5- to 4.8-fold increase) and the transgenic JX lines (1.3- to 2.3-fold increase) than in the wild type. The protein contents of the transgenic soybean seeds were signi ficantly elevated compared to the wild type,suggesting that the scarcity of methionine in soybeans may limit protein accumulation in soybean seeds. The increased protein content did not alter the pro file of major storage proteins in the seeds. Generally, this study provides a promising strategy to increase the levels of methionine and protein in soybean through the breeding programs.
soybean (Glycine max (L.) Merr.), constitutive expression, cystathionine γ-synthase (CGS), methionine content,storage protein
1. Introduction
Soybean (Glycine max (L.) Merr.) is a major crop throughout the world to feed humans for thousands of years. Their seeds possess high contents of protein (35–50%) and oil(approximately 20%) with additional high levels of vitamins and minerals as well as phytochemicals including iso fla-vones, tocopherols, saponins, and anthocyanins (Kim et al.2012). Additionally, the vegetative tissues of soybean can also be used to feed animals as a rich nutritional source.However, soybean seeds suffer from low levels of the sulfur containing amino acids, methionine, and cysteine (Cooper 1983; Amir et al. 2012). Thus, improving the sulfur amino acid contents in soybean, similar to lysine improvement in cereals (Yu et al. 2004; Mattoo et al. 2010), is an important goal of soybean breeding. In animals, cysteine can be converted from methionine, thus, the quality improvement of soybean can focus on elevating methionine content.
In vegetative tissues of plant, methionine is mainly regulated by the expression level of its first unique enzyme, cystathionine γ-synthase (CGS), which combines the carbon/amino skeleton derived from aspartate, with the sulfur moiety derived from cysteine (Kim et al. 1996). Recent studies performed in Arabidopsis, tobacco, Azuki bean, and soybean (Hanafy et al. 2013; Matityahu et al. 2013; Song et al. 2013; Cohen et al. 2014) have demonstrated that seed-speci fic expression of feedback-insensitive mutated forms of Arabidopsis CGS(AtD-CGS) (Hacham et al. 2006) led to signi ficantly higher levels of methionine in the transgenic seeds. However,previous biochemical and genetic studies mainly preformed in Arabidopsis and wheat proposed that methionine could also transport from vegetative tissues towards seeds in the form of S-methyl methionine (SMM) and then convert back to methionine in seeds (Bourgis et al. 1999; Ranocha et al.2001; Gallardo et al. 2007; Lee et al. 2008; Frank et al. 2014).
Although the biosynthetic pathway of methionine is clear(Fig. 1), the exact contribution of the aspartate family CGS and the SMM to methionine synthesis in seeds is still required for further study. Integrated comparative proteomic and transcriptomic analyses of different sections of Medicago truncatula seeds suggest that these two pathways contribute to methionine synthesis in seeds at different stages of seed development (Gallardo et al. 2007). The researchers implied that methionine is synthesized through the aspartate family pathway in endosperm and embryo up to the mid-stages of seed filling, but in the late stages of seed filling, methionine is mainly synthesized from the SMM, which is mainly from the phloem and degrades to methionine, since the seed coat supports the synthesis of the storage substances in filial tissues.And the transmitted compounds (mainly sugars) including SMM from the phloem supplies the methionine accumulation during the seed filling stage (Gallardo et al. 2007).
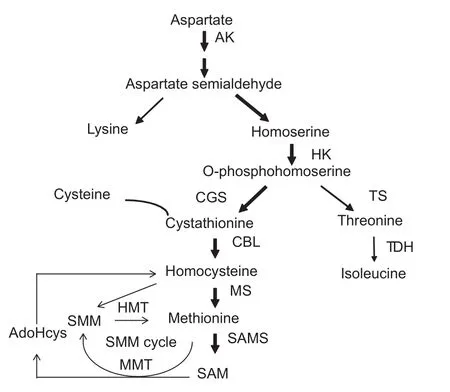
Fig. 1 The biosynthesis pathway of methionine. This depicts the biosynthesis branch of methionine in the aspartate family. AK,aspartate kinase; HK, homoserinekinase; CGS, cystathionine γ-synthase; CBL, cystathionine β-lyase; TS, threonine synthase; MS, methionine synthase; SAMS, SAM synthase;SAM, S-adenosylmethionine; AdoHcy, adenosylhomocysteine;MMT, methionine S-methyltransferase; HMT, homocysteine S-methyltransferase; TDH, threonine dehydratase. The schematic of the pathway was drawn according to previous report (Amir 2010).
Support to the assumption that higher levels of methionine in leaves can result in the accumulation of methionine in seeds, had also derived from the plants overexpressing heterologous feedback insensitive forms of the last two enzymes in the cysteine biosynthesis pathway, the yeast Met25 encoding to O-acetyl(thiol)lyase in flax plants (Czuj et al. 2009), and the bacterial gene encoding to serine acetyltransferase (EcSAT) in rice (Nguyen et al. 2012). These plants that have higher levels of cysteine and methionine contents in leaves exhibit signi ficantly higher levels of these amino acids in seeds of these plants, implying that these two amino acids that synthesized in vegetative tissues can be transferred and accumulated in the seeds. However, this is a plant-preference effect since transgenic tobacco plants, with signi ficantly higher levels of methionine and SMM in their leaves, have only slightly higher levels of these metabolites in seeds (Hacham et al. 2007, 2008).
The goals of this study were to over-express the AtD-CGS in soybeans and to reveal its effect on the content of methionine in vegetative tissues and seeds. This is an important biotechnological goal, since higher level of methionine in these tissues will improve their nutritional quality.In this study, we particularly used a late-maturing variety,Zigongdongdou (ZD) that adapts to grow at low latitudes, but can be planted at high latitude as a forage crop to produce green feed due to their in finite vegetative growth and large biomass (Jiang et al. 2011).
