不同放牧强度下植物种群密度的季节性变化
2017-12-29单玉梅温超陈海军张璞进晔薷罕木兰常虹孙海莲
单玉梅, 温超, 陈海军, 张璞进, 晔薷罕, 木兰, 常虹, 孙海莲
不同放牧强度下植物种群密度的季节性变化
单玉梅1,2, 温超1,2, 陈海军3, 张璞进1,2, 晔薷罕1,2, 木兰1,2, 常虹1,2, 孙海莲1,2
1. 内蒙古自治区农牧业科学院, 呼和浩特 010031 2. 中国科学院内蒙古草业研究中心, 呼和浩特 010031 3. 内蒙古自治区生物技术研究院, 呼和浩特 010010
采用随机区组的试验方法, 着力于探索内蒙古荒漠草原3种主要植物种群密度在不同生长季节随放牧强度变化而呈现出季节性的波动规律。研究对象主要是短花针茅()、冷蒿()和无芒隐子草()。结果表明: 上述3种主要植物种群总密度在放牧区均高于对照区; 短花针茅和无芒隐子草枝条总密度的变化趋势基本一致, 冷蒿枝条总密度总体逐月降低。整体分析而言, 放牧期间(6—9月), 短花针茅、冷蒿和无芒隐子草枝条总密度的峰值均出现在HG(或MG), 说明放牧家畜的持续采食及践踏, 导致了植物种群株丛发生破碎而趋向小型化, HG(或MG)由母株破碎而产生的小株丛多, 故株丛总密度相应增加; 另外, 连续放牧致使大多数植株未能进行生殖生长即被采食, 从而促进了植物的营养繁殖和营养生长, 增加了种群密度。
放牧强度; 荒漠草原; 种群密度; 季节性
1 前言
荒漠草原处于草原与荒漠之间的过渡地段, 作为草原畜牧业主要生产资料地的内蒙古荒漠草原, 更是处于极特殊的地域, 是我国北部边疆的生态屏障, 其草原类型也属于最旱生的草原类型之一。植被种类比较贫乏, 以强旱生的小半灌木或矮禾草为主, 并且草层低矮、群落结构单一, 生态系统比较脆弱, 容易受到自然和人为因素的干扰, 存在退化的潜在危机[1–3]。然而, 在草原生态系统中, 最主要的利用方式是放牧, 放牧家畜通过采食牧草、践踏土壤和粪便回归直接影响着生态系统的结构与功能[4,5], 其中, 最直接的表现就是植物的种群生物学特征。
