啮齿动物对不同林木种子的搬运和取食微生境选择机制
2018-01-03康海斌王得祥常明捷胡有宁滕青林
康海斌,王得祥,*,常明捷,康 冰,胡有宁,4,于 飞,5,庞 越,滕青林
1 西北农林科技大学林学院,杨凌 712100 2 甘肃小陇山森林生态系统国家定位观测研究站,天水 741020 3 西北农林科技大学生命科学学院,杨凌 712100 4 西安文理学院生物与环境工程学院,西安 710065 5 河南师范大学生命科学学院,新乡 453007
啮齿动物对不同林木种子的搬运和取食微生境选择机制
康海斌1,2,王得祥1,2,*,常明捷1,康 冰3,胡有宁1,4,于 飞1,5,庞 越1,滕青林1
1 西北农林科技大学林学院,杨凌 712100 2 甘肃小陇山森林生态系统国家定位观测研究站,天水 741020 3 西北农林科技大学生命科学学院,杨凌 712100 4 西安文理学院生物与环境工程学院,西安 710065 5 河南师范大学生命科学学院,新乡 453007
不同植物种子依靠不同的方式实现扩散,啮齿动物对林木种子搬运后在取食点微生境和贮藏方式的选择存在偏好,研究其贮藏行为与微生境的关系是探究幼苗建成的关键。在秦岭中段火地塘林区,采用标签标记法,以锐齿槲栎、华山松和油松种子为材料,探究了小型啮齿动物对松栎混交林建群种种子扩散过程的影响。结果表明:1)油松种子原地取食率显著高于锐齿槲栎和华山松种子,且啮齿动物更倾向于搬运后取食(60%)和埋藏(4.33%)华山松种子,搬运后取食距离也为华山松最大(2.49 m);锐齿槲栎小种子被搬运后埋藏的距离最大(4.92 m)。2)除华山松种子外,其他类型种子被搬运后单个取食的比例均在85%以上;油松种子不存在埋藏点,而其他类型种子90%以上均以单个形式被埋藏。3)大部分种子被啮齿动物搬运后选择在裸地丢弃;锐齿槲栎大种子(87.5%)、小种子(78.57%)和华山松种子(53.33%)较大比例被啮齿动物埋藏在灌丛下方,埋藏在裸地的种子较少。4)大部分种子在灌丛下方被取食,仅华山松种子被啮齿动物搬运到洞穴取食;除油松种子被大量原地取食外,其他类型种子被搬运到取食点的种子比例基本呈现由微生境植被复杂到简单(灌丛—草丛—灌丛边缘—裸地)而逐渐减小的趋势。种子的营养价值及取食和搬运过程中啮齿动物付出的成本是影响种子命运的关键性因子,且啮齿动物对种子埋藏和取食地点的微生境存在较明显的选择性。
小型啮齿动物;种子扩散;觅食对策;微生境;松栎混交林;秦岭
动物在森林群落种子扩散和更新过程中扮演着较为重要的角色[1-2],林内的啮齿动物既能通过贮食行为参与植物种子扩散和实生幼苗的建成[3],又会因为大量取食种子而影响植物的天然更新,其和植物构成了一个包含捕食和互惠作用的系统[4]。植物种子被动物搬运和埋藏后,可以远离母树,有效避免与母树的竞争、避免病原菌以及幼苗间竞争,还可以使种子有更多的机会扩散到适合其萌发的定居地点[5-6]。之前的研究多集中在动物贮食行为对植物种子扩散以及林分生境类型对种子命运的影响[7],很少深入涉及动物的埋藏特征和对取食种子点微生境的选择及其微生境与植物种子萌发和幼苗建成的关系。但是,上述问题的研究却往往是揭示动物传播种子对植物天然更新影响的关键,而不同大小的同种植物种子以及不同植物种子的命运是了解森林群落更新很好的切入点。
啮齿动物对较大且具有较高营养价值的种子有一定的取食和贮藏偏好[6, 8-9]。动物在觅食时会权衡投入成本(时间和能量)而做出不同的取食和扩散对策[10],表现为对于种皮较坚硬的种子倾向于搬运后取食和贮藏,而喜欢就地消费种皮薄的种子[11]。另外,季节变化[12-13]、种子品质的好坏[14-15]、次级代谢物[9]、灌丛高度[16]、种子挥发物的多少[17]等也会影响啮齿动物对种子的取食和扩散。可见,啮齿动物的贮食行为受到诸多因素的综合影响,其扩散模式的形成是一个复杂的过程。啮齿动物通过这种扩散模式将植物种子扩散到适合种子萌发的地点,不同类型种子选择不同的埋藏点和微生境,形成植物天然更新的分布格局[18-19]。