2. Materials and methods
2.1. Binary plasmid construction
The chloroplast transit peptide of pea rbcS-3A followed with the cDNA encoding AtD-CGS and the OCT terminator fragment, were removed by NcoI and XbaI double digestion.The fragment was then subcloned into the pET28a/35S plus vector, driven by the 35S promoter of cauli flower mosaic virus. The intact expression cassette was subsequently cloned into the plant transformation binary vector pTF-101.1,which carried glufosinate ammonium resistance (Becker et al. 1992) within the HindIII and XbaI restriction sites. The plasmid was transformed into Agrobacterium tumefaciens EHA101 by electroporation.
2.2. Growth and screening of transgenic plants
Transgenic soybeans from T0–T2generations were planted in the greenhouse at Chinese Academy of Agricultural Sciences (CAAS), Beijing. Transgenic soybeans after T3generation along with the wild types were grown in experimental plots with 1.5 m row and 0.6 m space in the Experimental Farm af filiated to Nanchong Academy of Agricultural Sciences, Sichuan Province, China. It is geographically located in 28.36°N, 115.55°E. A concentration of 160 mg L–1glufosinate used for T0plants, the leaves were painted with 160 mg L–1glufosinate for three times during V3 (the 3rd trifoliolate unfolded) to V4 stages on different leaves(Fehr and Caviness 1977). The plants after T1generation were sprayed with 250 mg L–1glufosinate when the second trifoliate fully developed for screening. The transgenic soybeans were naturally self-pollinated, podded and harvested for generations to obtain homozygous individual plants.
2.3. Transcription level detection
Total RNA was isolated using Trizol Reagent (Life Technologies, Thermo-Fisher, USA), according to the RNA extraction protocol, and it was treated with DNase I (QIAGEN, China).Nucleic acid quanti fication was carried out by using Nanodrop 2000 (Thermo, USA). A total of 1 mg of RNA were used as template for cDNA 1st strand synthesis, using a PrimeScript 1st Strand cDNA Synthesis Kit (TaKaRa, Japan).
AtD-CGS was ampli fied with forward (5´-AGCAATGGT GGAAGAGTAAA-3´) and reverse (5´-CCATACTCGAAACT CACACTC-3´) primers. GmLectin was ampli fied with the forward (5´-CTTTCTCGCACCAATTGACA-3´) and reverse(5´-TCAAACTCAACAGCGACGAC-3´) primers. The reaction procedure adopted as: step one, 94°C for 5 min; step two, 94°C for 30 s, 55°C for 30 s, 72°C for 40 s; step three,72°C for 5 min.
2.4. SDS-PAGE and Western blot analysis
Soybean leaves and seeds were homogenized with 25 mmol L–1phosphate extraction buffer (pH=8.0) and centrifuged at 4°C and 14 000 r min–1for 15 min. The supernatant was transferred to a new Eppendorf tube. The protein samples,25 μg each, were fractionated on a 12% SDS-PAGE (sodium dodecyl sulfate polyacrylamide gel electropheresis), transferred onto a PVDF membrane (Amersham Biosciences,USA), blocked for 2 h in 5% (v/v) skim milk, and incubated with α-AtD-CGS primary antibody (speci fic against AtDCGS) that previously prepared in our lab (dilution ratio 1:100), followed by incubation with a goat α-rabbit secondary antibody (dilution ratio 1:200). The membrane was incubated in ECL (Thermo Scienti fic Kit, Thermo-Fisher, USA),and immune detection was conducted with an enhanced chemiluminescence Kit (Pierce, USA), according to the manufacturer’s instructions.
2.5. Measurement of amino acids
Free amino acids were extracted from soybean leaves or seeds, and amino acid detection was performed by GC-MS(gas chromatography-mass spectrometry), as previously described (Cohen et al. 2014). A total of 25 mg of mature seeds or 100 mg leaves was ground in 1 mL of methanol.After 10 min of centrifugation at 4°C and 14 000 r min–1, the supernatant was collected and 700 μL of chloroform and 375 μL double distilled water were added to the supernatant.After 30 min of centrifugation at 3 000 r min–1, 450 μL was collected from the upper water phase, dried and dissolved in 140 μL of 20 mg mL–1methoxyamine hydrochloride in pyridine. We used 30 m Rtx-5SIL MS column (0.25 mm inner diameter, 0.25 μm film thickness) and performed the gas chromatography (GC) step with the parameter set as the injection temperature 230°C and the interface 250°C.The helium carrier gas flow was set as 1 mL min–1. And the temperature was set as: 70°C for 5 min, 5°C min–1raised to 310°C and last for 1 min. The MS scanning range was 50–600 m/z for 2 scans s–1. We calculated the quantity of amino acids by using the calibration curves of the standards and by using the norlucine as the reference substance for quanti fication (Amira et al. 2005).
To measure total amino acid content, 10 mg of mature seeds were ground to powder using a Retsch MM 301 homogenizer (Retsch, Germany), suspended in 10 mL Norleucine solution (2 mg mL–1) and vortexed vigorously. For leaves, 50 mg samples were ground with liquid nitrogen into a fine powder and suspended in 10 mL Norleucine solution with vigorously vortexed. A total of 200 mL of this suspension was collected in a glass tube, frozen at –70°C and dried in a lyophilizer, followed by 6 N HCl acidic hydrolysis for 22 h at 110°C under a vacuum. The hydrolysis products were suspended in 1 mL double distilled water containing 0.3 mL chloroform. Following centrifugation (4 000 r min–1for 30 min), 0.3 mL aliquots were collected from the upper phase, and the samples were treated as described for free amino acids analysis (Cohen et al. 2014).
2.6. Determination of total protein and lipid
Bruker MATRIX-I FT-NIR Spectrometer was used for the total protein and lipid determination of soybean seeds. The soybean determination model was calibrated, the accuracy of obtained data from this model was highly correlated(r>0.95) with the data obtained using Kjeldahl and Solvent methods for protein and lipid determination. Seeds from each individual were placed into three sample vials for the measurements and the values were calculated and used for subsequent analysis.
