黄条不同组织碳稳定同位素的转化率与分馏
2018-01-02韩羽嘉田甲申李多慧王震鹿志创木云雷
韩羽嘉 ,田甲申 ,李多慧 ,王震 ,鹿志创 ,木云雷
(1.大连海洋大学,辽宁 大连 116023;2.辽宁省海洋水产科学研究院,辽宁省海洋生物资源和生态学重点实验室,辽宁 大连 116023;3.大连市水产研究所,辽宁 大连 116019)
韩羽嘉1,2,田甲申2,李多慧3,王震1,2,鹿志创2,木云雷2
(1.大连海洋大学,辽宁 大连 116023;2.辽宁省海洋水产科学研究院,辽宁省海洋生物资源和生态学重点实验室,辽宁 大连 116023;3.大连市水产研究所,辽宁 大连 116019)
在水温18.79~22.15℃的实验室条件下,给体质量(34.2±1.8)g的黄条Seriola aureovittata投喂已知碳源的冰鲜玉筋鱼Ammodytes personatus肌肉,研究黄条不同组织碳稳定同位素的转化率差异及分馏系数,以探讨组织生长和代谢对不同组织碳稳定同位素转化的相对贡献。结果表明,黄条各组织碳稳定同位素半衰期从小到大依次为肝脏(3.2d)、肠(3.4d)、鳃(3.6d)、心脏(5.5d)、肌肉(6.4d),各组织与饵料间的碳稳定同位素富集系数(Δδ13C)从小到大依次为肝(0.15‰)、肠(0.48‰)、心脏(0.98‰)、鳃(1.02‰)、肌肉(1.08‰)。代谢作用对肝脏和肠组织的碳稳定同位素转化率的贡献占主导地位(>60%),生长作用对其贡献相对较小。肝脏和肠组织可反映黄条短期内的摄食特征,而肌肉组织则可反映较长时间的摄食特征。本研究结果可丰富黄条基础生物学内容,为不同时间尺度的稳定同位素研究取样和工厂化养殖中的营养强化、饵料选择等提供参考。
黄条;碳稳定同位素;同位素分馏;转化率
近年来,稳定同位素技术已广泛应用于海洋生态系统研究,成为海洋生物摄食特征、食物网结构特征、生物间的营养关系等问题研究的重要分析手段之一[1]。稳定同位素分析方法虽然能够反映生物较长期的摄食特征,但当生物体摄食发生变化时,其稳定同位素特征反映的摄食变化存在着时间上的滞后[2]。监测生物稳定同位素比率在食性转变后重新达到分馏平衡所需的时间,将同位素比率随时间的变化拟合成函数探究稳定同位素转化规律,可以提高稳定同位素分析方法在研究生物摄食中的精确度[3,4]。稳定同位素转化由新组织的生长和旧组织的降解代谢共同决定,稳定同位素转化率不仅具有物种特质,还受个体生长阶段、食物营养结构、环境温度等多种因素的影响,甚至同一生物个体的不同组织间也存在差异。Witting等[5]研究美洲拟鲽Pseudopleuronectes americanus稳定同位素转化率发现,水温较高时肌肉组织半衰期较短,认为代谢活动与同位素转化率呈正相关。Logan等[6]研究发现,与肌肉组织和血液细胞相比,内部器官和血浆具有较高的稳定同位素转化率。组织在不同的时间尺度整合食物,对多个组织的检查可以潜在地提供关于资源使用的时间动态信息[7]。一般来说,肝组织代谢速率高于肌肉和心脏,仔鱼的半衰期不到一个星期,而大鱼可能要几个月[5-7]。在应用稳定同位素方法分析生物体摄食习性时,要根据所研究的时间选择适合的组织。
目前在实验室条件下对鱼类不同组织碳稳定同位素转化率和分馏系数(Δδ13C)的研究较多。如Guelinckx等[8]研究发现,刺鳍鱼 Pomatoschistus minutus肌肉组织的转化率较低,半衰期为25d,而肝脏的半衰期仅为3d,肌肉和肝脏的Δδ13C分别为0.21‰和0.77‰。Buchheister和Latour[9]发现犬齿牙鲆Paralichthys dentatus肌肉和肝脏的半衰期分别为49d和10d,Δδ13C分别为0.23‰和0.91‰。张妙等[10]研究发现,黄颡鱼Pelteobagrus fulvidraco肝脏和鳃的半衰期为 26d和 197d,Δδ13C为 -2.1‰和0.3‰。但关于黄条碳稳定同位素转化率与分馏效应的研究尚未见报道。
1 材料与方法
1.1 材料
1.2 方法
1.2.1 实验设计和日常管理
