池塘分区养殖系统对水质和浮游植物群落结构的影响
2017-12-29李俊伟朱长波颉晓勇郭永坚陈素文常华张宁曾林胡瑞萍
李俊伟, 朱长波, 颉晓勇,*, 郭永坚, 陈素文, 常华, 张宁, 曾林, 胡瑞萍
池塘分区养殖系统对水质和浮游植物群落结构的影响
李俊伟1, 朱长波1, 颉晓勇1,*, 郭永坚1, 陈素文1, 常华2, 张宁2, 曾林2, 胡瑞萍3
1. 中国水产科学研究院南海水产研究所, 广东省渔业生态环境重点实验室, 农业部南海渔业资源开发利用重点实验室, 广州 510300 2. 海大集团股份有限公司, 广州 511400 3. 广州市海威水产科技有限公司, 广州 510300
通过对虾池塘内浮游植物群落结构的生态学特征指标进行连续采样监测, 研究分区养殖系统对于虾塘水质和浮游植物群落组成的影响。采用进排水管和水泵将虾塘和鱼塘相连, 通过泵出虾塘底层水和回流鱼塘上层水实现水体流动。分区组(R)由对虾单养塘和鱼类混养塘组成, 对照组(C)为对虾单养塘, 监测时间为30天。结果显示: 通过定期循环处理可使对虾养殖池塘内总悬浮颗粒物(TPM)、颗粒有机物(POM)含量分别降低36%—45% 和15%—20%。R组虾塘水质改善, 溶解态活性磷含量(PO4–P)增加, 显著高于C组(<0.05); 实验结束时R组虾塘NH4+–N含量显著低于C组(<0.05)。实验期间, R组浮游植物种类多于C组, 均表现为绿藻门>蓝藻门>硅藻门>裸藻门; R组蓝藻生物量低于C组, 尤其C组具有较高比例的颤藻。实验结束时, R组绿藻门的优势度明显高于C组(<0.05); R组浮游植物的香农多样性指数(2.91)略高于C组(2.62)(>0.05)。实验期间, 2个处理组浮游植物群落的Margale丰富度指数均逐渐升高, 并且R组在第20、30 d高于C组(<0.05)。结果表明: 分区养殖可以通过减少颗粒物而改善水质, 水体流动可以加速含磷物质释放。水体流动和较低的N/P可能有助于提高虾塘内绿藻种类优势度而减少蓝藻发生, 并且在提高浮游植物种类多样性以及丰富度方面具有积极作用。
凡纳滨对虾; 混养; 分区养殖系统; 浮游植物
1 前言
凡纳滨对虾()是中国池塘主要养殖品种之一, 2015年中国凡纳滨对虾的养殖产量近162万吨[1]。然而, 养殖过程中大量的饲料投入和动物排泄等产生的有机质积累导致池塘内源性污染加剧, 容易引起对虾疾病的发生, 同时养殖废水的排放也加剧了周边环境的污染程度。研究发现对虾养殖系统内较大比例的氮磷物质不能被对虾利用[2]。因此, 针对养殖池塘内富含营养物质水体的净化修复技术和池塘养殖模式的优化研究日趋重要。
近年来, 对虾与不同生物种类的池塘混养模式研究取得较大进展, 如鱼—虾混养[3–5]、虾—蟹混养[6–7]以及虾—藻[8]等养殖模式的改进在一定程度上可以提高营养物质利用率和改善养殖水体[9]。以上研究多是在同一个养殖池塘中进行多品种的搭配养殖。分池环联养殖模式是将两种或两种以上的养殖池塘环联在一起, 使系统间产生互利作用或更好地利用空间资源, 达到高效、高产的目的[10], 可以避免空间、溶解氧竞争等[11–12]。广东湛江开展的虾—鱼—贝—藻多池循环水生态养殖模式可以提高养殖废水的利用和对虾的成活率, 具有生态环保等优点[13]。然而, 国内对虾养殖池塘格局和现状决定了传统养殖基础上简单的分区养殖模式更易为养殖户所接受。