2.7. Statistical analysis
The data were analyzed by using analysis of variance(ANOVA) and the signi ficant difference was evaluated by Duncan’s multiple comparison at the signi ficance level of 0.05 in this study.
3. Results
3.1. Generation of transgenic soybeans overexpressing AtD-CGS
The feed-back insensitive AtD-CGS gene which possesses an internal deletion in the N-terminus region was used(Hacham et al. 2006). The expression cassette containing the AtD-CGS gene under the control of the CaMV 35S promoter was cloned into the plant binary expression vector,pTF-101.1, which carried BASTA resistance (Fig. 2-A). The A. tumefaciens strain EHA101 harbored the plasmid was used to transform Zigongdongdou (ZD) and Jilinxiaoli 1 (JX),two soybean varieties from South and Northeast China,respectively. The Agrobacterium-mediated cotyledonary node transformation method (Appendix A) was adopted to obtain transgenic soybeans. Leaves from transgenic ZD and JX lines (T3generation) that resistant to 250 mg L–1BASTA were screened at V3 stage (Fehr and Caviness 1977). Leaf samples were collected at R5 stage (beginning seed) (Fehr and Caviness 1977) for the expression analysis of AtD-CGS gene by semi-quantitative RT-PCR(Fig. 2-B). Two transgenic ZD (ZD36 and ZD42-1) lines and one JX (JX51) line that have the high expression levels were selected for the further analysis. Transgenic soybean lines were self-pollinated and screened by BASTA to obtain homozygous plants among generations.
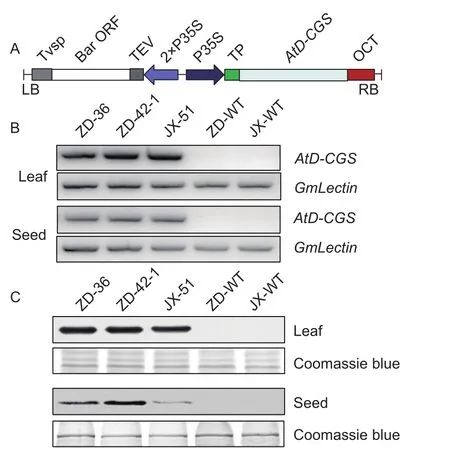
Fig. 2 The expression of AtD-CGS in transgenic soybeans.The transgenic lines were validated by expression analysis.A, the exhibited construct used for soybean transformation.Tvsp, terminator from soybean vegetative storage protein gene; Bar, phosphinothricin acetyl transferase gene; TEV,tobacco etch virus translational enhancer; P35S, the cauli flower mosaic virus 35S promoter; TP, chloroplast targeting peptide;AtD-CGS, Arabidopsis thaliana deleted cystathionine γ-synthase; OCT, octopine synthase terminator. B, the transcription level of AtD-CGS in the leaves and seeds by semiquantitative RT-PCR. ZD, Zigongdongdou; JX, Jilinxiaoli 1; WT,wild type. C, the Western blot results in the leaves and seeds,Coomassie blue staining was used to ensure the equal loading.
Semi-quantitative RT-PCR and Western blot were performed to detect the transcript (Fig. 2-B) and protein expression level (Fig. 2-C) in leaves at R5 stage and in seeds after full maturity (R8) (Fehr and Caviness 1977)from T3generation. These analyses demonstrate that the AtD-CGS gene expressed in leaves and seeds.
3.2. The soluble methionine level signi ficantly increased in the leaves of transgenic soybean
To assess whether the high expression level of AtD-CGS affected methionine content, the levels of soluble methionine in the leaves at R5 stage of these transgenic lines and the wild type were measured by using GC-MS. The level of soluble methionine content increased in ZD-36 and ZD-42-1 by 6.6- to 7.3-fold, respectively, while JX-51 showed higher methionine content by 3.7-fold compared to the corresponding wild type in leaves (Fig. 3-A). To determine whether a higher level of soluble methionine affected total methionine, including methionine incorporated into seed proteins, amino acid levels were measured after protein hydrolysis. Whereas the analysis revealed that the level of total methionine in leaves was not signi fi-cantly changed between the transgenic and the wild-type lines (Fig. 3-B).