1.2.2 样品处理
1.2.3 稳定同位素测定
所有样品在辽宁省海洋水产科学研究院稳定同位素实验室,用稳定同位素质谱仪为菲尼根Flash 2000 HT型元素分析仪和菲尼根Delta V Advantage同位素比率质谱仪相连进行测定。稳定C同位素的自然丰度表示为:

式中,R代表13C/12C。δ13C是相对于PDB(Pee Dee Belemnite)标准的自然丰度[14]。每个样品测定3个平行样,每测定5个样品后插测1个标准样。δ13C精密度<±0.15‰。
1.2.4 相关指标的计算
相对增长率WR=ln(Wt/Wi)/t×100%
特定生长率SGR=(lnWt-lnWi)/t×100%/d
式中,Wi、Wt分别为初始和t时的体质量(湿质量,g);t为实验时间。

式中,δt代表t时各组织的同位素比值;δf代表各组织同饵料达到平衡的最终同位素比值;δi代表食性转变前初始同位素比值;转化常数为生长系数(k)和新陈代谢转化常数(m)之和。
生长常数:Wt=Wiekt
半衰期:T0.5=-ln(0.5)/(k+m)
由k和m与(k+m)的比值计算生长和代谢对鱼类组织碳稳定同位素转化的贡献率,通常用Pg和Pm表示[15]。

1.2.5 数据处理
实验数据以平均值±标准差(X±SD)表示。采用SPSS 20.0软件处理和分析数据,用OriginPro 8.0软件曲线拟合和制图。
2 结果与分析
2.1 黄条的生长
2.2 饵料鱼的碳稳定同位素值
饵料鱼的碳稳定同位素比值(δ13C)变化范围为-20.88‰~-20.50‰,跨度为0.38‰,经t检验发现,饵料鱼的 δ13C 无显著性差异(P>0.05)(表 1)。
2.3 黄条不同组织碳稳定同位素营养分馏

图1 黄条的生长Fig.1 The growth of yellow tail Seriola aureovittata
2.4 黄条不同组织碳稳定同位素转化率及半衰期
2.5 生长和代谢对黄条不同组织碳稳定同位素转化率的影响

表1 饵料鱼的δ13CTab.1 The δ13C value of the fish diets
表2 黄条不同组织的δ13CTab.2 The δ13C values of different tissues in yellow tail Seriola aureovittata
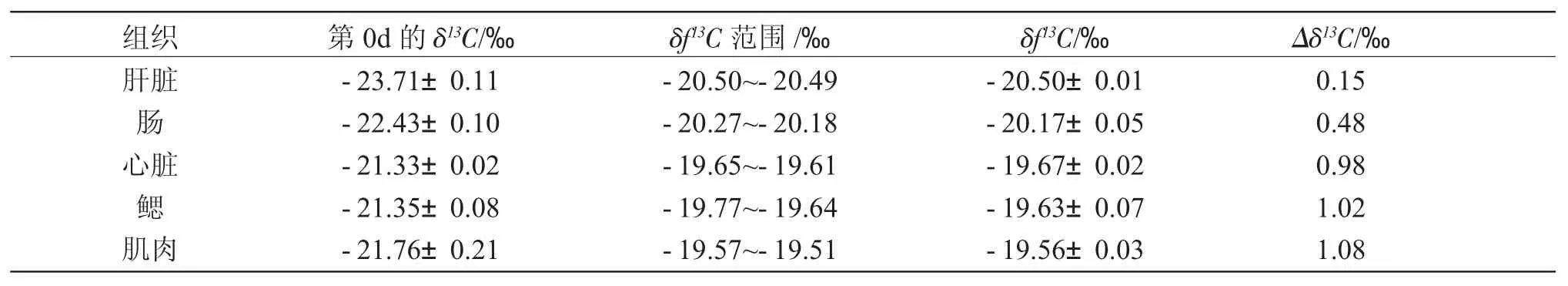
表2 黄条不同组织的δ13CTab.2 The δ13C values of different tissues in yellow tail Seriola aureovittata
组织肝脏肠心脏鳃肌肉第 0d 的 δ13C/‰-23.71±0.11-22.43±0.10-21.33±0.02-21.35±0.08-21.76±0.21 δf13C 范围 /‰-20.50~-20.49-20.27~-20.18-19.65~-19.61-19.77~-19.64-19.57~-19.51 δf13C/‰-20.50±0.01-20.17±0.05-19.67±0.02-19.63±0.07-19.56±0.03 Δδ13C/‰0.15 0.48 0.98 1.02 1.08
3 讨论
3.1 黄条不同组织碳稳定同位素的转化率
生物体的碳稳定同位素值是由其所摄食的食物所决定,当摄食改变时,生物体组织中稳定同位素值取决于稳定同位素在其体组织中的转化速率[7,16,17]。大量的研究结果表明[17-19],摄食新食物后,生物的体质量要增长2.5~4倍才能与其食物达到同位素平衡。如果生物体没有与食物达到同位素平衡,稳定同位素分馏分析就可能产生偏差[20]。本研究中,实验30d后黄条体质量增长3倍,第30d与第35d、第40d肌肉的 δ13C无显著差异(P>0.05),认为黄条各组织已与其饵料达到稳定同位素分馏平衡,不同组织的转化率由大到小依次为:肝、肠、鳃、心脏、肌肉,其中肝脏的转化率大约是肌肉的2倍,与Witting[5]等对犬齿牙鲆食性转变实验的结果一致。生物体各组织与食物达到稳定同位素平衡的速率与其蛋白质周转速率有关,这可能是本研究中肝脏同位素转化率最快的主要原因。Tieszen等[1]在对沙鼠Meriones unguienlatus的摄食生态研究中发现,脂质含量高的组织同位素转化速率高。Church等[21]对虹鳟Oncorhynchus mykiss不同组织稳定同位素转化率分析后发现,粘液具有很高的免疫活性和再生能力,同样有较高的同位素转化率。肠和鳃丝都是黏膜结构,脂质含量高且能够分泌黏液,这可能是引起本研究中黄条肠、鳃组织转化率高的原因之一。

