BCI 50 hm2森林大样地2010年活立木空间结构分析
2018-01-03周红敏王宏翔赵中华惠刚盈
彭 辉 ,周红敏 ,王宏翔 ,赵中华 ,惠刚盈*
(1.中国林业科学研究院林业研究所,国家林业局林木培育重点实验室,北京 100091;2.浙江凤阳山自然保护区管理处,浙江 龙泉 323700; 3.龙泉市林业科学研究院,浙江 龙泉 323700)
BCI50hm2森林大样地2010年活立木空间结构分析
彭 辉1,2,周红敏3,王宏翔1,赵中华1,惠刚盈1*
(1.中国林业科学研究院林业研究所,国家林业局林木培育重点实验室,北京 100091;2.浙江凤阳山自然保护区管理处,浙江 龙泉 323700; 3.龙泉市林业科学研究院,浙江 龙泉 323700)

BCI样地;空间结构;二元分布
森林结构是森林动态变化过程中测度时点的林分状态的高度概括和度量。随着森林可持续经营对森林精确信息的需求增加,森林空间结构研究越显重要[1-3]。林分空间结构决定林木间的竞争态势及其空间生态位,在很大程度上决定林分的稳定性、发展的可能性和经营空间的大小[4-7]。目前,在森林结构解析中,基于相邻木空间关系的林分空间结构分析方法已被广泛应用,由于其可释性、简洁性和可操作性等优点,已经广泛应用在森林类型的研究中,为结构化经营提供了科学依据[8]。森林固定监测大样地在前所未有的尺度上为研究种群动态等生态学规律和现象提供了良好的契机[9],是当前生态学研究中最活跃的领域之一[10-13]。基于最近相邻木关系的林分空间结构参数角尺度、混交度和大小比数等在国内外关于林分空间结构分析、林木竞争与优势度计算、物种多样性测度以及结构恢复重建与优化调整等研究目前主要是以1 hm2或更小面积的样地作为研究对象,在全球的森林大样地中应用较少。各国学者对位于巴拿马的BCI 50 hm2的大样地进行了大量的研究,在生物多样性的维持机制、物种的点格局和群落动态等方面产生了巨大的影响[14-17]。但对BCI 50 hm2大样地空间结构一元及二元分布特征的分析却鲜有报道。因此,作者以BCI 50 hm2大样地为研究对象,利用空间结构参数一元分布和二元分布,量化评价BCI 50 hm2大样地(2010年第7次普查)的活立木空间结构特征,探讨空间结构参数在大样地中的应用前景以及对空间结构参数理论体系进行进一步验证。
1 材料
分析采用BCI样地第7次(2010年)的普查数据资料。数据来源网络以R.data形式保存的公开数据(http://dx.doi.org/10.5479/data.bci.20130603),数据的详细信息及全部物种名录详见:https://repository.si.edu/handle/10088/20925。

2 研究方法
BCI样地数据存储的格式为 Plotname.full#.rdata,#代表调查的次数,如第7次调查(2010年)文件的名称为bci.full7.rdata。每次调查的数据框(dataframe)包含着相同的行数(number of rows), 其中“treeID”行包含了所有调查过的树木的编号,无论其补员(Recruitment)、存活(Alive)或者死亡(Dead)都会永久保留其标识号,因为行数上的匹配,历次的调查数据可以方便的进行比较分析。本研究以样地中DBH≥1 cm的活立木(Alive)为分析对象,并剔除数据中的缺失值(NA)。BCI样地第7次调查中活立木有297个物种,共201 434株林木。其中冠层包含180个物种7 410株林木,占总物种数的60%,占总株数的3.68%;下层有290个物种194 024株林木,占总物种数的97.6%,占总株数的96.3%。
在热带森林中,一般以胸径(DBH)超过一定标准作为冠层的划分,例如Hubbell和Fosetr[21]以20 cm划分冠层,Steege等以30 cm作为冠层[24],La Frankie等以40 cm作为冠层[25]等,本文将BCI样地第7次调查数据中所有活立木个体分为冠层林木(DBH≥20 cm)和下层林木(1 cm≤DBH<20 cm)进行空间结构分析。
采用R 3.3.2软件分析,Excel 2013作图,分析BCI样地整体林分以及冠层和下层活立木的一元和二元分布空间特征。空间结构二元分布是基于最近 4 株相邻木的空间关系构建的林分空间结构分析方法,可从两个方面同时解析林分空间结构的微观信息,结合空间结构一元分布对林分宏观特征的解析,其理论基础是基于有直观明了的空间解析能力以及彼此独立且有相同的结构划分的角尺度 (W) ,混交度(Mingling,M) 和大小比数(U) 3个空间结构参数,其计算方法详见文献[26-27],为避免边缘效应对林分结构的影响,计算时缓冲区设置为5 m。