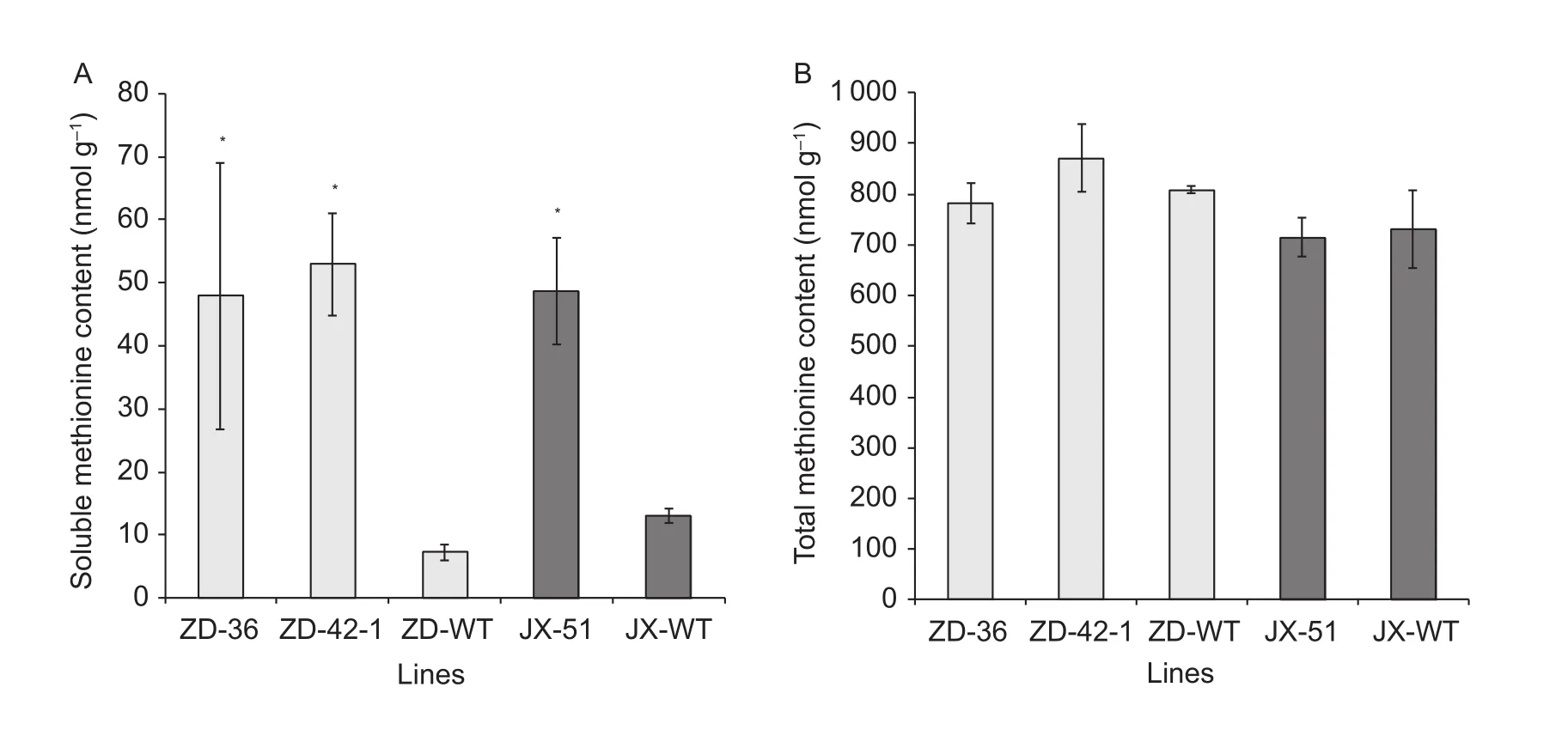
Fig. 3 The soluble (A) and total methionine (B) contents in leaves of transgenic and wild type (WT). Methionine content was detected using GC-MS (gas chromatography-mass spectrometry) using Norleucine as the internal control and calculated as nmol g–1 dry weight of the leaves. ZD, Zigongdongdou; JX, Jilinxiaoli 1. Mean±SD values were showed, the histogram showed each individual line transgenic or wild type line. Statistically signi ficant differences (P<0.05) are marked by a asterisk.
3.3. The methionine content was altered in the seeds of transgenic soybean expressing AtD-CGS
We examined the level of methionine in matured seeds of these transgenic plants. The T4lines ZD-36a, ZD-36b and ZD-42-1a, ZD-42-1b derived from the transgenic event ZD-36 and ZD-42-1, and the lines JX-102, JX-107, JX-112,JX-113, and JX-119 derived from transgenic event JX-51 were tested.
The soluble methionine content was not signi ficantly increased in seeds of the transgenic ZD lines compared to the wild type (Fig. 4-A). Notably, although the level of methionine did not signi ficantly increase, the level of aspartate,the precursor of the carbon-amino skeleton to aspartate family of amino acids (Fig. 1), that include methionine, lysine, threonine and isoleucine, were signi ficantly increased in these seeds. Additionally, the level of lysine signi ficantly reduced in three out of four transgenic lines compared to the wild type (Appendix B).
The total methionine level determined after protein hydrolysis signi ficantly increased in transgenic ZD lines compared to the wild type, demonstrating a 1.5- to 2.9-fold in ZD-36a, ZD-36b and 2.0- to 4.8-fold increase in ZD-42-1a,ZD-42-1b, respectively (Fig. 4-B; Appendix C). In addition,the levels of alanine and aspartate were signi ficantly elevated in three out of four lines.
In seeds of the transgenic JX-51-derived lines, the soluble methionine content was signi ficantly increased by 2.7- to 4.4-fold (Fig. 4-C). The contents of methionine related to amino acids, homoserine, asparagine, lysine, threonine, and isoleucine were also signi ficantly increased (Appendix D).
The level of total methionine in seeds increased by 1.3- to 2.3-fold in the transgenic lines compared with the wild-type JX51 (Fig. 4-D). Additionally, the levels of serine, threonine,isoleucine, and asparagine were signi ficantly accumulated at least in three lines (Appendix E).
3.4. Total protein and lipid contents in transgenic soybean seeds
The increased amount of total amino acids in the ZD and JX lines suggested that these seeds may have a higher protein content. To verify this, we measured the total protein content in matured seeds. We found that the amounts of proteins in ZD-36 and JX-51 lines had signi ficantly higher levels of proteins (with an increase of 2%) compared to their wild types, whereas no signi ficant change was observed for ZD-42-1 (Table 1).
As previous suggested, methionine and cysteine contents might affect the major storage proteins in soybean seeds(Holowach et al. 1984). Thus, to identify if higher levels of total methionine in seeds may cause alteration of major storage protein pro file in soybean seeds, the seeds of ZD-36(with both signi ficant higher total methionine and protein content in seeds) and ZD wild type were detected using difference gel electrophoresis (DIGE). The results suggested that the protein pro file of the major storage proteins was not signi ficantly altered (Appendix F).