图2 黄条不同组织的δ13C随时间的变化Fig.2 The changes in δ13C value with time in different tissues of yellow tail Seriola aureovittata
表3 黄条不同组织基于时间模型的碳稳定同位素转化参数Tab.3 The transformation parameters of carbon stable isotope in different tissues of yellow tail Seriola aureovittata based on time model

表3 黄条不同组织基于时间模型的碳稳定同位素转化参数Tab.3 The transformation parameters of carbon stable isotope in different tissues of yellow tail Seriola aureovittata based on time model
组织心脏鳃肠肌肉肝脏转化率m+k/d 0.1249 0.1928 0.2016 0.1090 0.2197代谢常数m/d 0.0372 0.1051 0.1139 0.0213 0.1320半衰期T0.5/d 5.5 3.6 3.4 6.4 3.2转化95%时间T0.95/d 24.0 15.5 14.9 27.5 13.6代谢贡献比例Pm/%19.2 54.5 57.1 14.0 61.9生长贡献比例Pg/%80.8 45.5 42.9 86.0 38.1
众多研究表明[22-25],稳定同位素在生物体内会发生差异分配,即生物体所摄取的不同食物成分会被其特定组织或部位选择性吸收;生物不同组织对食物变化的响应速度也不同,组织达到同位素分馏平衡所需的时间差异较大,这可能导致动物特定组织无法反映其整体的同位素组成和变化规律,因此在采用稳定同位素方法开展动物食性与营养相关研究时,应根据特定研究目的和时间选取具有代表性的体组织。
3.2 生长与代谢对黄条不同组织碳稳定同位素转化的影响
稳定同位素转化由两个不同的过程组成,即组织生长和新陈代谢过程[3]。生长过程中,新合成组织的稳定同位素组成会稀释原有组织的稳定同位素组成,而代谢过程则产生组织替换作用[26]。Nathan等[27]认为,生长作用对于生长速率较快的生物同位素转化的影响占主导地位,而代谢作用可能对生长缓慢的生物影响更明显。但Suzuki等[22]对此提出异议,认为生物体不同组织的氨基酸、脂肪酸等营养成分组成及生理代谢速率不同,导致代谢作用对不同组织同位素转化率的影响不同。本研究中,新组织生长对黄条心脏和肌肉组织的转化率贡献占主导作用,而代谢作用对肝脏、肠和鳃组织同位素转化率贡献比例较生长作用更大,这可能与肝脏、肠、鳃组织的代谢速率较高有关,与Madigan等[28]对太平洋蓝鳍金枪鱼Thunnus orientalis同位素转化率研究结果一致。
3.3 黄条不同组织碳稳定同位素的富集系数