植物种群生物学特征对不同放牧强度的响应是放牧生态学研究的热点和焦点, 从放牧生态学的角度来讲, 通过对不同放牧强度下主要植物种群生物学特征指标的测定, 能够反映出随着放牧强度的变化植物种群的变化, 在草原生态系统的放牧管理中具有重要作用。在荒漠草原生态系统, 其地理位置、气候特点和植被类型均具有独特性, 季节的变化直接改变着系统的自然因子, 而人为因素中最主要的就是放牧, 因此季节的改变和放牧的干扰是脆弱的荒漠草原地区植被变化的关键驱动因子。已有的诸多研究都是关于植物种群生态学特征对一些自然因素和放牧干扰的可塑性响应[6–8], 而对于一个完整生长季的观测研究较少, 缺少季节的变化因子与放牧交互效应的动态规律。因此, 从放牧生态学的角度, 开展荒漠草原主要植物种群特征对放牧强度的季节性响应的研究, 可以加深了解荒漠草原生态系统中放牧强度对植物种群特征的季节性影响规律, 为认识放牧影响下荒漠草原退化的过程和机理提供理论依据, 为荒漠草原的合理利用和草地畜牧业的可持续发展提供保障[9]。
2 材料与方法
2.1 试验区概况
该试验样地位于内蒙古乌兰察布市四子王旗王府一队, 地理位置为41°47′17″N和 111°53′46″E, 平均海拔在1450 m左右, 气候属于典型的大陆性季风气候, 春季干旱, 大风频繁, 夏季炎热, 降雨量集中期为5月—8月, 多年平均降雨量为280 mm, ≥10 ℃积温为1900—2500 ℃, 平均无霜期为180 d, 平均温度最高的月份是6、7和8月。土壤类型为淡栗钙土, 距地表50—60 cm处多出现白色钙积层。草地类型为短花针茅+冷蒿+无芒隐子草的荒漠草原, 植被草层比较低矮, 一般在8 cm左右, 植被较稀疏, 盖度一般在15%—20%之间, 种类组成较贫乏, 群落主要由20多种植物组成, 建群种为短花针茅, 优势种为冷蒿和无芒隐子草。