对虾养殖初期, 池塘水体中由于缺乏营养物质而易导致“倒藻”现象发生[14], 因此需要及时补充富含营养物质和藻类的水体。对虾养殖后期, 池塘水体中有机质积累过多, 需要减少有机质的积累。对虾池塘水体中的颗粒有机物质可以作为滤食性鱼类的饵料, 溶解性营养物质可以供鱼类池塘内浮游植物生长所需, 成为滤食性鱼类的饵料组成。因此, 在对虾养殖初期将鱼塘中稳定的水体抽入到对虾池塘中以保证其水体稳定; 在对虾养殖中后期, 将池塘内富含颗粒有机物的水体抽入到鱼类养殖池塘中, 缓解对虾池塘中的有机物质积累, 同时也可提高营养物质在鱼类生长过程中的物质利用率, 主要途径是通过增加生物多样性、改善养殖系统内食物网结构[15], 从而提高养殖系统内能量和物质利用效率。
本研究采用进排水系统将对虾养殖池塘和鱼塘相连, 通过输出和回流实现两种养殖池塘中的物质和能量流动。该研究通过连续采样监测对虾池塘内浮游植物群落结构的生态指标变化, 以研究该分区养殖系统对于改善对虾池塘水质和浮游植物群落组成的影响, 以期为对虾养殖过程中浮游植物调控提供理论依据。
2 材料与方法
2.1 实验池塘与设计
分区水产养殖系统包括1口对虾单养池塘和1口鱼类混养池塘, 通过水泵和进排水管道将鱼塘与虾塘相连接(图1)。对虾池塘内水泵安置在距离底部15 cm水层, 将池塘内富含颗粒物质的水体泵入到鱼塘, 而鱼塘中的水泵位于水体上层, 将富含浮游植物且有机颗粒物较少的水体泵回虾塘, 循环系统运行周期为5天一次。对虾养殖池塘面积为4亩, 鱼类混养池塘面积为8亩, 池塘平均水深为1.5 m, 池塘内对角安置2台水车式增氧机, 中央位置安放一台涡轮式增氧机。采用3套分区养殖系统作为处理组, 3口对虾单养池塘作为对照组; 对虾(2.20 g)养殖密度为40000尾·亩-1, 鱼类池塘放养鲢鱼(480 g·ind-1)、鲫鱼(150 g·ind-1)、鲻鱼(65 g·ind-1)和草鱼(500 g·ind-1), 其密度分别为65 ind·亩-1、98 ind·亩-1、1588 ind·亩-1、18 ind·亩-1。

注1: 水泵; 2: 输水管道
实验期间, 对虾养殖池塘投喂相同的饲料, 日投喂量按照对虾存塘量的5%—7%进行投喂。鱼类混养池塘在实验初期仅依靠泵入的颗粒物质, 在实验后期投喂草鱼饲料以提供营养物质。
2.2 样品的采集、测定与计算

式中,P=n/;n为物种的个体数;为群落样品总个体数;为群落中的物种总数;f为该种在该地区出现的频率。
浮游植物密度计算公式:= (C/(F×F)) ×(/) ×P
式中:为1 L水中浮游植物的丰度(cell·L-1),C为计数框面积(mm2),F为一个视野的面积(mm2),F为计数过的视野数,为1 L水样经沉淀浓缩后的体积(mL),为计数框容积(0.1 mL),P为在F个视野中所计数到的浮游植物个体数。
此外, 每次采样水体中氨氮(NH4+–N)、亚硝态氮(NO2–N)、硝态氮(NO3–N)、活性磷(PO4–P)含量按照《养殖水化学实验》进行测定[16]。叶绿素(Chl.)按照海洋监测规范(2007)进行测定[17]。总颗粒物质(TPM)及颗粒有机物质含量(POM)通过烘干及550℃灼烧方法获得[18]。水体中不同粒径颗粒组成比例采用贝克曼库尔特颗粒计数仪测定(Multisizer 3, 贝克曼库尔特有限公司, 美国)。实验期间, 池塘水体溶解氧(DO)、温度()和pH采用YSI进行现场测定, 池塘水体的透明度采用萨式透明度盘测定。