3 结果与分析
3.1 BCI大样地空间结构一元分布特征
3.2 BCI大样地空间结构的二元分布特征

图1 BCI样地第7次(2010年)普查空间结构一元分布Fig.1 The univariate distribution of spatial structure in BCI plot
3.2.3 BCI林分的混交度和角尺度的二元分布 在W-M的组合中(图2C),混交度的整体分布趋势随着角尺度的增大表现为先增大后减小,均为正态分布。在随机分布轴(Wi=0.50)上,混交度频率值之和最大,为57.4%,在均匀分布状态(Wi=0.25)时,频率值为21.2%,两者的频率值总和为78.6%。在很均匀分布(Wi=0),林木株数最少,所占频率值为0.7%,其次为聚集(很不均匀)分布状态(Wi=1),频率值为3.8%。在混交度(Mi=0.00)和角尺度(Wi=0.00)的组合上频率值为0.000 02,表明只有5株林木在零度混交时均匀分布。最大频率值出现在混交度(Mi=1.00)和角尺度(Wi=0.50)的结构组合上,频率值为42.9%,即样地中大部分林木个体处于随机分布且极强度混交状态,这也从参照树同种的最近邻体(Mi=0.00→0.50)的分布频率不足8%得到证明。
3.3 BCI大样地冠层的空间结构二元分布
以中度混交轴(Mi=0.50)和中庸轴(Ui=0.50)为界(图2D),在零度混交(Mi=0.00)时,林木株数几乎为0,在低混交等级(Mi=0.00→0.25)上,冠层林中优势木(Ui=0.00,Ui=0.25)频率几乎为0。说明冠层林木中参照树周围最近4株相邻木中极少同种的个体存在。而在高混交等级(Mi=0.75→1.00)时,优势木频率最高。在高度混交(Mi=1.00)时,各等级大小比数林木株数均较多,达到78%。在同等级大小比数上,林木株数随着混交度的增大而增大。
由图2E可知,随着大小比数等级的增加角尺度频率值为先增大后减小,大小比数均在Wi=0.5时频率值最大,各等级大小比数之和在Wi=0.5轴上的总和为57.5%,冠层中各等级大小比数的林木株数差异不显著。从图2F可知,在相同的混交度等级上,角尺度的频率值为正态分布,即先增大后减小。在Wi=0.50时,混交度各频率值之和最大为57.5%,说明冠层有1/2以上的林木属于随机分布。而Wi=0.00和Wi=1.00的比例为1%和1.6%,说明BCI样地冠层林木个体中较少处于很均匀和很不均匀分布的状态。
3.4 BCI大样地下层的空间结构二元分布
BCI样地(2010年)下层林木的混交度和大小比数(M-U)的组合中(图2G),零度混交轴(M=0.00)的大小比数的频率值之和接近于0,其余大部分林木处于高度混交状态。混交度和大小比数的最大频率值为(Mi=1.00,Ui=0.00)的组合为16.4%,即下层中处于极强度混交的劣势木的株数较多。图2H为下层林木角尺度和大小比数(W-U)的二元分布,各等级角尺度的林木株数随着大小比数等级的增加而增大,说明下层中处于劣势状态的林木较多。各等级大小比数的频率值与林分整体和冠层的趋势相似,也表现为正态分布。而且在随机分布状态时,频率值之和最大,为57.4%,处于很均匀分布和聚集分布的林木较少,分别为0.7%和3.8%。下层的林木个体处于零度混交的株数极少(图2I),处于弱度混交(Mi=0.25)的林木个体株数仅占下层总株数的1.8%。各等级混交度的频率值随着角尺度等级的增大表现为先增加后减小,在随机分布状态(Wi=0.50)时,混交度频率值之和最大为57.4%。混交度和角尺度的最大频率值出现在(Mi=1.00,Wi=0.50)的组合上为42.1%,为其它24个组合频率均值的17.5倍。

图2 BCI样地第7次(2010年)普查空间结构二元分布Fig.2 The bivariate distribution of spatial structure in BCI plot
4 讨论
(1)在自然条件下,种群的结构和空间分布往往是种群生物学特性、种内种间关系以及环境条件长期相互适应、相互作用的结果[14]。采用基于最近相邻木的空间结构参数对BCI大样地第7次调查(2010年)活立木进行分析,林分整体表现为随机分布。在混交度方面,整体林分表现为大部分林木处于强度和极强度混交状态,而且处于极强度混交的优势木株数较多。样地内大多数林木DBH在5 cm以下,说明林分中幼苗储备充足,样地中处于劣势的林木较多。