2.2 试验设计
试验区设置在内蒙古农牧业科学院综合试验示范中心的长期放牧控制试验平台, 已具有连续10年的放牧历史。
试验区采用随机区组设计, 将试验区分成3个区组, 每个区组内设有1个无牧区和3个放牧区, 其中无牧区作为对照, 3个放牧区分别设3个放牧强度, 即总共设4个处理, 分别表示为对照(CK)、轻度放牧(LG)、中度放牧(MG)和重度放牧(HG), 放牧强度分别为: 0、0.93、1.82和2.71 只·hm-2, 区组作为重复, 共计12个试验区, 完全随机排列, 整个试验样地占地约为50 hm2, 每个试验区的面积基本相等, 分别表示为CK1、LG1、HG1和MG1; HG2、CK2、MG2和LG2; LG3、CK3、MG3和HG3, 轻度、中度和重度放牧试验区内的放牧羊只数量分别为4只、8只和12只(图1)。
2.3 测定方法
采用固定样方观测法。在每个试验小区内随机设置5个1 m2的观测样方, 在生长季(5—9月), 分别用涂有不同颜色的铁丝标记3种主要植物植株(或株丛), 即每次进行观测时, 固定观测上述被标记的短花针茅、冷蒿和无芒隐子草植株(或株丛)。观测时, 分别记录在各样方内出现的植物种群的枝条密度(营养枝和生殖枝)。取样时间分别为5月15日、6月3日、6月26日、7月20日、8月15日和9月16日, 每一个观测时间均在各试验区的固定样方内。
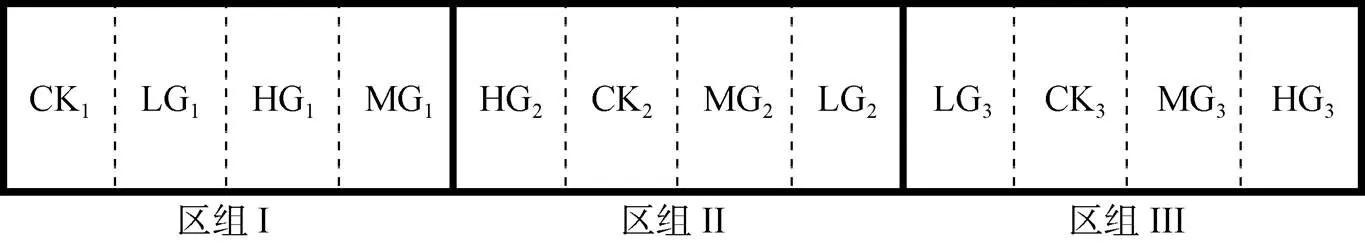
图1 放牧试验区完全随机区组分布示意图
2.4 数据处理分析
数据分析采用SAS9.0软件, 作图软件为Sigmaplot12.0。
3 结果与分析
3.1 放牧强度对种群总密度的影响
短花针茅、冷蒿和无芒隐子草种群总密度随着放牧强度的增加而变化。其中, 短花针茅种群总密度表现为HG>LG>MG>CK, 分别为13.4 株丛·m-2、10.4 株丛·m-2、10.1 株丛·m-2和8.6 株丛·m-2, 并且仅CK与HG间差异显著(<0.05)(图2)。