锐齿槲栎(Quercusalienavar.acuteserrata)、油松(Pinustabuliformis)和华山松(Pinusarmandii)在秦岭南坡分布十分广泛,是秦岭山地次生林的主要建群种,种子雨均发生于9—10月。锐齿槲栎种子大小差异较大,含有淀粉和单宁。华山松种子富含脂肪、蛋白质等,外种皮较厚且坚硬。油松种子相对较小,外种皮较薄。在成熟期这些种子均是小型啮齿类动物的主要食物资源,且受到不同程度的取食与搬运。该地区锐齿槲栎植株结实量有限,存在年际间的差异;相反,华山松和油松等树种结实量均较大,基本可以维持群落内啮齿动物种群的稳定。已有不少研究围绕啮齿动物对秦岭植物群落中种子的取食和扩散开展,但针对松栎混交林中啮齿动物与搬运和取食点微生境之间关系的研究相对较少。张博等[14]在秦岭南坡调查了鼠类在食物丰富度影响下对完好和虫蛀锐齿槲栎种子的选择差异,发现食物丰富的秋季鼠类偏好贮藏完好种子,在食物匮乏的春季,鼠类更倾向于同时取食两种类型种子。于飞等[20]通过研究林窗干扰下锐齿槲栎和华山松种子扩散过程,结果表明微生境条件对秦岭林区锐齿槲栎和华山松种子扩散和存活具有显著影响,啮齿动物为其林窗更新发挥了积极作用。针对于如何通过啮齿动物的觅食行为来选择出适合林木种子萌发的微生境,来指导秦岭地区森林恢复,达到保护生态系统的现实问题。本研究于2015年9—10月在秦岭南坡火地塘林区松栎混交林内,通过释放锐齿槲栎大小种子、华山松和油松种子,探讨啮齿动物觅食行为影响下各类种子命运及其搬运和取食种子地点微生境的选择,旨在阐明啮齿动物对该区域松栎混交林建群种种子扩散的影响和其对森林更新的作用,以期进一步明确动、植物间的相互关系,为揭示松栎混交林的天然更新和演替机制提供科学依据。
1 研究地区与研究方法
1.1 研究地点概况
试验研究于陕西秦岭南坡中段火地塘林区进行,火地塘(108°25′—108°30′E, 33°25′—33°29′N)地处秦岭南坡中部,位于中国北亚热带和暖温带的过渡地带,海拔1450—2473 m,年平均气温8—10℃,年降水量1000—1200 mm,年蒸发量800—950 mm,年日照时数1100—1300 h,无霜期170 d。土壤类型主要以黄棕壤、棕壤、暗棕壤及草甸土为主,土层厚度45—105 cm,pH值5.14—5.81,偏酸性。该林区内的森林植被属温带针阔混交林和寒带针叶林,森林覆盖面积达93.8%,主要成林树种有锐齿槲栎、油松、华山松、铁杉(Tsugachinensis)、红桦(Betulaalbo-sinensis)等。该区域在20世纪60—70年代进行了全面的森林主伐,现在森林植被基本已得到较好的恢复,以天然次生林为主,郁闭度在0.9以上[21]。该地区分布的啮齿动物主要有大林姬鼠(Apodemuspeninsulae)、中华姬鼠(Apodemusdraco)、社鼠(Niviventerconfucianus)、岩松鼠(Sciurotamiasdavidianus)等。
1.2 研究方法
1.2.1种子收集、标记和释放
锐齿槲栎种子成熟时,直接从母树上采摘或者收集散落于林冠层下的种子,并利用水浮法筛选出健康完好的种子,共筛选出1000粒,由于其形状多为卵形,且大小差异较大,将筛选出的1000粒锐齿槲栎种子分为大橡子((1.73±0.04) g,mean ± SE,n=100)和小橡子((0.74±0.02) g,n=100)。华山松和油松种子均购自当地于2014年秋季采集的种子,大小分别为((0.27±0.01) g,n=100)和((0.02±0.001) g,n=100)。采用塑料标签标记法进行种子标记[4,22],在种子一端用微型电钻钻一直径0.3mm的小孔,再用长12cm、直径0.3mm的细铁丝将其与白色塑料标签(3.0cm×2.0cm,<0.1g)相连,每个标签上记录对应种子释放点、种子类别及种子序号等信息,以便后续对种子进行追踪和调查。小型啮齿动物取食种子或进行搬运埋藏后,标签通常会留在地面,因此,在试验开始后,可通过搜寻标签来确定每一粒种子的命运。
于2015年10月在典型松栎混交林中设置4条平行的样线进行种子释放。样线间隔20m,每条样线上,间隔20m设置一个种子释放点,面积为1m2,共计10个,4条样线共设置释放点40个。4条样线依次单独放置锐齿槲栎大种子、锐齿槲栎小种子、华山松种子和油松种子,每个释放点放置种子30粒,成列均匀放置,共计释放种子4(样线)×10(释放点)×30粒=1200粒。
1.2.2种子追踪