碳同位素富集系数即动物组织与其食物之间同位素丰度(δ13C)的差异值(Δδ13C),是应用稳定性同位素技术分析生态系统营养关系、食性和食物网时的重要参数[29]。Peterson等[30]比较大量研究文献后发现,消费者的碳稳定同位素和食物来源基本相似,富集系数总体平均值在0‰~1‰,同一营养级的碳稳定同位素富集为1.5‰左右,经常用于分析食物来源及物质与能量的流动。Rio等[31]指出,在食性转变实验中,选用的饵料应尽量满足研究生物的营养需求,若饵料缺乏该种生物在特定生长阶段必需的营养物质,生物体就会提高摄食量来弥补食物中缺乏营养物质,这使得生物体必须进行更多非必须营养物质的代谢循环,导致更多重稳定同位素在生物体内大量积累,使生物与食物间分馏系数过高。Trueman等[32]通过投喂人工饲料研究发现,大西洋鲑Salmo salar肌肉的Δδ13C为2.1‰、肝脏为1.6‰。周晖等[33]以啤酒酵母和玉米蛋白为饵料,发现军曹鱼 Rachycentron canadum肌肉的 Δδ13C为 4.04‰。本研究中黄条不同组织的Δδ13C差异可能是由于脂肪含量不同引起,生物体在合成脂肪的过程中,发生明显的碳歧视效应,即趋向于利用较轻的12C合成脂肪,脂肪中的δ13C要比蛋白质和单糖类化合物低6‰[34]。肝脏的脂肪含量高于其他组织,可能就是本研究中肝脏的分馏系数小于其他组织的原因。黄条不同组织的Δδ13C为0.15‰~1.08‰,不同组织的富集系数从小到大依次为:肝、肠、心脏、鳃、肌肉,这与黄条在新陈代谢过程中体内物质与能量的流动途径基本一致,即从肝、肠等消化器官通过血液流入心脏,再通过循环系统进入鳃完成气体交换最后到达肌肉。蔡德陵等[35]对鲈Lateolabrax japonicus的稳定同位素分馏作用研究中也发现相似的规律。本研究认为,结合生物体不同组织的稳定同位素转化率和不同组织的稳定同位素富集系数这两项参数共同分析,可能会量化展示出生物体内营养物质和能量的流动途径。
[1]Tieszen LL,Boutton TW,Tesdahl K G,et al.Fractionation and turnover of stable carbon isotopes in animal tissues:implications for δ13C analysis of diet[J].Oecologia,1983,57(1-2):32-37.
[2]Fry B and Arnold C.Rapid13C/12C turnover during growth of brown shrimp (Penaeus aztecus)[J].Oecologia,1982,54(2):200-204.
[3]Macavoy S E,Macko S A and Garman G C.Isotopic turnover in aquatic predators:quantifying the exploitation of migratory prey[J].Canadian Journal of Fisheries&Aquatic Sciences,2001,58(5):923-932.
[4]Mcintyre P B and Flecker A S.Rapid turnover of tissue ni-trogen of primary consumers in tropical freshwaters[J].Oecologia,2006,148(1):12-21.
[5]WittingD A,Chambers R C,Bosley K L,et al.Experimental evaluation of ontogenetic diet transitions in summer flounder (Paralichthys dentatus),using stable isotopes as diet tracers[J].Canadian Journal of Fisheries&Aquatic Sciences,2004,61(11):2069-2084.
[6]Logan J,Haas H,Deegan L,et al.Turnover rates ofnitrogen stable isotopes in the salt marsh mummichog,Fundulus heteroclitus,following a laboratory diet switch[J].Oecologia,2006,147(3):391-395.