冷蒿种群总密度变化趋势与短花针茅相似, CK、LG、MG和HG分别为10.1株丛·m-2、11.3 株丛·m-2、11.2 株丛·m-2和14.9 株丛·m-2, 对照区与放牧区之间均存在显著性差异(<0.05)(图2)。
无芒隐子草种群总密度变化趋势则为LG>MG> HG>CK, 分别为15.3株丛·m-2、14.8株丛·m-2、12.7 株丛·m-2和10.3株丛·m-2, 且仅在LG与MG间存在不显著差异(>0.05), 其余处理间均存在显著性差异(<0.05)(图2)。
可见, 上述3种主要植物种群总密度在放牧区均高于对照区, 说明连续放牧和家畜的持续采食及践踏, 导致了植物种群株丛发生破碎而趋向小型化。
3.2 短花针茅种群株丛枝条(营养枝&生殖枝)总密度动态
短花针茅枝条总密度, 整个生长季在各放牧强度下均表现为先增加后降低的趋势, 最低值出现在5月, 峰值出现在6月末, 各月份间均存在显著差异(<0.05)。同一月份不同放牧强度间相比较, 5月(放牧前期), 枝条总密度随放牧强度的增加而降低, 彼此间均存在显著差异(<0.05); 6—8月, 枝条总密度在放牧区表现为随着放牧强度的增加而显著增加(<0.05)。9月, 则为MG>HG>CK>LG, 彼此间均存在显著差异(<0.05)(图3, A和B)。
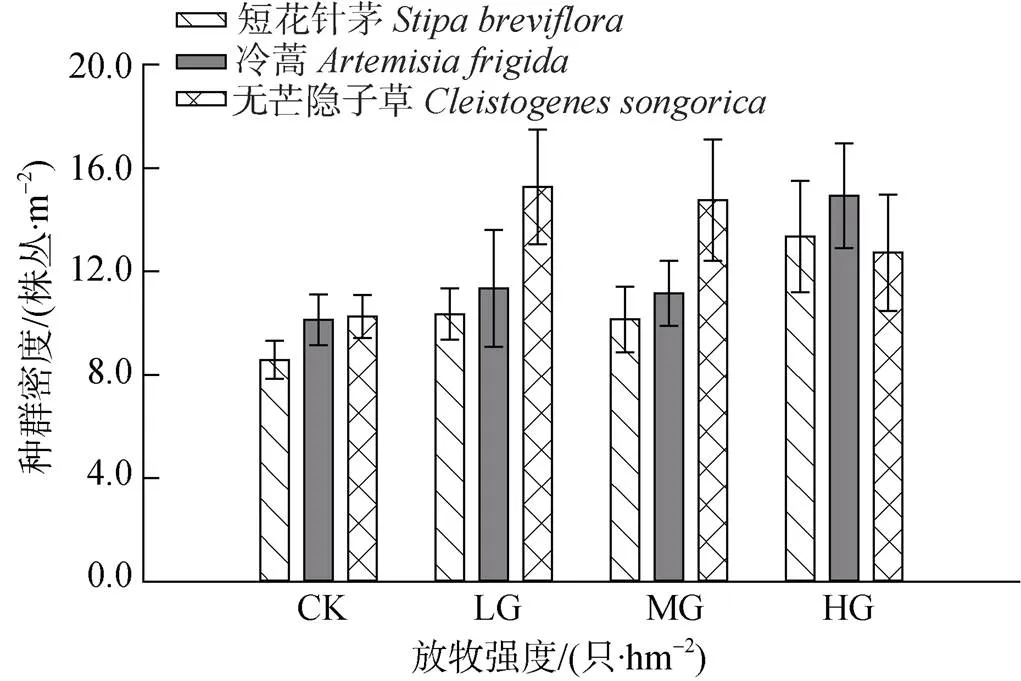
图2 不同放牧强度下短花针茅、冷蒿和无芒隐子草植物种群密度变化
3.3 冷蒿种群株丛枝条(营养枝&生殖枝)总密度动态
冷蒿枝条总密度, 整个生长季在各放牧强度下变化基本一致, 整体均呈现为逐渐降低的趋势, 各月间均存在显著差异(<0.05)。6月末, 其枝条总密度表现为随放牧强度的增加而降低, 可能原因为此时冷蒿处于营养生长旺期, 羊只乐于采食所致; 其他月份, 冷蒿枝条总密度在放牧区表现为随着放牧强度的增加而显著增加(<0.05)(图4, A和B)。