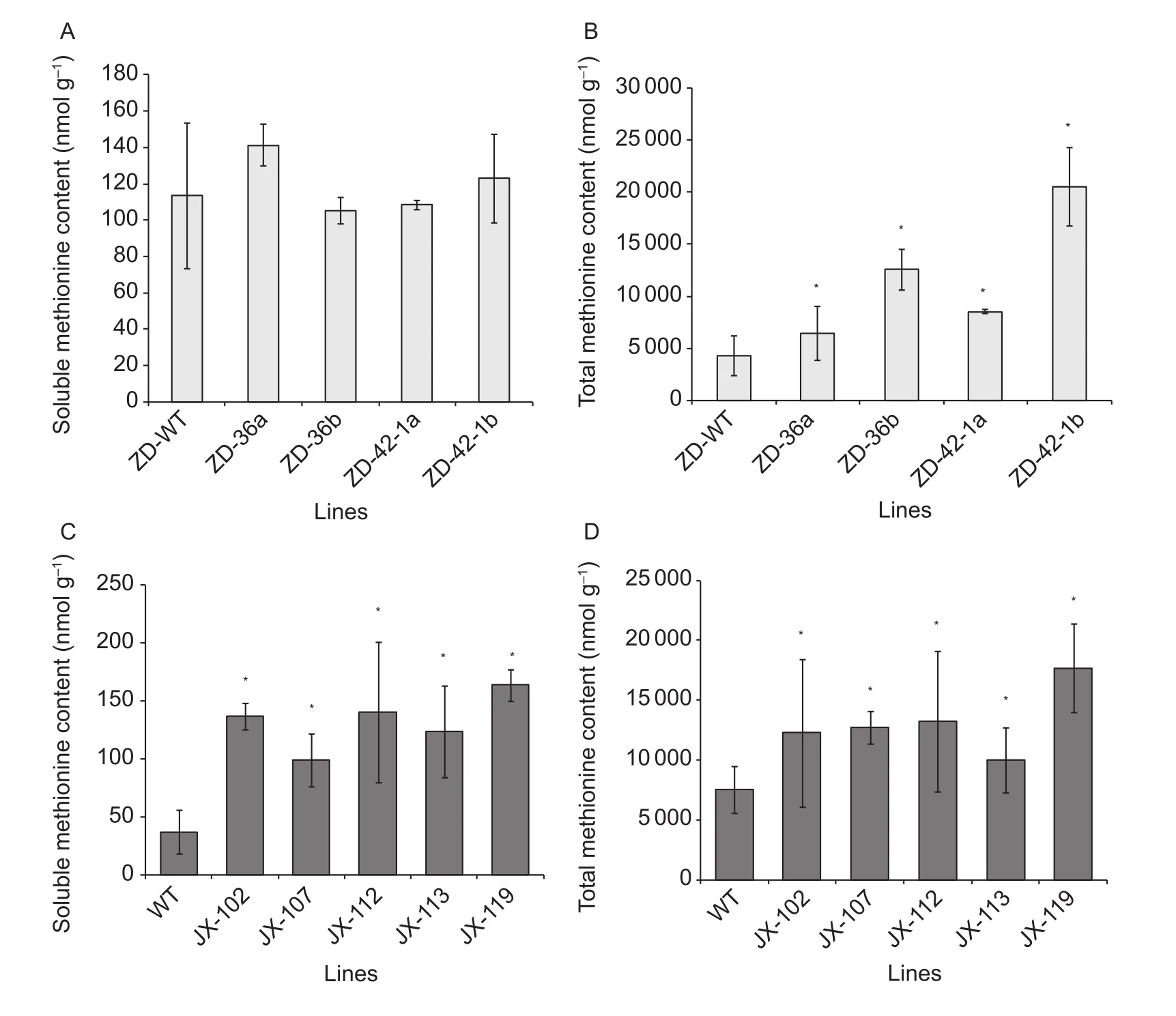
Fig. 4 The soluble (A and C) and total methionine (B and D) contents in seeds of transgenic and wild type. Methionine contents in mature dry seeds of wild types and transgenic seeds following GC-MS (gas chromatography-mass spectrometry) analysis.Norleucine used as the internal control and calculated as nmol g–1 dry weight of the seeds. ZD, Zigongdongdou; JX, Jilinxiaoli 1;WT, wild type. Mean±SD values were shown, the histogram showed each individual line transgenic or wild-type line. Statistically signi ficant differences (P<0.05) are marked by a asterisk.
It has been reported that high protein content in soybean seeds is associated with a decrease in oil concentration(Hernández-Sebastià et al. 2005). Thus, the changes in total protein levels in the transgenic soybean seeds may affect the levels of lipids. The result revealed that the total lipid content of the transgenic seeds was signi ficantly reduced compared with that of the wild type. In JX-51, the total protein content in seeds was signi ficantly increased,and the lipid content was signi ficantly decreased (Table 1).
3.5. No signi ficant morphological changes were observed in transgenic lines
To evaluate the effects of AtD-CGS overexpression on soybean development, the morphological traits of transgenic soybeans were compared to wild-type plants. There were no signi ficant morphological changes in the plants during growth (data not shown) or in the matured seeds. The weight of the 100-seed of transgenic soybeans was similar to that of the wild type. Similarly, the germination rate wasnot signi ficantly altered as well (Appendix G).