从试验开始,分别在第2、4、6、8、10天检查种子释放点,并在释放点20m直径范围内寻找被扩散的种子并详细记录其种子命运:原地剩余、原地取食、搬运后取食、搬运后丢弃、搬运后埋藏和丢失;调查记录搬运后丢弃点、取食点和埋藏点的微生境和种子贮藏方式(土壤埋藏和洞穴贮藏)。对搬运后的种子记录其编号,并测量其搬运距离(与对应释放点的距离)。
1.2.3啮齿动物群落结构调查
本研究采用活捕笼法(27cm×14cm×11cm)[23],旨在了解研究区内小型啮齿动物的种类组成,未进行种群的大小和密度的相关调查。选择在种子扩散试验结束后,在试验区设置2条间隔100m的样线,每条样线上间隔5m放置活捕笼1个,每条样线放置15个,共计30个。以花生和香肠为诱饵,调查开始当天下午置笼,第二天检查,共调查5d,累计150个活捕笼日(15活捕笼×2样线×5天),对被捕获的动物进行称重、拍照后原地释放。捕获率=捕获数/总笼数。
1.3 数据处理
采用SPSS21.0软件对有关数据进行统计分析。计算不同种子命运的百分比、种子被搬运后(取食、丢弃或贮藏)的距离及其分布频次,并统计不同种子被啮齿动物搬运后丢弃点、埋藏点和取食所处地的微生境。以Cox回归法分析4种类型种子扩散速度的差异。通过General Linear Model分析不同类型种子命运之间的差异。所有数据均以平均值±标准误差表示。所有图表均在Origin8.1中完成。
2 结果与分析
2.1 种子存留动态
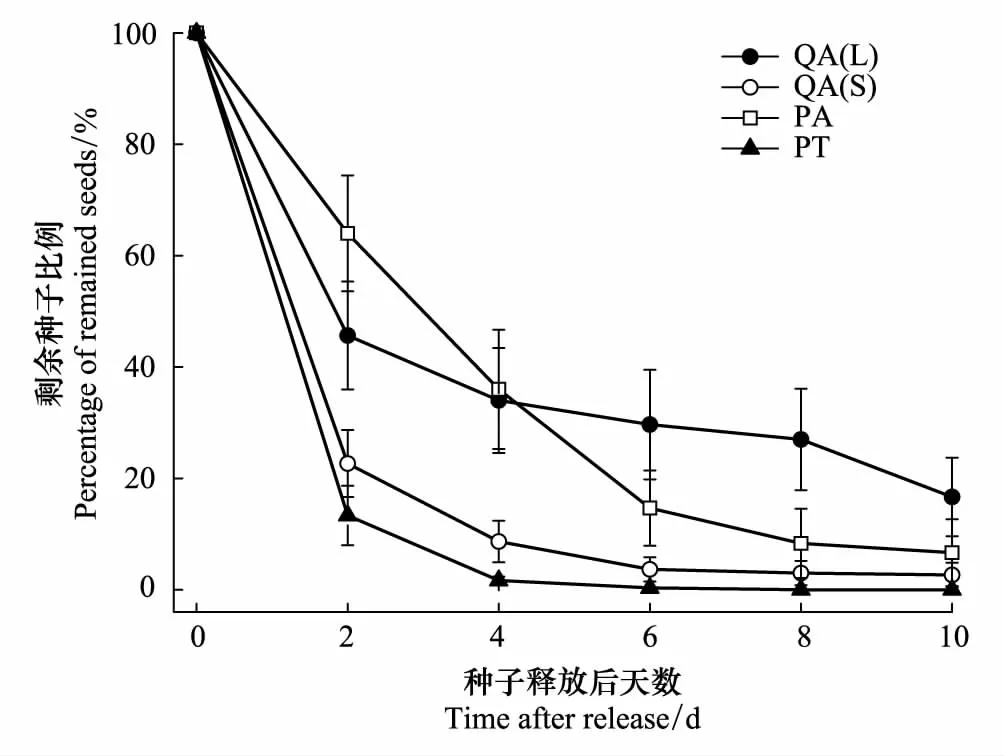
图1 种子释放点4种类型种子存留动态 Fig.1 Seed removal rates of four types of seeds after being place at seed sites in the study area. Data are expressed as mean ± SE.QA(L):锐齿槲栎(大) Quercus aliena var. acuteserrata (large);QA(S):锐齿槲栎(小)Quercus aliena var. acuteserrata (small);PA:华山松 Pinus armandii;PT:油松 Pinus tabuliformis
4种类型种子存留动态如图1所示。截止试验调查结束,油松种子被完全扩散,扩散速度最快;依次为锐齿槲栎小种子、华山松种子、锐齿槲栎大种子;其中,锐齿槲栎大种子表现为前期扩散较快,后期较缓慢。Cox回归分析表明,啮齿动物对4种种子的扩散速度存在显著差异(Wald=12.443,P=0.005)。
2.2 种子命运
General Linear Model分析结果显示(图2),不同类型种子被啮齿动物原地取食(F=33.947,P<0.0001)、搬运后取食(F=8.200,P<0.0001)和丢失比例(F=3.387,P=0.028)均存在显著差异,相反,种子原地剩余(F=2.078,P=0.12)、搬运后埋藏(F=0.986,P=0.41)和搬运后丢弃(F=0.881,P=0.46)种子比例在不同类型种子间无显著差异。
其中,油松种子被啮齿动物原地取食比例显著高于其他三种类型种子(P<0.01),另锐齿槲栎大种子(P<0.01)和小种子(P<0.05)的原地取食率均显著高于华山松种子;华山松种子被啮齿动物搬运后取食的比例显著高于锐齿槲栎大种子(P<0.01)、小种子(P<0.05)和油松种子(P<0.01),而锐齿槲栎小种子搬运后的取食率也显著高于油松种子(P<0.01);锐齿槲栎大种子(P<0.05)、小种子(P<0.01)和华山松种子(P<0.01)的丢失率均显著高于油松种子。种子被原地取食率以油松最高(83.33%),远大于锐齿槲栎大种子(36.67%)、小种子(32%)和华山松(9%);而华山松种子被搬运后取食的比例最高(60%),油松种子最低(15.33%)。

图2 研究区4种类型种子扩散后的命运Fig.2 Fates of four types of seeds after dispersal by small rodents in the study area. Data are expressed as mean ± SE
2.3 种子扩散距离