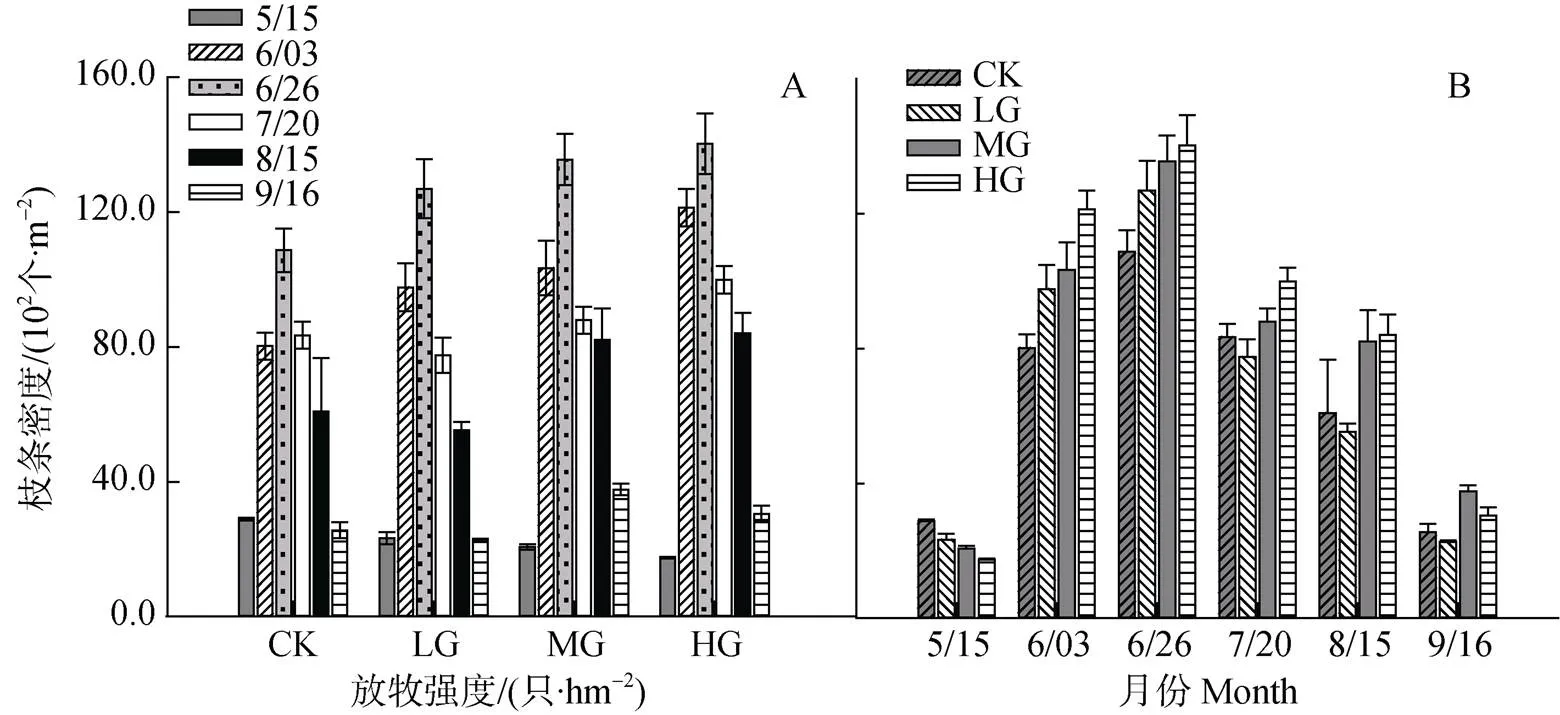
图3 不同放牧强度下短花针茅枝条总密度动态
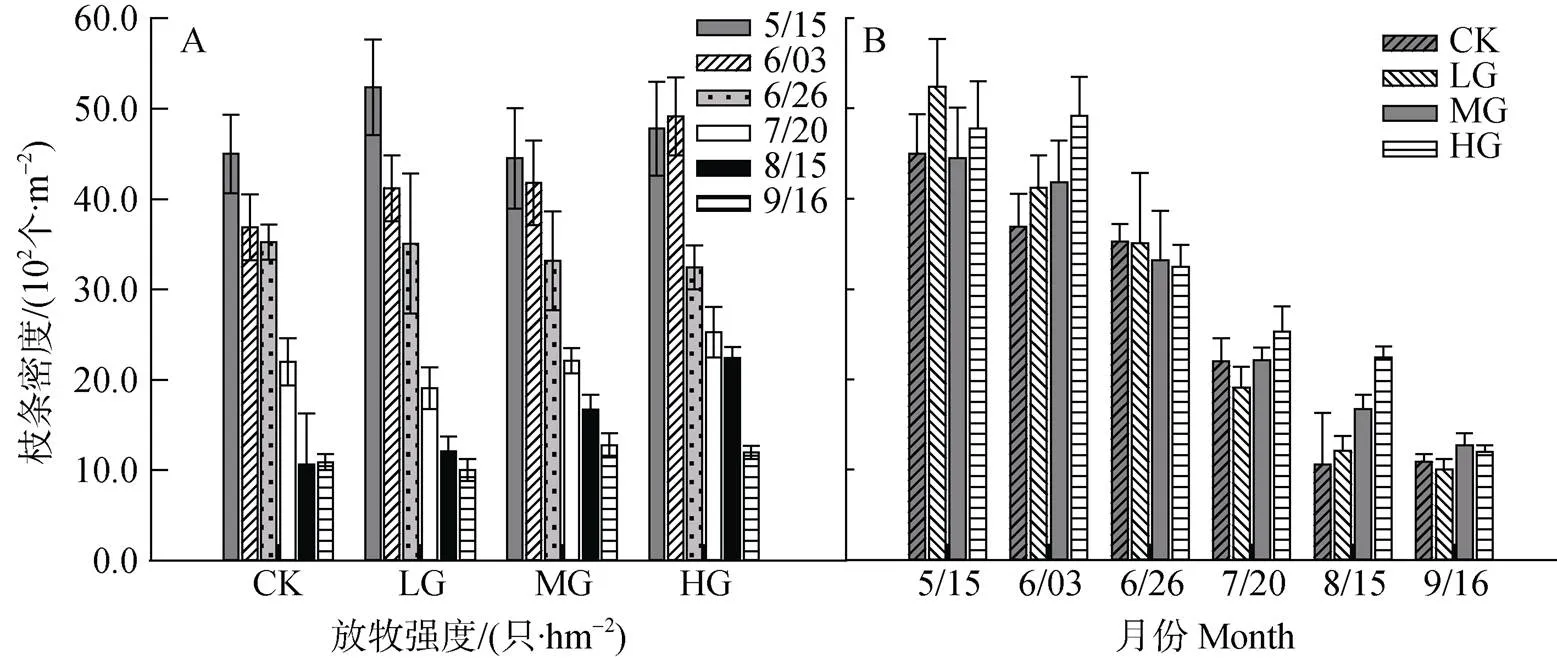
图4 不同放牧强度下冷蒿枝条总密度动态
3.4 无芒隐子草种群株丛枝条(营养枝&生殖枝)总密度动态
无芒隐子草枝条总密度, 在6—9月变化趋势一致, 呈现出先增加后降低的趋势, 最低值出现在9月, 各月份间均存在显著差异(<0.05)。5—8月, 同一月中, 枝条总密度均表现为随放牧强度的增加而增加, 彼此间差异均显著(<0.05)(图5, A和B)。
4 结论与讨论
放牧作为内蒙古荒漠草原的一种经济且简便的草地利用方式得到广泛应用, 也是该草地类型主要的人为干扰方式之一。放牧对草原植物的影响是多方面的, 常随着放牧强度、放牧方式和时间的变化而变化, 并且其影响具有累积效应[10–12]。草原植被对放牧强度的响应最直接的表现就是植物种群特征的变化, 比如种群密度的变化。因此, 草原植物种群特征反映了长期放牧对草原影响的结果, 不仅能指示草地状况、生产潜力、演替规律和承载能力等方面, 也能准确、客观、有效地阐释放牧生态学中的一些问题、现象及规律。
总体而言, 短花针茅和无芒隐子草枝条总密度的变化趋势基本一致, 冷蒿枝条总密度逐月降低。短花针茅、冷蒿和无芒隐子草枝条总密度的峰值均出现在HG(或MG), 主要是因为家畜的连续采食和践踏, 导致了短花针茅、冷蒿和无芒隐子草种群株丛的破碎化, 破碎的株丛即可脱离母株进行营养繁殖, 从而刺激了植物种群的营养繁殖和生长, 使种群得以维系, 这与白永飞等研究结果相一致[10,11,13–19]。种群株丛营养枝密度沿放牧强度呈现增加的趋势, 因此, HG种群营养枝密度有所增加。另一方面, 在HG下的家畜数量较多, 不断的反复采食, 致使大多数植株来不及抽穗、开花至结实, 使开花结实的株丛相应减少, 株丛进行光合所固定的能量多用于营养繁殖和营养生长, 因此营养枝密度必然增加, 从而增加了种群密度, 并且随着放牧强度的增加和放牧时间的持续, “分丛”现象也会更明显。