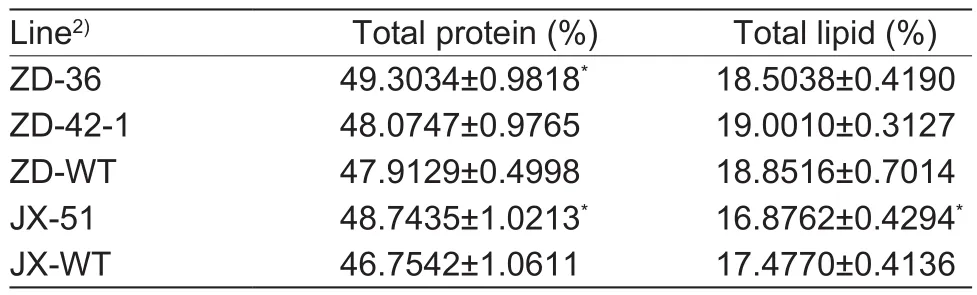
Table 1 Seed total protein and lipid contents of transgenic ZD and JX lines compared to wild type1)
4. Discussion
CGS gene was shown to be inhibited by feedback regulation at the post-transcriptional level (Chiba et al. 1999; Ominato et al. 2002; Hacham et al. 2006). The previous studies demonstrated that several deletion forms of CGS in the first exon was insensitive to the feedback regulation and remained the high levels expression of CGS. Therefore, it led to continuous synthesis of the methionine and promoted the accumulation of methionine. Our results demonstrated that by overexpressing the AtD-CGS gene, the soluble methionine level was signi ficantly elevated in the leaves of transgenic soybeans, while the total methionine content in leaves was not signi ficantly altered. These finding propose that the proteins that accumulated in seeds were not methionine-rich proteins. The excess of soluble methionine is probably transported to sink organs and accumulated there (e.g., seeds). However, the signi ficantly higher levels of methionine that incorporated to seeds storage proteins in both ZD and JX lines, suggested that seeds may have more methionine-rich proteins, or the levels of seed storage proteins were universally increased. However, protein profiling did not show signi ficant effects, which required more accurate analysis to de fine if changes occur. In addition, it demonstrated that over-expression of feedback insensitive CGS not only resulted in increased levels of methionine, but also other aspartate-derived amino acids. We hypotheses that the increased synthesis and accumulation of methionine led to excess supply of biosynthesis precursors into the aspartate derived amino acids synthesis pathways that promote the accumulation of related amino acids including lysine, threonine, and isoleucine.
We hypotheses that the increased synthesis and accumulation of methionine led to excess supply of biosynthesis precursors into the aspartate derived amino acids synthesis pathways that promote the accumulation of related amino acids including lysine, threonine and isoleucine.
The higher level accumulation of methionine in the transgenic seeds could result from methionine transported from the leaves, and also from de-novo synthesis of methionine in the seeds. The fold change of the total methionine (1.5- to 2.9-fold in ZD-36 and 2.0- to 4.8-fold in lines derived from ZD-42-1) is higher than those obtained in our previous study (Song et al. 2013), in which ectopic expression by seed-speci fic promoter legumin B4 failed to facilitate vegetative organs as the major methionine synthesis sources.The legumin B4 promoter expresses the heterologous gene only in late phase of seed development, when amino acids and protein are synthesized, whereas the 35S CaMV promoter is constitutively expressed through all stages of seed development (Sunilkumar et al. 2002) and causes excess synthesis of methionine from leaves. Therefore, the results implied that constitutive expression of At-DCGS led to higher methionine in soybean seeds and leaves. High level of methionine in leaves may also contribute to the accumulation of the soybean seeds protein. This is also in accordance to the findings described in Medicago truncatula by Gallardo et al. (2007). The required protein diet needs total 3.5 g sulfur-containing amino acids (methionine and cysteine) per 100 g, whereas in soybean seed, the content of methionine and cysteine is about 1.3 g per 100 g (Shewry 2000; WHO 2007), which needed an increment over 2.6 folds. In this study, we generated the transgenic soybean lines that can provide the highest 4.8-fold increases in methionine content which met the requirement for feeding human and animals.
The higher level of soluble methionine in JX transgenic seeds leads to signi ficantly higher levels of other soluble amino acids related to methionine. Such effect was not observed in ZD transgenic lines. Elevations in the other amino acids were previously found in seeds of tobacco,soybean and Arabidopsis, that seed-speci fic expressing the AtD-CGS gene, which affected the levels of total amino acids and proteins in these seeds. The increased total methionine in soybean caused the additional accumulation of storage proteins, while the protein pro file was not largely altered in transgenic seeds. Hence, the de ficiency in methionine may limit the accumulation of seed storage protein in soybeans and other legume plants.
In the view of the efforts toward quality improvement of feed crops in recent years (Kingston-Smith et al. 2013), the methionine improved soybean which also possess a higher protein content provide a superior soybean meal to feed the livestock. In addition, the soybean cv. Zigongdongdou (ZD)is late-maturing and can maintain vegetative growth in long day condition or high altitude areas due to its sensitivity to photoperiod (Jiang et al. 2011), making the methionine-enhanced transgenic ZD be a high-quality green forage and a soil improving crop for agricultural and pastoral areas(Peoples et al. 1995). This could also ease the expanding demand for feed and bene fit famers.
The higher levels of soluble methionine in seeds of JX,unlike those of ZD, suggested that the excess methionine can be further incorporated into storage protein. To test this assumption, it will be worthwhile in the future, to express CGS in these seeds containing methionine-rich storage proteins. These can include the 10 kDa γ-zein protein that previously found to elevate the levels of methionine from 15.49 to 18.57% (Li et al. 2005) or the 15 kDa zein protein that increased by 12 to 20% of the methionine content(Dinkins et al. 2001). Expression of these two methionine rich proteins did not lead to signi ficantly higher levels of total methionine (Krishnana 2005), suggesting that the level of soluble methionine might limit the methionine-rich protein synthesis (Amir et al. 2012). According to our study, total methionine levels were increased to 1.5- to 4.8-fold, indicating that the bottleneck of the methionine elevation was the synthesis ‘source’ rather than the ‘sink’ in soybean seeds.Thus, increased methionine synthesis by expression AtDCGS ef ficiently promoted the accumulation of methionine in soybean. In addition, constitutive expressing of At-DCGS expanded the synthesis ‘source’ and led to signi ficant higher methionine in soybean leaves. It contributed to the accumulation of methionine in seeds and gives to higher methionine content than the soybean with limited expressing At-DCGS in seed only. Expanded synthesis source gives to higher accumulation of methionine in soybean seeds and leaves which provided not only superior feed as soymeal, but also the green forage for livestock. We also addressed that the de ficiency in methionine accumulation could limit protein accumulation in soybean seeds, and excess accumulation in methionine led to universal increased protein content in seeds. Hence, as the source, abundant synthesis of methionine is imperative not only to make up the de ficiency of methionine but also to improve the protein content in soybean which inspires us a new approach for improving soybean protein quality and quantity in further breeding process.