研究区4种类型种子被啮齿动物搬运后取食的平均距离(图3)以华山松种子最大(2.49m),油松最小(0.20m);华山松种子被啮齿动物搬运后丢弃的距离最大(2.45m),其他依次为锐齿槲栎小种子(1.06m)、锐齿槲栎大种子(0.25m)和油松种子(0.23m);锐齿槲栎大种子、锐齿槲栎小种子、华山松种子和油松种子被啮齿动物搬运后埋藏距离分别是2.39、4.92、2.38、0m。啮齿动物更喜欢将华山松种子搬运至较远的地方进行取食,而对油松种子基本采取就近消费的方式,对锐齿槲栎小种子倾向于搬运较远的距离后进行埋藏。

图3 研究区4种类型种子平均和最大扩散距离Fig.3 Average and maximum seed dispersal distance of four types of seeds from the seed release sites in the study area
种子扩散距离的分布频次结果表明(图4),锐齿槲栎大种子、小种子和华山松种子绝大多数的扩散距离都在10m以下,但油松种子的扩散距离均处于1m以下的水平。锐齿槲栎大种子随着扩散距离增大被扩散的种子比例逐渐减少,主要集中在1m以下(58.87%),其他依次为1—3m、3—5m、5—8m和>10m(分别为26.61%、8.87%、4.84%和0.81%),在8—10m范围没有分布;锐齿槲栎小种子在所有距离区组均有分布,同样主要集中在1m以下(61.16%),在1—3m(19.83%)和3—5m(12.4%)也有较多的分布,在8—10m分布频次最低(0.83%);华山松种子主要集中在1—3m、<1m和5—8m距离区组(分别为39.9%、30.54%和18.23%),在>10m距离区组分布频次最低,为0.49%。
2.4 取食和埋藏点的种子数量
锐齿槲栎大种子、小种子、华山松和油松种子大部分表现为搬运后单个取食(分别为85.71%、92.55%、72.37%和87.5%),其中仅华山松种子存在含有5个以上的取食点,含有7个种子以上的取食点均只有1个,占全部取食点的1.3%;锐齿槲栎大种子和小种子均表现为单一埋藏点,油松种子无埋藏点,华山松种子主要为单一埋藏点(91.7%),含有2个种子的埋藏点仅占8.3%。
2.5 种子被搬运后丢弃和埋藏点微生境
根据调查统计种子被啮齿动物搬运后丢弃点的微生境结果如图5所示。大部分种子被啮齿动物搬运后在裸地丢弃。锐齿槲栎大种子被搬运后主要丢弃在裸地(89.47%);锐齿槲栎小种子被丢弃在裸地、草丛和灌丛下方的比例相等(33.33%);华山松种子被丢弃在灌丛下方的比例最大(44.44%),其次为草丛(33.33%),灌丛边缘和裸地相等(11.11%);油松种子主要被丢弃在裸地(66.67%)和灌丛下方(33.33%)。
表1研究区4种类型种子被啮齿动物扩散后不同大小取食点和埋藏点分布比例/%
Table1Distributionpercentageofdifferentsizesofpredationandcachesitesoffourtypesofseedsafterdispersalbyrodentsinthestudyarea
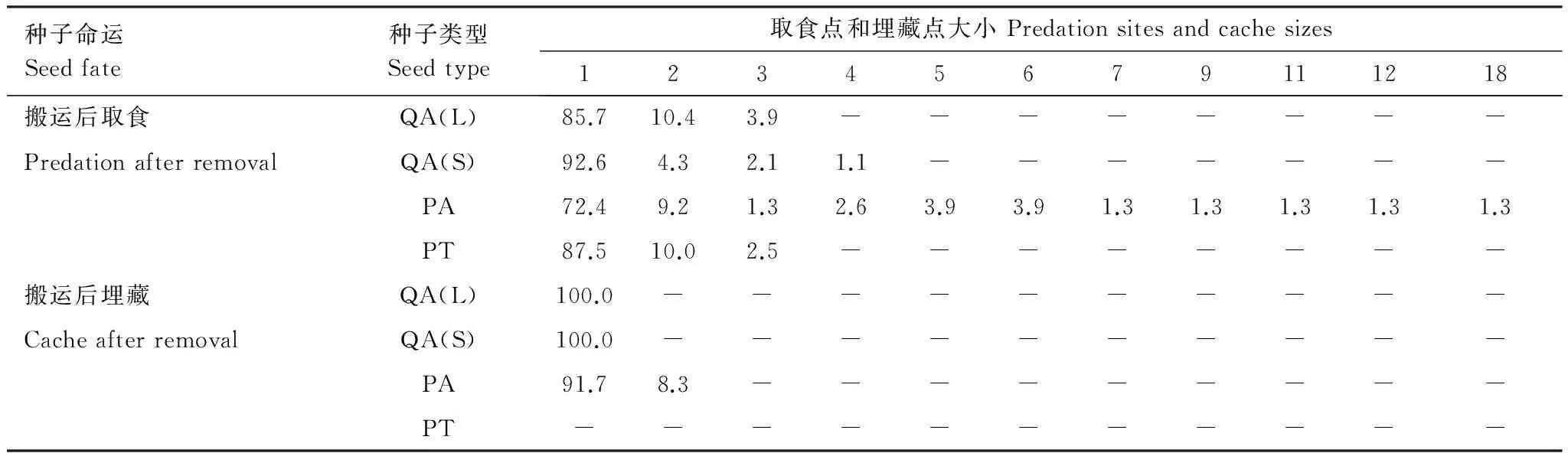
种子命运Seedfate种子类型Seedtype取食点和埋藏点大小Predationsitesandcachesizes12345679111218搬运后取食QA(L)85.710.43.9--------PredationafterremovalQA(S)92.64.32.11.1-------PA72.49.21.32.63.93.91.31.31.31.31.3PT87.510.02.5--------搬运后埋藏QA(L)100.0----------CacheafterremovalQA(S)100.0----------PA91.78.3---------PT-----------
种子被啮齿动物搬运后埋藏点的微生境统计如图6所示。除油松种子不存在埋藏点外,其他3种类型种子大部分种子被啮齿动物选择埋藏在灌丛下方,另华山松被埋藏的微生境更为多样化。锐齿槲栎大种子主要被埋藏在灌丛下方(87.5%)和洞穴中(12.5%);锐齿槲栎小种子被埋藏在灌丛下方最多(78.57%),依次为草丛(14.29%)、灌丛边缘(7.14%),华山松种子被埋藏在灌丛下方比例较大(53.33%),选择草丛、灌丛边缘和洞穴埋藏的比例均相等(13.33%),埋藏在裸地最少(6.67%)。
2.6 种子搬运后取食点微生境