[7]Del Rio C M,Wolf N,Carleton S A,et al.Isotopic ecology ten years after a call for more laboratory experiments[J].Biological Reviews of the Cambridge Philosophical Society,2009,84(1):91-111.
[8]Guelinckx J,Maes J,Van D D P,et al.Changes in δ13C and δ15N in different tissues of juvenile sand goby Pomatoschistus minutus:a laboratory diet-switch experiment[J].Marine EcologyProgress,2007,341(1):205-215.
[9]Buchheister Aand Latour R J.Turnover and fractionation of carbon and nitrogen stable isotopes in tissues of a migratory coastal predator,summer flounder(Paralichthys dentatus)[J].Canadian Journal of Fisheries&Aquatic Sciences,2010,67(3):445-461.
[10]张妙,陈新军,陈亚,等.黄颡鱼不同组织碳氮稳定同位素的周转与分馏研究[J].上海海洋大学学报,2016,25(6):822-830.
[11]姜大为,林乐玲,陈勇,等.黄条室内越冬及生长观察[J].大连海洋大学学报,2001,16(3):223-227.
[12]钟建兴,蔡良候,郑惠东,等.黄条胚胎发育观察[J].渔业研究,2010(2):22-25.
[13]Bligh E G and Dyer W J.A rapid method oftotal lipid extraction and purification[J].Canadian JournalofBiochemistryand Physiology,1959,37(8):911-917.
[14]Mariotti A.Atmospheric nitrogen is a reliable standard for natural15N abundance measurements[J].Nature,1983,303(5919):685-687.
[15]Buchheister A B and Latour R J.Turnover and fractionation of carbon and nitrogen stable isotopes in tissues of a migratorycoastal predator,summer flounder(Paralichthys dentatus)[J].Canadian Journal of Fisheries&Aquatic Sciences,2009,67(3):445-461.
[16]Deniro MJ and Epstein S.Influence of diet on the distribution of carbon isotope ratios in animals[J].Geochim CosmochimActa,1978,42(5):495-506.
[17]Deniro MJ and Epstein S.Influence of diet on the distribution of nitrogen isotopes in animals[J].Geochim CosmochimActa,1981,45(3):341-351.
[18]Anderson R K,Parker P L and Lawrence A.A13C/12C Tracer studyofthe utilization ofpresented feed by a commercially important shrimp Penaeus vannamei in a pond growout system[J].Journal ofthe World Aquaculture Society,1987,18(3):148-155.
[19]Vander Zanden MJ,HulshofM,Ridgway M,et al.Application of stable isotope techniques to trophic studies of age-0 smallmouth bass[J].Transactions of the American Fisheries Society,1998,127(5):729-739.
[20]O′Reilly C M,Hecky R E,Cohen A S,et al.Interpreting stable isotopes in food webs:recognizing the role of time averaging at different trophic levels[J].Limnology&O-ceanography,2002,47(1):306-309.
[21]Church MR,Ebersole J L,Rensmeyer K M,et al.Mucus:a new tissue fraction for rapid determination of fish diet switchingusingstable isotope analysis[J].Canadian Journal ofFisheries&Aquatic Sciences,2009,66(1):1-5.
[22]Suzuki K W,Kasai A,Nakayama K,et al.Differential isotopic enrichment and half-life among tissues in Japan[J].Canadian Journal of Fisheries&Aquatic Sciences,2005,62(3):671-678.
[23]SeminoffJ A,Bjorndal K A and Bolten A B.Stable carbon and nitrogen isotope discrimination and turnover in pond sliders Trachemys scripta:insights for trophic study of freshwater turtles[J].Copeia,2001,2007(3):534-542.
[24]Sweeting C J,Barry J,Barnes C,et al.Effects of body size and environment on diet-tissue δ15N fractionation in fishes[J].Journal of Experimental Marine Biology&E-cology,2007,340(1):1-10.
[25]Yan V,Banas D,Thomas M,et al.Stable isotope variability in tissues of the Eurasian perch Perca fluviatilis[J].Comparative Biochemistry&Physiology Part A Molecular&Integrative Physiology,2007,148(3):504-509.
[26]Hesslein R H,Hallard K A and Ramlal P.Replacement of sulfur,carbon,and nitrogen in tissue of growing broad whitefish (Coregonus nasus)in response to a change in diet traced by δ34S,δ13C,and δ15N[J].Canadian Journal of Fisheries&Aquatic Sciences,1993,50(6):1267-1272.