5. Conclusion
As one of the most important crops which possess high protein content for human and animal consuming, soybean could be used as superior plant origin protein source.However, the content of sulfur-containing amino acids limited the protein nutrition due to the necessity of such essential amino-acids. The AtD-CGS gene, which encodes a feedback-insensitive cystathionine γ-synthase, elevates the methionine content of soybeans. Constitutive overexpression of AtD-CGS increased methionine synthesis and promoted the accumulation of methionine in leaves and seeds. The transgenic soybeans also displayed increased total protein contents in leaves and seeds, which provided us a practicable strategy to improve methionine and protein contents in soybeans.
Acknowledgements
We gratefully acknowledge Miss Tao Jinlu and Yao Weiwei(CAAS) for their assistance in soybean transformation and neve filler from Migal Galilee Technology Center, Kiryat Shmona, Israel for assistance in amino acids analysis. This work was supported by the Major Science and Technology Projects of China (2014ZX08004-003, 2014ZX08010-004),the Natural Science Foundation of China (31471571), the earmarked fund for China Agriculture Research System(CARS-04), and the Agricultural Science and Technology Innovation Project of CAAS, as well as by Israel Science Foundation (ISF grant 231-09).
Appendixassociated with this paper can be available on http://www.ChinaAgriSci.com/V2/En/appendix.htm
Amir R. 2010. Current understanding of the factors regulating methionine content in vegetative tissues of higher plants.Amino Acids,39, 917–931.
Amir R, Han T, Ma F. 2012. Bioengineering approaches to improve the nutritional values of seeds by increasing their methionine content. Molecular Breeding,29, 915–924.
Amira G, Ifat M, Tal A, Hana B, Shmuel G, Rachel A. 2005.Soluble methionine enhances accumulation of a 15 kDa zein, a methionine-rich storage protein, in transgenic alfalfa but not in transgenic tobacco plants. Journal of Experimental Botany,56, 2443–2452.
Becker D, Kemper E, Schell J, Masterson R. 1992. New plant binary vectors with selectable markers located proximal to the left T-DNA border. Plant Molecular Biology,20,1195–1197.
Bourgis F, Roje S, Nuccio M L, Fisher D B, Tarczynski M C, Li C, Herschbach C, Rennenberg H, Pimenta M J, Shen T L,Gage D A, Hanson A D. 1999. S-methylmethionine plays a major role in phloem sulfur transport and is synthesized by a novel type of methyltransferase. The Plant Cell,11,1485–1498.
Chiba Y, Ishikawa M, Kijima F, Tyson R H, Kim J, Yamamoto A, Nambara E, Leustek T, Wallsgrove R M, Naito S. 1999.Evidence for autoregulation of cystathionine γ-synthase mRNA stability in Arabidopsis. Science,286, 1371–1374.
Cohen H, Israeli H, Matityahu I, Amir R. 2014. Seedspeci fic expression of a feedback-insensitive form of CYSTATHIONINE-γ-SYNTHASE in Arabidopsis stimulates metabolic and transcriptomic responses associated with desiccation stress. Plant Physiology,166, 1575–1592.
Cooper A J. 1983. Biochemistry of sulfur-containing amino acids. Annual Review of Biochemistry,52, 187–222.
Czuj T, Żuk M, Starzycki M, Amir R, Szopa J. 2009.Engineering increases in sulfur amino acid contents in flax by overexpressing the yeast Met25 gene. Plant Science,177, 584–592.
Dinkins R D, Srinivasa Reddy M, Meurer C A, Yan B, Trick H, Thibaud-Nissen F, Finer J J, Parrott W A, Collins G B. 2001. Increased sulfur amino acids in soybean plants overexpressing the maize 15 kDa zein protein. In Vitro Cellular & Developmental Biology-Plant,37, 742–747.
Fehr W R, Caviness C E. 1977. Stages of Soybean Development.Special Report No. 80. Cooperative Extension Service,Agriculture and Home Economic Experiment Station, Ames,Iowa, Iowa State University, USA.
Frank A, Cohen H, Hoffman D, Amir R. 2015. Methionine and S-methylmethionine exhibit temporal and spatial accumulation patterns during the Arabidopsis life cycle.Amino Acids,47, 497–510.
Gallardo K, Firnhaber C, Zuber H, Héricher D, Belghazi M, Henry C, Küster H, Thompson R. 2007. A combined proteome and transcriptome analysis of developing Medicago truncatula seeds evidence for metabolic specialization of maternal and filial tissues. Molecular & Celluar Proteomics,6, 2165–2179.
Hacham Y, Matityahu I, Schuster G, Amir R. 2008.Overexpression of mutated forms of aspartate kinase and cystathionine γ-synthase in tobacco leaves resulted in the high accumulation of methionine and threonine. The Plant Journal,54, 260–271.
Hacham Y, Schuster G, Amir R. 2006. An in vivo internal deletion in the N-terminus region of Arabidopsis cystathionine gamma-synthase results in CGS expression that is insensitive to methionine. The Plant Journal,45, 955–967.
Hacham Y, Song L, Schuster G, Amir R. 2007. Lysine enhances methionine content by modulating the expression of S-adenosylmethionine synthase. The Plant Journal,51,850–861.
Hanafy M S, Rahman S M, Nakamoto Y, Fujiwara T, Naito S, Wakasa K, Ishimoto M. 2013. Differential response of methionine metabolism in two grain legumes, soybean and azuki bean, expressing a mutated form of Arabidopsis cystathionine γ-synthase. Journal of Plant Physiology,170,338–345.