种子被啮齿动物搬运后取食点的微生境统计如图7所示。4种类型种子主要被集中在灌丛下方取食,仅华山松种子被啮齿动物搬运到洞穴取食。其中,锐齿槲栎大种子大部分被搬运至灌丛下方取食(69.23%),被搬运到草丛取食的比例次之(14.29%),被搬运到裸地和灌丛边缘的种子较少(分别为9.89%和6.59%);锐齿槲栎小种子主要被搬运到灌丛下方和裸地进行了取食(分别为37.21%和29.46%),其次为草丛(21.71%)和灌丛边缘(11.63%);华山松种子大多数被搬运至灌丛下方取食(77.27%),被啮齿动物搬运至洞穴取食的种子比例次之(14.65%),其余3种取食点所占比例均较小,分别为草丛(3.54%)、灌丛边缘(3.03%)和裸地(1.52%);油松种子被搬运到灌丛边缘取食的比例最大(40.43%),依次为裸地(31.91%)、草丛(21.28%),搬运至灌丛下方取食的比例最小(6.38%)。

图4 研究区4种类型种子扩散距离的分布频次Fig.4 Distribution frequency of the seed dispersal distance of four types of seeds from the seed release sites in the study area

图5 研究区4种类型种子被啮齿动物搬运后丢弃点微生境Fig.5 Discarding microhabitat of four types of seeds after removal by rodents in the study areaBL: 裸地Bare land; SE: 灌丛边缘Shrub edge; G: 草丛Grass; US: 灌丛下方Under shrub

图6 研究区4种类型种子被啮齿动物搬运后埋藏点微生境 Fig.6 Cache microhabitat of four types of seeds after removal by rodents in the study areaH: 洞穴Hole

图7 研究区4种类型种子被啮齿动物搬运后取食点的微生境 Fig.7 Predation microhabitat of four types of seeds after removal by rodents in the study area
2.7 小型啮齿动物群落结构
试验区共捕获小型啮齿动物3种,共51只(表2),分别为大林姬鼠、中华姬鼠和岩松鼠,总捕获率为34%,主要以大林姬鼠居多,其次为中华姬鼠和岩松鼠,调查期间多次见到松鼠及其活动的痕迹。

表2 啮齿动物的种类组成及捕获率
3 讨论
啮齿动物对不同植物种子存在一定程度上的行为分化,其取食和贮藏种子的行为受到种子大小、形状、种皮(果皮)厚度、营养成分、昆虫侵染等因素的影响[14, 24-25]。大量研究表明,啮齿动物对不同大小的种子取食和扩散模式存在差异,倾向于取食和搬运较大且具有较高营养价值的种子以获取更多的能量净收益[1, 6, 9],在遇到具有厚而坚硬种皮的种子往往为了避免在取食过程中由于捕食时间过长而带来较大的风险,会采取贮藏或者搬运到安全地点取食的策略,即啮齿动物更倾向于就地取食种皮较薄的种子,贮藏种皮较厚且坚硬的种子[11, 26]。本研究发现,油松种子和锐齿槲栎小种子的扩散速度较华山松和锐齿槲栎大种子快;啮齿动物存在就地取食油松种子的偏好,而倾向于将具有较厚且坚硬种皮的华山松种子搬运到合适的取食地点进行消费,这是由于啮齿动物在捕食华山松种子时需要处理的时间更长,其本身被捕食的风险增大,导致啮齿动物做出一定的觅食对策。华山松种子被搬运后埋藏的比例也较大,这可能与华山松种子具有较高的营养价值有关,而锐齿槲栎种子含有大量的次级代谢化合物单宁。有研究表明单宁含量较高的种子可能不利于啮齿动物对蛋白质的吸收,甚至具有潜在的毒性作用,动物在捕食后可能会出现肝肾衰竭、体内氮流失等症状[27],肖治术等[28]通过研究四川都江堰地区的小泡巨鼠对森林种子的选择和贮藏发现,小泡巨鼠喜好取食和贮藏单宁含量较低的种子,而不喜欢取食和贮藏单宁含量较高的种子。因此,啮齿动物更愿意花费较多的觅食时间和精力去贮藏高脂肪含量的华山松种子,以供后续生存和繁殖消费。