实验结束时, 对虾的成活率(Survival rate)采用网捕方法进行计算。喂料1 h后, 在池塘内3个采样点处分别抛网, 根据网面积与数量、逃跑系数来估算。计算公式: 存活率=平均每网捕获数÷平均捕获面积×池塘面积×/放养量(为逃跑系数,=1.2)。同时, 称取60尾对虾总重求得平均重量。
2.3 数据统计
采用Excel和SPSS17.0进行数据处理和统计分析。数据均表示为平均数±标准误差, 处理组之间进行单因素方差分析(one-way ANOVA), 以<0.05作为差异显著水平。图标中不同小写字母代表差异显著(<0.05)。
3 结果
3.1 池塘水体营养盐含量变化
实验期间, 对照组和分区组对虾养殖池塘具有相近的水温、盐度、溶解氧和pH变化。池塘水体温度变化范围为29.0—31.4 ℃, 盐度变化范围为0—3, 溶解氧含量变化为4.0—8.0 mg·L-1, pH变化范围分别为7.7—9.3。实验初期, 两个处理组对虾养殖池塘水体的透明度变化范围为21—23 cm, 而在实验中期和后期, 分区组池塘内透明度增加至30 cm, 且显著高于对照组(<0.05)。在实验后期, 对照组池塘透明度一直处于较低水平, 主要原因可能是池塘内蓝藻发生导致透明度较低。
实验初期, 对照组和分区组池塘内的亚硝态氮、硝态氮、氨氮和活性磷含量均无显著差异(>0.05), 随着养殖时间增加和实验处理因素的影响, 池塘内的氮、磷营养盐含量均发生变化(图2)。实验结束时, 分区组池塘内亚硝态氮含量低于对照组(<0.05), 然而在实验中期采样阶段, 分区组亚硝态氮含量也具有高于对照组的现象(图2a); 实验中后期, 分区组池塘内硝态氮含量显著高于对照组(<0.05)(图2b); 实验后期, 分区组池塘内氨氮含量低于对照组且达到显著差异(<0.05)(图2c)。实验期间, 分区组对虾养殖池塘中活性磷含量表现出升高的趋势, 并且在实验中后期显著高于对照组(<0.05) (图2d)。

图2 实验期间池塘水体中营养盐含量变化
3.2 池塘水体中颗粒物质的特征变化
实验期间, 对照组和分区组对虾养殖池塘水体中总颗粒物质含量(TPM)变化范围分别为55—90 mg·L-1和35—60 mg·L-1, 平均值分别为70 mg·L-1和49.5 mg·L-1。实验结束时, 分区组池塘中TPM含量显著低于对照组(<0.05)(图3 a)。实验期间, 对照组池塘水体中的颗粒有机物质含量(POM)具有升高的趋势, 在实验结束时分区组池塘中的POM含量也有所增加, 但是在第3和4次采样阶段仍然显著低于对照组(<0.05)(图3b)。由TPM和POM数据可知, 对虾池塘水体分区处理有助于悬浮颗粒物质含量(TPM和POM)的降低。
实验期间, 采用贝克曼库尔特颗粒计数仪测定水体中粒径为20 μm—100 μm的颗粒物质数量。对照组和分区组对虾池塘内的颗粒物质数量变化范围分别是17.7×104—24.1×104个·L-1和12.2×104—19.6× 104个·L-1, 分区组中颗粒物质平均含量(15.8±3.7)×104个·L-1低于对照组(21.6±3.4)×104个·L-1。

图3 实验期间池塘水体中总颗粒物质含量和颗粒有机物含量