(2)以BCI整体林分为例,通过一元分布可知林分中林木个体大多数处于极强度混交,但不知处于极强度混交的林木个体处于哪种分布格局或在林分中处于怎样的态势。通过混交度-大小比数(图2A)可知BCI样地中处于极强度混交状态的林木在大小比数各等级的林木株数相差不大;角尺度-混交度(图2C),可以很直观的展示处于极强度混交状态的林木个体在随机分布状态的频率值为0.429,说明林分中处于极强度混交状态的林木大部分处于随机分布状态。
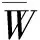
5 结论
(1)BCI 样地(2010年)整体林分为随机分布格局,处于很均匀分布的林木较少。林分处于极强度混交状态,处于中低度混交等级的林木较少,在零度、弱度和中度3个等级的频率之和不足8%。处于零度混交状态且处于均匀分布的林木最少(频率值为0.000 02),大部分林木处于随机分布状态且极强度混交状态。林木株数在各大小比数等级上的频率差别不大,处于极强度混交和绝对优势状态的林木较多,占总株数的17%。处于绝对劣势状态且随机分布的林木株树最多。
(2)BCI 样地(2010年)冠层林木大多数处于高度混交状态,在零度混交状态的林木极少。在同一大小比数等级上,不同混交状态的林木株数存在显著差异。冠层林木大多数处于随机分布状态,在团状聚集和很均匀分布状态的林木较少。冠层林木在各等级优劣程度上的株树相差不大,大致为1∶1∶1∶1。
(3)BCI 样地(2010年)下层林木分布格局为随机分布,大部分林木处于高度混交状态,处于弱度混交状态的林木株树也较少,处于极强度混交状态且处于随机分布状态的林木株数是其它混交度-角尺度组合的17.5倍。下层林木中处于劣势状态的林木株数相对较多。
综上所述, BCI 样地(2010年)林分整体、冠层林木、下层林木分布格局均表现为随机分布。林分整体、冠层林木和下层林木大部分林木均处于极强度混交状态,处于绝对劣势的林木株数最多,处于劣势的林木株数相对较少;整体表现为中庸偏劣势的状态。
[1] 赵中华,惠刚盈,胡艳波,等. 基于大小比数的林分空间优势度表达方法及其应用[J]. 北京林业大学学报, 2014, 36(1):78-82.
[2] Pommerening A. Approaches to quantifying forest structures[J]. Foresty, 2002, 75(3):305-324.
[3] 周红敏,彭 辉,柳新红,等.萌生杉木林经营后林分空间结构变化[J]. 浙江林业科技, 2015, 35(4):41-46.
[4] 巫志龙,周成军,周新年,等. 杉阔混交人工林林分空间结构分析[J]. 林业科学研究, 2013, 26(5):609-615.
[5] 马 琳,梁 军,周国英,等. 昆嵛山天然赤松次生林空间结构特征的研究[J]. 中南林业科技大学学报, 2013, 33(9):50-54.
[6] 周红敏,惠刚盈,赵中华,等. 森林结构调查中最适样方面积和数量的研究[J]. 林业科学研究, 2009, 22(4):482-495.
[7] 惠刚盈,李 丽,赵中华. 林木空间分布格局分析方法[J]. 生态学报, 2007, 27(11):4717-4728.
[8] 周红敏,何必庭,彭 辉,等.萌生杉木林空间结构特征研究[J]. 林业科学研究, 2015, 28(5):686-690.
[9] Chisholm R A, Condit R, Rahman K A,etal. Temporal variability of forest communities: empirical estimates of population change in 4000 tree species[J]. Ecology Letters, 2014, 17(7):855-65.
[10] Hubbell S P, Ahumada J A, Condit R,etal. Local neighborhood effects on long-term survival of individual trees in a Neotropical forest[J]. Ecological Research, 2001, 16(5):859-875.