对食物资源的贮藏距离啮齿动物会根据食物特征和竞争者做出一定决策,竞争者的存在加强了动物的贮食行为,也加快了其对资源的占有[29]。研究发现,锐齿槲栎、华山松种子和少量油松种子被搬离种子资源点(种子释放点)均保持在5 m范围内,扩散速度均表现为前期扩散较快,后期较缓慢的趋势。华山松种子被搬运取食的距离显著大于其他3种类型种子,充分说明了啮齿动物在处理种皮较厚且坚硬的种子时会尽可能增大搬运距离,以寻找可降低被捕食风险的安全地点进行取食。啮齿动物搬运后埋藏距离的结果并没有表现为锐齿槲栎大种子大于锐齿槲栎小种子,相反,锐齿槲栎小种子的搬运埋藏距离却大于锐齿槲栎大种子。结合最优贮藏空间模型[30],营养价值较高的食物被贮藏在远离资源的地点,而营养价值较低的食物被贮藏在资源附近。焦广强[9]对人工种子的试验发现,啮齿动物喜欢原地取食单宁含量高的种子,偏好扩散单宁含量低的种子,且埋藏率较高,在扩散的距离上,单宁含量低的种子扩散距离比单宁含量高的种子远。这些研究与本研究结果基本一致。另外,大种子同样具有较厚的种皮和质量,这些因素可能会造成啮齿动物在搬运过程中付出更多的搬运成本,减缓搬运速度而增加被捕食风险。因此,啮齿动物可能更倾向于少量多次地搬运和埋藏小种子。
同域分布的食果实动物,由于利用食物资源的方式相同,同种或异种个体间在食物资源利用上存在明显的竞争[9]。干扰者和竞争者的存在会影响种子扩散过程,干扰者的存在就意味着食物资源可能被干扰者窃取,为了保护已有食物,动物发展和进化了应对策略,改变食物贮藏策略和行为可能是其中的重要方面[27],分散贮藏食物恰恰就是动物一种重要的觅食对策[9, 31]。本研究中4种类型种子的取食点和贮藏点基本保持在一个种子的水平,锐齿槲栎大种子和小种子均表现为单一分散埋藏的模式,华山松种子的单一贮藏点也占到90%以上。这种贮藏策略可以有效地降低食物被竞争者盗取的风险来确保其自身的利益。另外,本研究结果还显示,华山松种子存在较大的取食点,甚至达到18个种子之多,这些较大取食点的存在暗示着啮齿动物更倾向于将种子多次搬运至同一地点进行消费,这种地点可能是其认为较为安全或者熟悉的地点,如岩石缝隙、洞穴及其附近较为隐蔽的灌丛等。
种子扩散过程存在显著的空间差异,复杂的植被状况可以有效降低啮齿动物的觅食风险,啮齿动物在觅食时通常会权衡捕食风险和竞争压力[32-33]。啮齿动物搬运种子丢弃点的微生境结果表明,大部分种子被啮齿动物选择在裸地丢弃,这可能与在较空旷的地点进行埋藏和取食风险较大有关[34],而空旷的生境更有利于种子的萌发[35]。对埋藏点微生境的选择,啮齿动物倾向于在灌丛下方埋藏种子。本研究进一步支持之前的发现[16]。研究同样也发现有较大比例种子被埋藏在草丛和灌丛边缘。张知彬[36]研究发现较草地和裸地微生境相比,灌丛边缘与母体或其他植物的竞争较弱,且易接受较多光照,土壤中的水分和营养物质较多,有利于埋藏的种子萌发,相反,虽然灌丛下方的营养物质更为丰富,但其种间竞争较为激烈,因此也不利于种子的萌发。种子被搬运后取食点微生境的调查结果表明,林内的灌丛下方是啮齿动物取食种子的主要场所,对具有较厚且坚硬种皮的华山松种子大部分选择搬运至灌丛下方和洞穴取食。除油松种子被大量原地取食外,其他3种类型种子被搬运到取食点的种子比例基本呈现从微生境植被复杂到简单(灌丛—草丛—灌丛边缘—裸地)而逐渐减小的趋势,避免了啮齿动物在相对开阔的生境取食而被捕食者更容易发现,在一定程度上为啮齿动物提供了有利的遮蔽条件。可见,啮齿动物会有目的地将食物搬运至不同微生境下埋藏和取食,而并非随机搬运,而且灌丛这种埋藏和取食模式同时可以为种子的萌发创造较好的生境条件,起到一定的积极作用,促进植物的天然更新。
综上所述,影响种子命运的关键性因子更倾向于种子的营养价值及取食和搬运过程中啮齿动物付出的成本,而啮齿动物对埋藏和取食种子的地点的微生境存在较明显的选择性。秦岭南坡松栎混交林建群种的种子扩散规律存在差异,啮齿动物对油松种子的捕食压力较大,而锐齿槲栎和华山松的种子被选择性地扩散至适宜地点埋藏。由于枯落物厚度、光照和水分等微生境条件是种子萌发和幼苗成功定居的关键性因素,啮齿动物的扩散行为为种子萌发和幼苗的构建提供了有利条件,在不同程度上促进了锐齿槲栎和华山松种群的天然更新。因此,研究啮齿动物贮藏行为与微生境的关系是探究幼苗建成的关键,通过借助啮齿动物的觅食行为来寻求适宜种子萌发的微生境,可以更好地为森林生态系统的保护和恢复提供服务。
[1] Vander Wall S B. Food Hoarding in Animals. Chicago: University of Chicago Press, 1990.