实验期间, 对照组和分区组对虾养殖池塘内颗粒物质(20—100 μm 粒径)主要由20—60 μm的物质组成, 而60—100 μm颗粒物质一直处于较低比例。随着实验时间的增加, 2个处理组中粒径为20—40 μm的颗粒物质表现出增加的趋势, 而粒径为40—60 μm的颗粒物质则表现出减少的趋势, 且实验结束时分区组中20—40 μm的颗粒物质明显高于对照组(图4)。结果表明, 分区循环处理有助于减少对虾池塘中的颗粒物质含量。
3.3 叶绿素
实验期间, 对照组和分区组对虾养殖池塘内Chl.含量均表现出上升的趋势, 2个处理组的变化范围分别为135—297 μg·L-1和140—255 μg·L-1; 实验结束时,对照组Chl.含量(297±32) μg·L-1高于分区组(255±23) μg·L-1, 但是未达到显著差异(>0.05)(图5)。
3.4 浮游植物鉴定结果
实验期间, 对照组和分区组对虾养殖池塘内分别鉴定出浮游植物门类5个和4个, 对照组共15属18种, 而分区组24属32种, 并且均表现为绿藻门类>蓝藻门类>硅藻门类>裸藻门类(表1)。
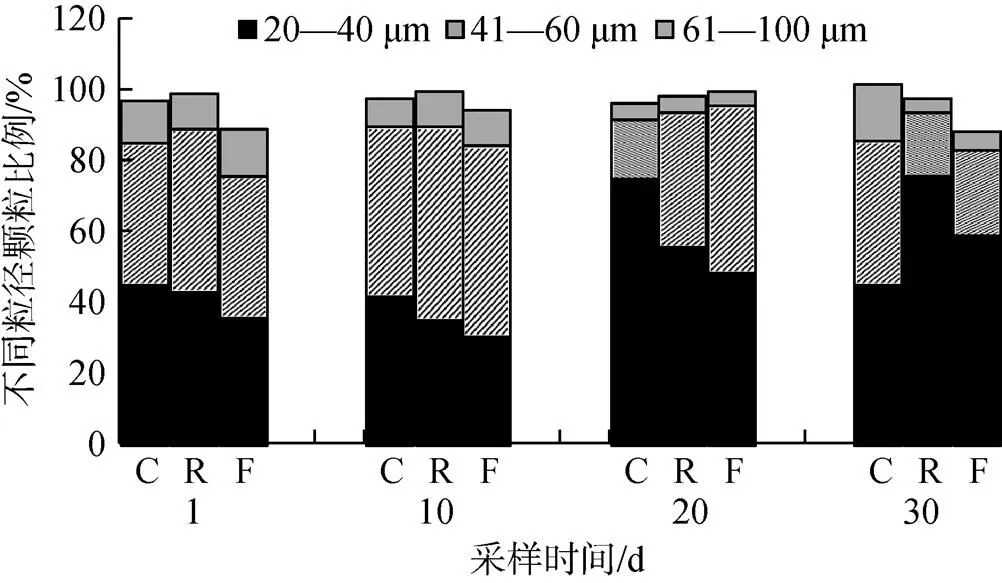
图4 实验期间不同粒径颗粒物质含量变化

图5 实验期间池塘水体中Chl. a 含量
实验初期, 对照组和分区组池塘水体中具有相近的浮游植物群落结构组成, 随着循环处理次数增加, 两个处理组池塘内的浮游植物群落结构组成发生明显变化。实验期间第10 d, 对照组和分区组具有相近的硅藻种类组成, 均是梅尼小环藻为主要种类; 然而, 分区组对虾养殖池塘内绿藻种类明显多于对照组, 分区组池塘中的空星藻()、波及卵囊藻()、被甲栅藻()和四尾栅藻()具有较高的生物量, 而对照组池塘内小孢空星藻为优势种类; 分区组池塘内蓝藻种类多于对照组, 伪鱼腥藻和针晶蓝纤维藻在2个处理组中均为优势种类, 而银灰平裂藻在分区组池塘中占较大优势, 却未在对照组池塘发现该藻; 对照组池塘内具有较大比例的颤藻, 而未在分区组池塘中发现颤藻。
实验期间第30 d, 2个处理组池塘内浮游植物群落结构组成表现出更大的差异。硅藻门中的梅尼小环藻在2个处理组池塘中均有发现, 而对照组池塘中的密度高于分区组; 同时, 分区组池塘中占较大比例的小环藻未在对照组发现。实验结束时, 对照组池塘中的颤藻占较大比例, 而分区组2口池塘中未发现颤藻, 另一口池塘仅有少量颤藻发生。对照组池塘中绿藻种类仍低于分区组, 四尾栅藻在2个处理组中均为优势种类, 其密度为288×104ind·L-1—480×104ind·L-1。
3.5 浮游植物群落结构特征指数
实验期间, 对照组和分区组池塘水体中硅藻门的优势度指数均表现出逐渐降低的趋势, 而2个处理组中蓝藻门的优势度表现出升高的趋势, 在第2个采样阶段分区组和对照组的硅藻门种类优势度和蓝藻门种类优势度均出现显著差异(图6 a, b)。在第20—30 d, 分区组池塘水体中绿藻门的优势度明显增加, 且在实验结束时高于对照组(>0.05), 只是其优势度指数低于蓝藻门(图6c)。2个处理组中裸藻门种类的优势度均较低, 且并未在整个实验期间连续发现裸藻门种类(图6d)。

表1 实验期间对虾养殖池塘中浮游植物种类组成

续表

图6 硅藻门(a)、蓝藻门(b)、绿藻门(c)、裸藻门(d)的优势度指数