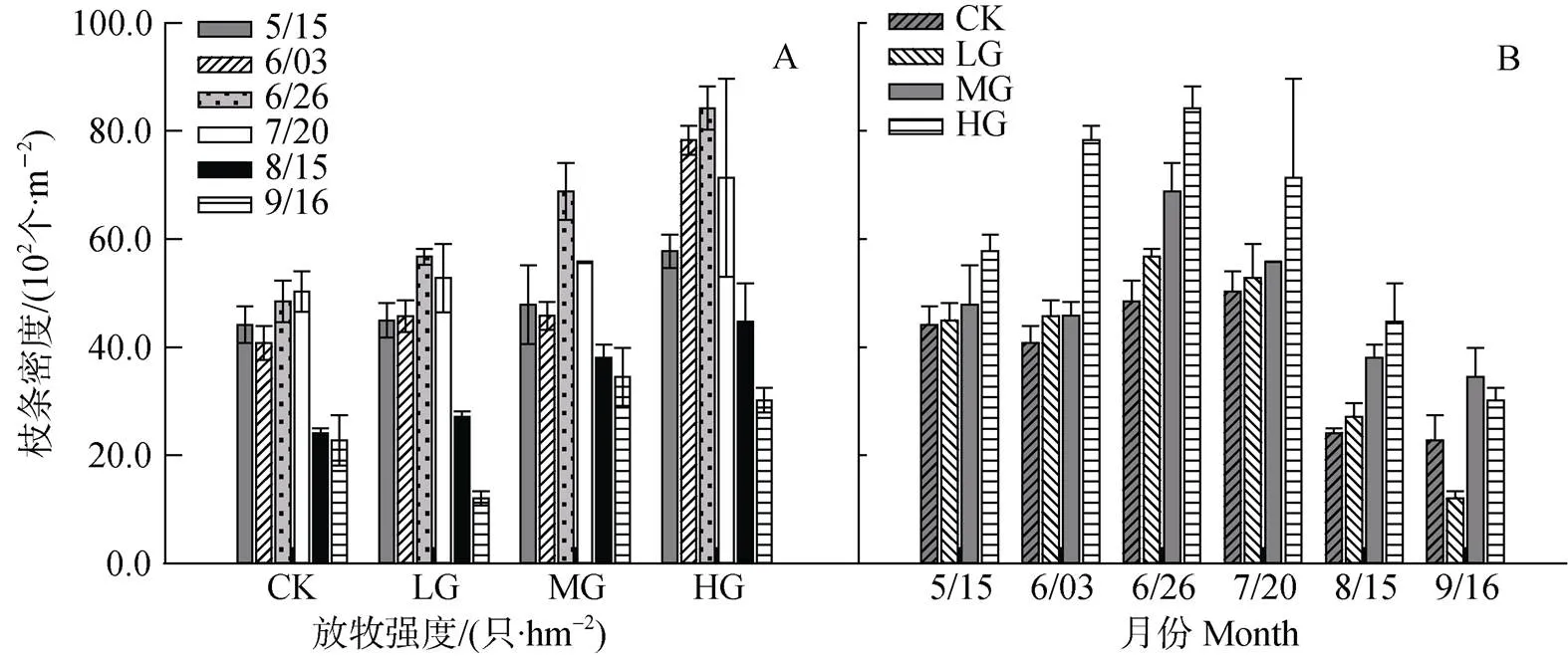
图5 不同放牧强度下无芒隐子草枝条总密度动态
从季节动态看, 整个生长季期间, 短花针茅枝条总密度整体呈现先升高后降低的趋势, 峰值出现在6月末。同一月份不同放牧强度之间比较, 6月初至8月, 短花针茅枝条总密度在放牧区均表现为随放牧强度增加而显著升高(<0.05), 9月, 各放牧强度间均无显著差异(>0.05); 原因可能为, 放牧在一定程度上刺激了短花针茅的分蘖, 促进其营养生长, 防止株丛老化, 使株丛枝条总密度与营养枝总密度保持适当比例, 具有相对较高的株丛密度。在生长旺季(7月—8月), 冷蒿与无芒隐子草枝条总密度均随着放牧强度的增加而增加。放牧强度越高, 家畜对冷蒿种群的践踏作用越强, 对冷蒿枝条芽的更新刺激作用越剧烈, 冷蒿的克隆生长也相应增强, 生长出更多的枝条, 致使冷蒿枝条密度明显增大, 这与杨持等研究结果相一致[20–23]。无芒隐子草是通过其分蘖节上的分蘖芽来增大地上枝条数量, 分蘖芽受放牧和季节的影响较大, 尤其是降雨的影响[24–26], 分蘖芽在较高放牧强度下受到的刺激作用明显增强, 分蘖出枝条数量也相应增多, 枝条总密度增加, 这与汪诗平和王艳芬等研究结果相一致[27]。
[1] 《内蒙古草地资源》编委会. 内蒙古草地资源[M]. 呼和浩特: 内蒙古人民出版社, 1990.
[2] 中国科学院内蒙古宁夏综合考察队. 内蒙古植被[M]. 北京: 科学出版社, 1985.
[3] 王玮, 刘钟龄, 郝敦元.内蒙古草原退化群落恢复演替的研究[J]. 植物生态学报, 1996, 20(5): 449–459.
[4] 陈佐忠, 汪诗平. 中国典型草原生态系统[Z]. 北京:科学出版社, 2000: 125–156.