Hernández-Sebastià C, Marsolais F, Saravitz C, Israel D,Dewey R E, Huber S C. 2005. Free amino acid pro files suggest a possible role for asparagine in the control of storage-product accumulation in developing seeds of lowand high-protein soybean lines. Journal of Experimental Botany,56, 1951–1963.
Holowach L P, Thompson J F, Madison J T. 1984. Effects of exogenous methionine on storage protein composition of soybean cotyledons cultured in vitro. Plant Physiology,74,576–583.
Jiang Y, Wu C, Zhang L, Hu P, Hou W, Zu W, Han T. 2011.Long-day effects on the terminal in florescence development of a photoperiod-sensitive soybean [Glycine max (L.) Merr.]variety. Plant Science,180, 504–510.
Kim E H, Ro H M, Kim S L, Kim H S, Chung I M. 2012.Analysis of iso flavone, phenolic, soyasapogenol, and tocopherol compounds in soybean [Glycine max (L.) Merrill]germplasms of different seed weights and origins. Journal of Agricultural and Food Chemistry,60, 6045–6055.
Kingston-Smith A H, Marshall A H, Moorby J M. 2013. Breeding for genetic improvement of forage plants in relation to increasing animal production with reduced environmental footprint. Animal,7, 79–88.
Krishnan H B. 2005. Engineering soybean for enhanced sulfur amino acid content. Crop Science,45, 454–461.
Lee M, Huang T, Toro-Ramos T, Fraga M, Last R L, Jander G. 2008. Reduced activity of Arabidopsis thaliana HMT2,a methionine biosynthetic enzyme, increases seed methionine content. The Plant Journal,54, 310–320.
Li Z, Meyer S, Essig J S, Liu Y, Schapaugh M A, Muthukrishnan S, Hainline B E, Trick H N. 2005. High-level expression of maize γ-zein protein in transgenic soybean (Glycine max).Molecular Breeding,16, 11–20.
Matityahu I, Godo I, Hacham Y, Amir R. 2013. Tobacco seeds expressing feedback-insensitive cystathionine gammasynthase exhibit elevated content of methionine and altered primary metabolic pro file. BMC Plant Biology,13, 206.
Mattoo A K, Shukla V, Fatima T, Handa A K, Yachha S K. 2010.Genetic engineering to enhance crop-based phytonutrients(nutraceuticals) to alleviate diet-related diseases. Bio-Farms for Nutraceuticals (Springer US),698, 122–143.
Nguyen H C, Hoefgen R, Hesse H. 2012. Improving the nutritive value of rice seeds: Elevation of cysteine and methionine contents in rice plants by ectopic expression of a bacterial serine acetyltransferase. Journal of Experimental Botany,63, 5991–6001.
Ominato K, Akita H, Suzuki A, Kijima F, Yoshino T, Yoshino M,Chiba Y, Onouchi H, Naito S. 2002. Identi fication of a short highly conserved amino acid sequence as the functional region required for posttranscriptional autoregulation of the cystathionine γ-synthase gene in Arabidopsis. Journal of Biological Chemistry,277, 36380–36386.
Peoples M, Ladha J, Herridge D. 1995. Enhancing legume N2 fixation through plant and soil management. Plant and Soil,174, 83–101.
Ranocha P, McNeil S D, Ziemak M J, Li C, Tarczynski M C, Hanson A D. 2001. The S-methylmethionine cycle in angiosperms, ubiquity, antiquity and activity. The Plant Journal,25, 575–584.
Shewry P R. 2000. Seed proteins. In: Black M, Bewley J D,eds., Seed Technology and its Biological Basis. Shef field Academic Press, Shef field. pp. 42–84.
Song S, Hou W, Godo I, Wu C, Yu Y, Matityahu I, Hacham Y,Sun S, Han T, Amir R. 2013. Soybean seeds expressing feedback-insensitive cystathionine γ-synthase exhibit a higher content of methionine. Journal of Experimental Botany,64, 1917–1926.
Sunilkumar G, Mohr L, Lopata-Finch E, Emani C, Rathore K S. 2002. Developmental and tissue-speci fic expression of CaMV 35S promoter in cotton as revealed by GFP. Plant Molecular Biology,50, 463–479.
WHO (World Health Organization). 2007. Protein and Amino Acid Requirements in Human Nutrition. Report of A Joint WHO/FAO/UNU Experts Conclusion. World Health Organization Press, Geneva.
Yu J, Peng P, Zhang X, Zhao Q, Zhy D, Sun X, Liu J, Ao G. 2004.Seed-speci fic expression of a lysine rich protein sb401 gene signi ficantly increases both lysine and total protein content in maize seeds. Molecular Breeding,14, 1–7.
12 December, 2016 Accepted 20 February, 2017
Correspondence Rachel Amir, E-mail: Rachel@migal.org.il; HAN Tian-fu, Tel: +86-10-82105875, Fax: +86-10-8210878, E-mail:tianfuhan@hotmail.com, hantianfu@caas.cn
*These authors contributed equally to this study.
© 2018 CAAS. Publishing services by Elsevier B.V. All rights reserved.
10.1016/S2095-3119(16)61599-X
Managing editor WANG Ning
杂志排行

Journal of Integrative Agriculture的其它文章
- Climate change and agriculture: Impacts and adaptive responses in Iran
- Development of elite restoring lines by integrating blast resistance and low amylose content using MAS
- Evaluation of stability and yield potential of upland rice genotypes in North and Northeast Thailand
- ldenti fication of the resistance gene to powdery mildew in Chinese wheat landrace Baiyouyantiao
- New clues concerning pigment biosynthesis in green colored fiber provided by proteomics-based analysis
- Characterization of groundnut (Arachis hypogaea L.) collection using quantitative and qualitative traits in the Mediterranean Basin