[2] Perea R, Miguel A S, Gil L. Flying vs. climbing: factors controlling arboreal seed removal in oak-beech forests. Forest Ecology and Management, 2011, 262(7): 1251- 1257.
[3] Vander Wall S B, Joyner J W. Recaching of Jeffrey pine (Pinusjeffreyi) seeds by yellow pine chipmunks (Tamiasamoenus): potential effects on plant reproductive success. Canadian Journal of Zoology, 1998, 76(1): 154- 162.
[4] Li H J, Zhang Z B. Effect of rodents on acorn dispersal and survival of the Liaodong oak (QuercusliaotungensisKoidz.). Forest Ecology and Management, 2003, 176(1/3): 387- 396.
[5] Briggs J S, Vander Wall S B, Jenkins S H. Forest rodents provide directed dispersal of Jeffrey pine seeds. Ecology, 2009, 90(3): 675- 687.
[6] Yu F, Wang D X, Yi X F, Shi X X, Huang Y K, Zhang H W, Zhang X P. Does animal-mediated seed dispersal facilitate the formation ofPinusarmandii-Quercusalienavar.acuteserrataforests? PLoS One, 2014, 9(2): e89886.
[7] 闫兴富, 周立彪, 刘建利. 啮齿动物捕食压力下生境类型和覆盖处理对辽东栎种子命运的影响. 生态学报, 2012, 32(9): 2778- 2787.
[8] Smith C C, Reichman O J. The evolution of food caching by birds and mammals. Annual Review of Ecology and Systematics, 1984, 15(1): 329- 351.
[9] 焦广强. 大小、形状及单宁含量对人工种子扩散和命运的影响[D]. 洛阳: 河南科技大学, 2011.
[10] 周立彪, 闫兴富, 王建礼, 周云峰. 啮齿动物对不同大小和种皮特征种子的取食和搬运. 应用生态学报, 2013, 24(8): 2325- 2332.
[11] 仝磊, 路纪琪. 黄胸鼠对假海桐和截头石栎种子的贮藏和取食. 兽类学报, 2010, 30(3): 270- 277.
[12] 马庆亮, 赵雪峰, 孙明洋, 路纪琪, 孔茂才. 啮齿动物作用下退耕地山杏种子扩散与贮藏的季节变化. 应用生态学报, 2010, 21(5): 1238- 1243.
[13] 常罡, 邰发道. 季节变化对锐齿栎种子扩散的影响. 生态学杂志, 2011, 30(1): 189- 192.
[14] 张博, 石子俊, 陈晓宁, 廉振民, 常罡. 昆虫蛀蚀对鼠类介导下的锐齿槲栎种子扩散的影响. 生态学报, 2014, 34(14): 3937- 3943.
[15] Yu F, Shi X X, Wang D X, Yi X F, Fan D F, Guo T D, Luo Y G. Effects of insect infestation onQuercusalienavar.acuteserrataacorn dispersal in the Qinling Mountains, China. New Forests, 2015, 46(1): 51- 61.
[16] 路纪琪, 张知彬. 灌丛高度对啮齿动物贮藏和扩散辽东栎坚果的影响. 动物学报, 2005, 51(2): 195- 204.
[17] 王真真. 山杏种子挥发性化学物质对鼠类种子贮藏行为的影响[D]. 武汉: 华中师范大学, 2015.
[18] Wauters L A, Githiru M, Bertolino S, Molinari A, Tosi G, Lens L. Demography of alpine red squirrel populations in relation to fluctuations in seed crop size. Ecography, 2008, 31(1): 104- 114.
[19] Theimer T C. Seed scatterhoarding by white-tailed rats: consequences for seedling recruitment by an Australian rain forest tree. Journal of Tropical Ecology, 2001, 17(2): 177- 189.
[20] Yu F, Shi X X, Wang D X, Wang T, Yi X F, Lou Y G. Seed predation patterns favor the regeneration of dominant species in forest gaps compared with the understory in an oak-pine mixed forest. Acta Theriologica, 2014, 59(4): 495- 502.
[21] 彭舜磊, 王得祥. 秦岭火地塘林区华山松人工林与天然次生林群落特征比较. 西北植物学报, 2009, 29(11): 2301- 2311.
[22] 张知彬, 王福生. 鼠类对山杏(Prunusarmeniaca)种子扩散及存活作用研究. 生态学报, 2001, 21(5): 839- 845.
[23] 于飞, 牛可坤, 焦广强, 吕浩秋, 易现峰. 小型啮齿动物对小兴安岭5种林木种子扩散的影响. 东北林业大学学报, 2011, 39(1): 11- 13.
[24] Smallwood P D, Steele M A, Faeth S H. The ultimate basis of the caching preferences of rodents, and the oak-dispersal syndrome: tannins, insects, and seed germination. Integrative & Comparative Biology, 2001, 41(4): 840- 851.
[25] 苗迎权, 闫兴富, 杜茜, 周云锋, 孙毅. 啮齿动物对模拟昆虫侵染种子的取食和扩散. 福建林业科技, 2016, 43(1): 46- 52, 88.
[26] 闫兴富, 余杨春, 周立彪, 周云峰. 啮齿动物对六盘山区辽东栎、野李和华山松种子的取食和搬运. 生物多样性, 2012, 20(4): 427- 436.
[27] 张洪茂. 北京东灵山地区啮齿动物与森林种子间相互关系研究[D]. 北京: 中国科学院研究生院, 2007.