[5] 陈海军, 单玉梅, 吴志毅, 等. 荒漠草原山羊和绵羊冬春季牧食行为及对草地的影响[J]. 干旱区资源与环境, 2010, 24(12): 172–178.
[6] WANG Z W, JIAO S Y, HAN G D, et al. Impact of stocking rate and rainfall on sheep performance in a desert steppe [J]. Rangeland Ecology Management, 2011, 64(3) : 249–256.
[7] 李永宏, 汪诗平. 放牧对草原植物的影响[J]. 中国草地, 1999, 21(3): 11–19.
[8] 董鸣, 张淑敏, 陈玉福. 匍匐茎草本蛇莓对基质养分条件的克隆可塑性[J]. 植物学报, 2000, 42(5) : 518–522.
[9] 任继周.草业科学研究方法[M] .北京:中国农业出版社, 1998.
[10] 卫智军, 常秉文, 孙启忠. 荒漠草原群落及主要植物种群特征对放牧制度的响应[J]. 干旱区资源与环境, 2006, 5(3): 33–38.
[11] 卫智军. 荒漠草原放牧制度和家庭牧场可持续经营研究[D]. 呼和浩特: 内蒙古农业大学, 2003.
[12] 李瑜清. 过渡放牧对生态环境的影响与控制对策[J]. 中国沙漠, 2005, 25(8): 404–408.
[13] 李永宏, 汪诗平. 草原植物对家畜放牧的营养繁殖对策初探[M]//中国科学院内蒙古草原生态系统定位研究站编. 草原生态系统研究(第5集). 北京: 科学出版社, 1997: 23–31.
[14] 白永飞, 李德新, 许志信, 等. 牧压梯度对克氏针茅生长和繁殖的影响[J]. 生态学报, 1999, 19(4): 479–484.
[15] 杜丽霞. 荒漠草原几种主要植物繁殖特性的研究[D]. 呼和浩特: 内蒙古农业大学, 2005.