实验结束时, 对照组和分区组池塘水体中的浮游植物优势种类优势度具有明显不同。蓝藻门中的伪鱼腥藻和银灰平裂藻分别在对照组和分区组具有最大优势度, 其优势度分别达到0.434和0.453(表格2)。对照组池塘内硅藻门类梅尼小环藻的优势度高于分区组; 对照组和分区组池塘内的绿藻门相同优势种类中仅有四尾栅藻, 而对照组中的其它优势种为葡萄藻和双列栅藻, 分区组中的其它优势种类为小球藻和二形栅藻(表格2)。
实验开始时, 对照组和分区组池塘内浮游植物群落的香农多样性指数均接近2.25,期间表现出逐渐升高的趋势, 在实验结束时对照组和分区组分别上升到2.62和2.91。整体而言, 实验期间分区组的物种多样性指数略高于对照组(>0.05)。实验期间, 分区组对虾养殖池塘内浮游植物的Pielou物种均匀度指数低于对照组, 且在第2个采样阶段出现显著差异(<0.05)。实验中后期, 两个处理组中浮游植物群落的Margale丰富度指数均表现出逐渐升高的趋势, 并且分区组在中期和末期显著高于对照组(<0.05) (图7)。

表2 实验结束时不同处理组浮游植物优势种类优势度
3.6 对虾的生长性能
实验期间, 通过网捕方法, 计算池塘内对虾的存活率。实验结束时, 对照组池塘对虾的存活率为78%, 而分区组池塘对虾的存活率为81% (表 3)。实验期间, 分区组对虾平均增重为7.02 g, 而对照组对虾平均增重为6.68g。
4 讨论
4.1 池塘分区养殖对水体营养盐的影响
分池环联养殖系统定期将对虾养殖池塘内富含残饵、碎屑和细菌等有机物质的水体转移到鱼类净化池塘中, 可以减轻对虾养殖池塘内有机负荷和保持水质稳定[12]。胡庚东等(2011)研究发现淡水池塘循环水养殖模式经过3级净化后, 养殖排水水质符合《地表水环境质量标准》V类标准, 并且对Chl.的去除效果明显[19]。池塘—湿地循环水养殖研究也表明池塘循环水对于TN、TP、NH4+–N及COD均有明显的去除率[20]。研究结果显示, 分区养殖可以改善对虾养殖池塘内水质。实验期间, 池塘分区养殖显著提高对虾池塘内的活性磷含量(<0.05), 在一定程度上可以降低NH4+–N含量, 且在实验结束时分区组池塘水体NH4+–N含量显著低于对照组(< 0.05)(图2)。活性磷含量的增加可能是由于下层水体的流动造成活性磷转化与释放加速; 颗粒有机物质的移除可能是影响分区组水体中NH4+–N含量低于对照组的主要因素。混养滤食性鱼类对于浮游植物和颗粒物质的利用会加速水体中氮磷物质的循环[21], 由实验结果表明, 分区养殖过程中引起的颗粒物减少和水体流动也加速了水体中磷的转化与循环。

图7 香农多样性指数(a)、Pielou物种均匀度指数(b)、Margalef丰富度指数(c)

表3 各处理组对虾成活率、终末体重量
4.2 池塘分区养殖对颗粒物质、叶绿素a含量的影响