[11] Wang X G, Wiegand T, Wolf A,etal. Spatial patterns of tree species richness in two temperate forests[J]. Journal of Ecology, 2011, 99(6):1382-1393.
[12] 马克平.生物多样性与生态系统功能的实验研究[J]. 生物多样性, 2013, 21(3): 247-248.
[13] 王慧杰,常顺利,张毓涛,等. 天山雪岭云杉森林群落的密度制约效应[J]. 生物多样性, 2016, 24(3):252-261.
[14] Condit R, Ashton P S,Baker P,etal.Spatial patterns in the distribution of tropical species[J]. Science, 2000, 288(5470):1414-1418.
[15] Condit R, Ashton P, Bunyavejchewin S,etal. The importance of demographic niches to tree diversity[J]. Science, 2006, 313(5783):98.
[16] Nathan R. Long-distance dispersal of plants[J]. Science, 2006, 313(5788):786-788.
[17] 郝占庆,李步杭,张 健,等.长白山阔叶红松林样地(CBS):群落组成与结构[J]. 植物生态学报, 2008, 32 (2):238-250.
[18] Bohlman S A. Species Diversity of Canopy versus Understory Trees in a Neotropical Forest: Implications for Forest Structure, Function and Monitoring[J]. Ecosystems, 2015, 18(4):658-670.
[19] Croat T R. Flora of Barro Colorado Island[M]. Stanford, CA: Stanford University Press, 1978.
[20] Leigh E G, Jr Rand S A, Windsor D M. The Ecology of a Tropical Forest: Seasonal Rhythms and Long- Term Changes[M]. Washington, DC: Smithsonian Institution Press, 1982.
[21] Hubbell S P, Foster R B. Diversity of canopy trees in a neotropical forest and implications for conservation[M]// Sutton S L, Whitmore T C, Chadwick A C. Tropical Rain Forest: Ecology and Management. Oxford: Blackwell Scientific Publications, 1983: 25-41.
[22] Condit R. Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and Comparison with Other Plots[M]. Berlin: Springer, 1998.
[23] Condit R, Chisholm R A, Hubbell S P. Thirty years of forest census at Barro Colorado and the importance of immigration in maintaining diversity[J]. PLoS ONE, 2012, 7(11):1-6.
[24] Steege H, Pitman N C A, Sabatier D,etal. Hyperdominance in the Amazonian tree flora[J]. Science, 2013, 342(6156):1243092.
[25] LaFrankie J V, Ashton P S, Chuyong G B,etal. Contrasting structure and composition of the understory in species-rich tropical rain forests[J]. Ecology, 2006, 87(9):2298-305.
[26] LI Y F,HUI G Y,ZHAO Z H,etal.The bivariate distribution characteristics of spatial structure in natural Korean pine broadleaved forest[J].Journal of Vegetation Science, 2012, 23(6):1180-1190.
[27] 李远发. 林分空间结构参数二元分布的研究[D].北京: 中国林业科学研究院, 2013.
[28] Wiegand T, Moloney K A.Rings, circles and null-models for point pattern analysis in ecology[J]. Oikos, 2004, 104(2):209-229.
[29] Wright S J. Plant diversity in tropical forests: a review of mechanisms of species coexistence[J]. Oecologia, 2002, 130(1):1-14.
[30] Barot S, Gignoux J, Menaut J C. Demography of a savanna palm tree: predictions from comprehensive spatial pattern analyses[J]. Ecology, 1999, 80(6):1987-2005.
ResearchonSpatialStructureof50hm2ForestPlotatBarroColoradoIslandin2010
PENGHui1,2,ZHOUHong-min3,WANGHong-xiang1,ZHAOZhong-hua1,HUIGang-ying1
(1.Key Laboratory of Tree Breeding and Cultivation, State Forestry Administration; Research Institute of Forestry,Chinese Academy of Forestry, Beijing 100091, China; 2.Fengyangshan National Nature Reserve, Longquan 323700, Zhejiang, China; 3.Longquan Forestry Research Institute, Longquan 323700, Zhejiang, China)

BCI plot;spatial structure;bivariate distribution
10.13275/j.cnki.lykxyj.2017.06.002
2017-03-20
国家自然科学基金项目(31370638)
彭 辉,工程师。
* 通讯作者:惠刚盈,研究员。E-mail:hui@caf.ac.cn
S718.54
A
1001-1498(2017)06-0895-07
彭南轩)