[16] 李博文集编辑委员会.李博文集[C]. 北京:科学出版社, 1995: 154–158.
[17] 王忠武. 载畜率对短花针茅荒漠草原生态系统稳定性的影响[D]. 呼和浩特: 内蒙古农业大学博士学位论文, 2009.
[18] 汪诗平, 王艳芬. 不同放牧率下糙隐子草种群补偿性生长的研究[J]. 植物学报, 2001, 43(4): 413–418.
[19] 祁永. 放牧对草原群落特征及种群繁殖特性的研究[D]. 北京: 中国农业大学, 2005.
[20] 焦树英, 韩国栋, 刘俊清. 荒漠草原地区冷蒿构件及其动态规律对载畜率的响应[J]. 中国草地学报, 2009, 31(4): 80–84.
[21] 杨持, 宝音陶格涛, 李良. 冷蒿种群在不同放牧强度胁迫下构件的变化规律[J]. 生态学报, 2001, 21(3): 405–408.
[22] 王静, 杨持, 王铁娟. 放牧退化群落中冷蒿种群生物量资源分配的变化[J]. 应用生态学报, 2005, 16(12): 2316– 2320.
[23] 王静, 杨持, 王铁娟. 冷蒿()种群在放牧干扰下构件的变化[J]. 生态学报, 2006, 26(3): 960–965.
[24] 焦树英, 韩国栋. 短花针茅草原主要植物种群特征及其资源分配对载畜率的响应[J]. 中国草地学报, 2012, 34(2): 76–80.
[25] 杜丽霞. 荒漠草原几种主要植物繁殖特性的研究[D]. 呼和浩特: 内蒙古农业大学, 2005.
[26] 杜利霞, 李青丰, 董宽虎. 放牧强度对短花针茅草原冷蒿繁殖特性的影响[J]. 草地学报, 2007, 15(2): 367–370.
[27] 汪诗平, 王艳芬, 陈佐忠. 气候变化和放牧活动对糙隐子草种群的影响[J]. 植物生态学报, 2003, 27(3): 337–343.
单玉梅,温超,陈海军, 等. 不同放牧强度下植物种群密度的季节性变化[J]. 生态科学, 2017, 36(6): 125-129.
SHAN Yumei, WEN Chao, CHEN Haijun, et al. The seasonal variation of population density under different grazing intensities[J]. Ecological Science, 2017, 36(6): 125-129.
The seasonal variation of population density under differentgrazing intensities
SHAN Yumei1,2, WEN Chao1,2, CHEN Haijun3, ZHANG Pujin1,2, YE Ruhan1,2, MU Lan1,2, CHANG Hong1,2, SUN Hailian1,2
1. Inner Mongolia Academy of Agricultural and Animal Husbandry Sciences, Hohhot 010031, China2.Inner Mongolia Research Center for Prataculture, Chinese Academy of Sciences, Hohhot 010031, China3. Inner Mongolia Institute of Biotechnology, Hohhot 010010, China
The experiment was designed with a randomized complete block in the desert steppe in Inner Mongolia, to explore the seasonal variation rule of population density with grazing intensity changing. The objects of study were,and. The results showed that the three plant population densities in grazing plots were higher than control. The variation trend ofbunch density was accordance to, and that ofdecreased with month. During grazing season, from June to September, the peak values of the three plant bunch density were observed in HG plots. It illustrated that the continuous ingesting and trample resulted in clump broken, and trended to miniaturization to generate small clump to increase the population density. In addition, the continuous grazing caused that most of plants were ingested before reproductive growth, thus, it accelerated plant vegetative propagation and vegetative growth to increase the plant population density.
grazing intensity; desert steppe; population density; seasonal
10.14108/j.cnki.1008-8873.2017.06.017
S812
A
1008-8873(2017)06-125-05
2016-10-09;
2016-12-08
国家自然科学基金项目(31200351); 内蒙古自治区自然科学基金项目(2014MS0318); 内蒙古自治区自然科学基金项目(2015MS3084); 内蒙古农牧业科学院青年创新基金项目(2014QNJJM11)
单玉梅(1980—), 女, 博士, 助理研究员, 主要从事草地生态系统结构与功能方面的研究, E-mail:shanym2008@163.com