对虾养殖池塘内的颗粒有机物主要来源于对虾残饵、粪便以及浮游生物等, 养殖中后期较多的颗粒有机物不利于池塘水质稳定, 尤其对虾精养池塘TPM和POM逐渐升高更易导致水质较差[18, 22]。分池环联养殖模式是将两种或两种以上的养殖池塘环联在一起, 使系统间产生互利作用或更好地利用营养物质[10], 而减少养殖动物间的相互影响[12]。申玉春等(2007)开展了多池循环水生态养殖研究, 对虾养殖废水经过贝类池塘后, 水体中颗粒物质含量降低, 由104.6 mg·L-1降低到45.4 mg·L-1。结果表明, 池塘分区养殖处理有助于悬浮颗粒物质(TPM和POM)的移除和降低(图3)。依据循环系统运行时混养鱼类对泵入颗粒物质的摄食现象可知颗粒物质可作为混养池塘内滤食性鱼类的饵料。分区组池塘内悬浮颗粒物质平均含量(49.5±10.6) mg·L-1低于对照组(70±15.2) mg·L-1, 可知, 通过定期循环处理可使对虾养殖池塘内TPM、POM含量分别降低36%—45% 和15%—20%。
通过贝克曼库尔特颗粒计数仪测定水体中粒径为20—200 μm的颗粒物质含量, 分区组对虾池塘水体中的颗粒物质平均含量为15.8×104个·L-1, 而对照组为21.6×104个·L-1, 结果表明池塘分区循环处理可以降低悬浮颗粒物质数量。养殖期间, 粒径小于20 μm的小型颗粒物质在总悬浮颗粒物中占较大比例[23], 由于未对20 μm以下的颗粒物质进行统计, 因此实验数值小于浮游植物丰度和实际数量。然而, 通过粒径组成分析, 可知20—160 μm的颗粒物质占较大比例, 并且可受到循环处理而减少。实验期间, 分区组和对照组对虾养殖池塘内Chl.含量均增加, 分区组Chl.含量低于对照组, 但是未达到显著差异(>0.05)(图5), 结果表明, 分区养殖使得颗粒物质含量降低, 提高了水体透明度, 但是并未明显提高浮游植物生物量。对照组中Chl.含量较高可能与实验期间蓝藻生物量较大有关。
4.3 池塘分区养殖对浮游植物的影响
环境因子对浮游植物群落的影响是多方面的, 而且极其错综复杂。池塘中的水温、溶解氧、pH及营养物质等因素的高低均对浮游植物的生长、繁殖、种群变化起着直接作用[21, 24]。本实验中, 不同处理组池塘间的t、DO、pH较为接近, 而TPM、溶解态营养盐以及水体流动是影响浮游植物群落结构组成的主要因素(图2, 3)。杨慧君等(2010)研究发现鱼—稻循环系统养鱼池塘内浮游植物种类(93种)、密度(247—1133)×104及生物量(3.07—10.83) mg·L-1均低于非循环塘100种, (511—1773)×104, (4.48—17.34) mg·L-1[25]。朱晓荣(2013)研究发现循环系统运行初期养殖池塘内浮游植物种类数量也少于对照区池塘和外河水源, 随着养殖时间增加和水质逐渐稳定, 循环组养殖池塘内的浮游植物种类增多[26]。本实验中, 分区组池塘内浮游植物种类(32种)高于对照组(18种), 主要原因可能是分区组池塘内氮磷物质转化和释放加速, 且活性磷含量明显增加(图2), N/P更有利于多种浮游植物的生长。一般情况下养殖水体属于磷限制水体[27], 而鱼—稻循环养殖系统内由于水稻对磷的利用率较高[25], 经稻田循环利用后仍属于磷限制水体, 因此循环系统养鱼池塘内浮游植物种类少于对照组。实验中分区组池塘循环水流动加速了活性磷的转化和释放, 有利于多种浮游植物发生。