[28] 肖治术, 张知彬, 王玉山. 小泡巨鼠对森林种子选择和贮藏的观察. 兽类学报, 2003, 23(3): 208- 213.
[29] Jenkins S H, Peters R A. Spatial patterns of food storage by Merriam′s kangaroo rats. Behavioral Ecology, 1992, 3(1): 60- 65.
[30] Dally J M, Clayton N S, Emery N J. The behaviour and evolution of cache protection and pilferage. Animal Behaviour, 2006, 72(1): 13- 23.
[31] Preston S D, Jacobs L F. Conspecific pilferage but not presence affects Merriam′s kangaroo rat cache strategy. Behavioral Ecology, 2001, 12(5): 517- 523.
[32] Jones F A, Peterson C J, Haines B L. Seed predation in neotropical pre-montane pastures: site, distance, and species effects. Biotropica, 2003, 35(2): 219- 225.
[33] 杨晓兰, 巩合德. 滇西北小型啮齿动物对不同植物种子的捕食差异调查. 浙江农林大学学报, 2015, 32(3): 440- 445.
[34] Wilkinson E B, Branch L C, Miller D L. Functional habitat connectivity for beach mice depends on perceived predation risk. Landscape Ecology, 2013, 28(3): 547- 558.
[35] 兰国玉, 雷瑞德, 安锋, 赵永华, 陈伟. 秦岭华山松种群格局规模与林窗特征. 生态学杂志, 2006, 25(6): 652- 656.
[36] Zhang Z B. Effect of burial and environmental factors on seedling recruitment ofQuercusliaotungensisKoidz. Acta Ecologica Sinica, 2001, 21(3): 374- 384.
Selectionmechanismofrodentsontheremovalandpredationmicrohabitatsofseedsfromdifferenttreespecies
KANG Haibin1,2, WANG Dexiang1,2,*, CHANG Mingjie1, KANG Bing3, HU Youning1,4, YU Fei1,5, PANG Yue1, TENG Qinglin1
1CollegeofForestry,NorthwestAgricultureandForestryUniversity,Yangling712100,China2NationalPositioningObservationStationofForestEcosystematXiaolongshaninGansu,Tianshui741020,China3CollegeofLifeSciences,NorthwestAgricultureandForestryUniversity,Yangling712100,China4CollegeofBiologicalandEnvironmentalEngineering,Xi′anUniversity,Xi′an710065,China5CollegeofLifeSciences,HenanNormalUniversity,Xinxiang453007,China
Different types of seeds could be dispersed in various ways. Rodents show a significant preference in the microhabitats of predation sites and caching methods after seeds removal. Understanding the relationship between storing behavior and microhabitat is the key to study seedling establishment. The objective of this study was to reveal the effects of small rodents on seed dispersal patterns of constructive species in pine-oak mixed forests. Field experiments were conducted in the Huoditang forest region in the middle of the Qinling Mountains, Shaanxi Province, northwestern China. Coded plastic tags were used to study the seed fates ofQuercusalienavar.acuteserrata,Pinustabuliformis, andPinusarmandii. The results showed that theinsituseed predation rate ofP.tabuliformisseeds was significantly higher than the seeds of the other three types of seeds.P.armandiiseeds had the highest predation rate (60%) and cache rate (4.33%) after removal by rodents, and the longest average dispersal distance of predation sites (2.49 m). The small acorns ofQ.alienavar.acuteserratahad the longest dispersal distance of cache sites after removal by rodents (4.92 m). All types of seeds exceptP.armandiiseeds had over 85% of the sites were used as predation sites.P.tabuliformisseeds had no cache sites. Over 90% of the cache sites of the other three types of seeds contained a single seed. Most of the seeds were discarded after removal by rodents. The proportions of seeds that were cached under shrubs were higher for large and small acorns ofQ.alienavar.acuteserrataandP.armandiiseeds, which were 87.5%, 78.57% and 53.33% of the total number of seeds after cache by rodents, respectively. A lower proportion of seeds was cached on bare land. Most of seeds were eaten under shrubs. OnlyP.armandiiseeds were eaten in the holes after removal by rodents. A higher proportion ofP.tabuliformisseeds were eateninsituthan the other three types of seeds. ExceptP.tabuliformisseeds, the predation rates for the other three types of seeds tended to decrease along microhabitat changing from simple to complex (shrub to grass to shrub edge to bare land). The nutritive value of the seeds and the cost of the rodents′ predation and removal process were the key factors affecting the fates of seeds, and rodents showed obvious preference in the microhabitats of seed storage and predation sites.
small rodent; seed dispersal; foraging strategy; microhabitat; pine-oak mixed forest; Qinling Mountains
国家自然科学基金项目 (31470644);河南省高等学校重点项目 (15A180047)
2016- 09- 17; < class="emphasis_bold">网络出版日期
日期:2017- 07- 12
*通讯作者Corresponding author.E-mail: wangdx66@126.com
10.5846/stxb201609171871
康海斌,王得祥,常明捷,康冰,胡有宁,于飞,庞越,滕青林.啮齿动物对不同林木种子的搬运和取食微生境选择机制.生态学报,2017,37(22):7604- 7613.
Kang H B, Wang D X, Chang M J, Kang B, Hu Y N, Yu F, Pang Y, Teng Q L.Selection mechanism of rodents on the removal and predation microhabitats of seeds from different tree species.Acta Ecologica Sinica,2017,37(22):7604- 7613.