[27]Wolf N,Carleton S A and del Rio C M.Ten years of experimental animal isotopic ecology[J].Functional Ecology,2009,23(1):17-26.
[28]Madigan D J,Litvin S Y,Popp B N,et al.Tissue turnover rates and isotopic trophic discrimination factors in the en-dothermic teleost,Pacific bluefin tuna(Thunnus orientalis)[J].Plos One,2012,7(11):e49220.
[29]Hobson K A and Clark R G.Assessing avian diets using stable isotopes I:turnover of13C in tissues[J].Condor,1992,94(1):181-188.
[30]Peterson B J and Fry B.Stable isotopes in ecosystem studies[J].AnnualReviewofEcology&Systematics,1987,18(1):293-320.
[31]Rio C MD and Wolf B O.Mass-balance models for animal isotopic ecology[M].In:Physiological and Ecological Adaptations toFeedingin Vertebrates(eds Starck,J.M.&Wang,T.).SciencePublishers,2005,Enfield,NewHampshire,pp.141-174.
[32]Trueman C N,Mcgill R A and Guyard P H.The effect of growth rate on tissue-diet isotopic spacing in rapidly growing animals.An experimental study with Atlantic salmon(Salmo salar)[J].Rapid Communications in Mass SpectrometryRcm,2005,19(22):3239-3247.
[33]周晖,陈刚,施钢,等.不同蛋白源对军曹鱼碳、氮稳定同位素分馏的影响[J].热带海洋学报,2014,33(5):35-40.
[34]DeNiro M J and Epstein S.Mechanism of carbon isotope fractionation associated with lipid synthesis[J].Science,1977,197(4300):261-263.
[35]蔡德陵,张淑芳,唐启升,等.鲈鱼新陈代谢过程中的碳氮稳定同位素分馏作用[J].海洋科学进展,2003,21(3):308-317.
Conservation Rate and Fractionation of Carbon Stable Isotope from Different Tissues in Yellow Tail Seriola aureovittata
HAN Yu-jia1,2,TIAN Jia-shen2,LI Duo-hui3,WANG Zhen1,2,LU Zhi-chuang2,Mu Yun-lei2
(1.Dalian Ocean University,Dalian 116023,China;2.Liaoning Key Laboratory of Marine Biological Resources and Ecology,Liaoning Ocean and Fisheries Science Research Institute,Dalian 116023,China;3.Dalian Fisheries Research Institute,Dalian 116085,China)
In this study,the conversion difference and fractionation coefficient of carbon stable isotope in different tissues were investigated in yellow tail Seriola aureovittata with body weight of(34.2± 1.8)g fed diets containing detected carbon source by stable isotope technique in a laboratory condition at water temperature of 18.79~22.15℃in order to understand the effects of tissue growth and metabolism on the relative contribution of the carbon stable isotope conversion in different tissues of yellow tail.The results showed that the minimal half-life of carbon stable isotope was observed in liver (3.2 d),followed by intestine (3.4 d),gill(3.6 d),heart(5.5 d),and muscle(6.4 d),and that the minimal enrichment coefficients of carbon stable isotope in liver and diet(0.15‰),followed by intestine(0.48‰),heart(0.98‰),gill(1.02‰)and muscle(1.08‰).The higher stable isotope conversion rates in liver and intestine were involved in high metabolic rate(>60%),thus relatively less contribution to growth.Liver and intestine can reflect the isotope alteration during the assimilation of dietary carbon in short-term feeding,while the muscle reflects the isotope alteration of long-term feeding.The findings improved the biology of yellow tail,and provided information for the stable isotope sample collection at different period,diet enrichment and selection in the aquaculture industry.
Seriola aureovittata;carbon stable isotope;isotope fractionation;conservation rate
S917
A
1005-3832(2017)06-0034-07
2017-06-05
辽宁省科技计划项目(2014203006);辽宁省海洋与渔业厅科研项目(201609).
韩羽嘉(1993-),男,硕士研究生,从事水产养殖研究.E-mail:luhuaqing@sina.cn
木云雷(1960-),男,副研究员,从事鱼类育种研究.E-mail:muyunlei@sina.com