分区组浮游植物种类多于对照组, 且绿藻门种类最多, 表现为绿藻门类>蓝藻门类>硅藻门类>裸藻门类(表1)。一般情况下, 养殖中后期绿藻门类占优势[25,28]。分区组池塘内蓝藻生物量低于对照组, 尤其对照组具有较高比例的颤藻, 而仅在分区组一口池塘中发现少量颤藻。在同一地区, 光照与水温基本相同, 浮游植物群落结构主要决定于营养盐类[27]。浮游植物可以利用系统底部再生的氮磷营养盐[29], 浮游植物群落结构与磷酸盐具有密切关系[14], 浮游植物体内氮磷比约为16:1[30]。N/P较大时, 蓝藻易占优势。N、P均丰富时, 绿藻易占优势[27]。分区组N/P范围为5.77—20.33, 平均为9.92; 而对照组N/P范围为10.57—51, 平均为28.01, 因此, 实验池塘中浮游植物种类变化可能与水体氮磷含量比值具有密切关系。同时, 分区组池塘内具有较高含量的银灰平裂藻, 而对照组池塘内未发现该藻。浮游植物群落结构的生态指标也表明池塘分区养殖处理对于虾塘浮游植物群落组成具有影响。实验后期, 分区组池塘水体中绿藻门的优势度明显高于对照组(<0.05)(图6c)。对照组蓝藻门类的伪鱼腥藻具有最大优势度, 而分区组内银灰平裂藻优势度最大, 分别达到0.434和0.453(表2), 分区组池塘内蓝藻门优势种类(银灰平裂藻、伪鱼腥藻)的总优势度明显低于对照组(针晶蓝纤维藻、颤藻、伪鱼腥藻)。实验期间分区组浮游植物的香农多样性指数(2.91)略高于对照组(2.62)(>0.05)。实验中后期, 2个处理组浮游植物群落的Margale丰富度指数均表现出逐渐升高的趋势, 并且分区组在实验期间第20 d和第30 d显著高于对照组(<0.05)(图7)。结果表明, 池塘分区循环养殖在提高浮游植物种类多样性以及丰富度方面具有一定作用。
5 结论
分池环联养殖模式是将两种或两种以上的养殖池塘环联在一起, 使系统间产生互利作用, 达到高效、高产的目的, 可以避免空间、溶解氧及食物竞争等。然而, 国内对虾养殖池塘格局和现状决定了传统养殖基础上简单的环联模式。在对虾养殖初期将鱼塘中较为稳定的水体抽入到对虾池塘中以保证水质稳定; 在养殖中后期, 将池塘中富含颗粒有机物的水体抽到鱼类池塘中, 缓解对虾池塘中的有机物质积累,提高了营养物质利用率; 同时, 减少养殖废水排放, 有利于生态环境保护。实验结果表明, 该分区养殖系统对于改善对虾池塘水质和浮游植物群落组成具有重要影响。分区处理可明显降低悬浮颗粒物质含量, 提高水体透明度。池塘水循环引起的颗粒物减少和水体流动也加速了水体中磷的转化与释放。同时, 分区循环处理有利于增加对虾养殖池塘内的浮游植物种类, 提高生物多样性, 改善养殖系统内食物网结构。实验期间, 鱼类混养池塘饲料投喂量较低, 仅投喂少量草鱼饲料, 池塘水体中浮游植物生物量较低, 而循环系统中对虾养殖废水并不能够支撑整个鱼塘的浮游植物营养盐需求, 因此, 混养鱼塘需要投喂一定数量的植物型饲料以维持整个鱼塘浮游生物的稳定。通过实验发现, 混养鱼塘与对虾养殖池塘的面积比例较大, 因此两种池塘的比例需要进一步优化研究。
[1] 农业部渔业局. 中国渔业统计年鉴. 北京: 中国农业出版社, 2016.
[2] FUNGE-SMITH S J, BRIGGS, M R P. The origins and fate of solids and suspended solids in intensive marine shrimp ponds in Thailand[M]. Summary report from Sonykhla Region of Thailand. Institute of Aquaculture, Stirling, 1994.
[3] TIAN Xiangli, LI Deshang, DONG Shuanglin, et al. An Experimental study on Closed–polyculture of Penaeid Shrimp with Tilapia and Constricted tagelus[J]. Aquaculture, 2000, 202 (3/4): 57–77.
[4] 李卓佳, 虞为, 朱长波, 等. 对虾单养和对虾—罗非鱼混养试验围隔氮磷收支的研究[J]. 安全与环境学报, 2012, 4: 50–55.
[5] 陈辉煌, 陆开宏, 郑忠明, 等. 夏季混养鳙和银鲫对凡纳滨对虾低盐养殖池塘浮游藻类群落结构的影响[J]. 生态科学, 2014, 3: 459–466.
[6] 李信书, 王洪斌, 陈文慧, 等. 虾蟹混养池塘异养菌区系组成及群落结构[J]. 水产科学, 2013, 02: 106–109.
[7] 张凯, 田相利, 董双林, 等. 三疣梭子蟹、凡纳滨对虾和菲律宾蛤仔混养系统氮磷收支的研究[J]. 中国海洋大学学报(自然科学版), 2015, 2: 44–53.
[8] 王大鹏, 田相利, 董双林. 对虾、青蛤和江蓠三元混养效益的实验研究[J]. 中国海洋大学学报, 2006, 36: 20–26.
[9] 张凯. 三疣梭子蟹()混养系统有机碳收支以及细菌生产力与代谢功能的研究[D].青岛: 中国海洋大学, 2014.
[10] 董双林. 中国综合水产养殖的发展历史、原理和分类[J]. 中国水产科学, 2011, 18(5): 1202–1209.
[11] 黄国强, 李德尚, 董双林. 一种新型对虾多池循环水综合养殖模式[J]. 海洋科学,2001, 25(4) : 48–49.
[12] 申玉春, 熊邦喜, 王辉, 等. 虾—鱼—贝—藻养殖结构优化试验研究[J]. 水生生物学报, 2007, 31(1): 30–36.
[13] 申玉春, 叶富良, 梁国潘, 等. 虾—鱼—贝—藻多池循环水生态养殖模式的研究[J]. 湛江海洋大学学报, 2004, 24(4): 10–16.
[14] RAHMAN M M, NAGELKERKE LEOPOLD A J, VERDEGEM MARC C J, et al. Relationships among water quality,food resources,fish diet and fish growth in polyculture ponds: A multivariate approach[J]. Aquaculture, 2008, 275: 108–115.
[15] YOKOYAMA H, HIGANO J, ADACHI K, et al. Evaluation of shrimp polyculture system in Thailand based on stable carbon and nitrogen isotope ratios[J]. Fisheries Science, 2002, 68: 745–750.
[16] 雷衍之. 养殖水环境化学实验[M]. 北京: 中国农业出版社, 2006: 80–85.
[17] 中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会. GB12763.6-2007海洋监测规范. 北京:中国标准出版社, 2008, 3–10.
[18] 李俊伟, 朱长波, 颉晓勇, 等. 对虾养殖池塘内混养鲻鱼和罗非鱼对水环境及对虾生长的影响[J]. 生态科学, 2015, 34(6): 86–92.
[19] 胡庚东, 宋超, 陈家长, 等. 池塘循环水养殖模式的构建及其对氮磷的去除效果[J]. 生态与农村环境学报, 2011, 27(3): 82–86.
[20] 彭刚, 刘伟杰, 童军, 等. 池塘循环水生态养殖效果分析[J]. 水产科学, 2010, 29(11): 643–647.
[21] 谷孝鸿. 不同养殖类型池塘浮游生物群落结构的初步分析[J]. 湖泊科学, 1994, 6(3): 276–282.
[22] 陈金玲, 赖秋明, 苏树叶, 等. 精养虾池主要生态因子变化特点与相关性分析[J]. 南方水产科学, 2012, 8(4): 49–56.
[23] 蔡建楠, 潘伟斌, 王建华, 等. 鲢和奥尼罗非鱼对池塘蓝藻水华及水质的影响[J]. 水生态学杂志, 2008,1(1): 56–61.
[24] CASÉ M, LEÇA E E, LEITÃO S N, et al. Plankton community as an indicator of water quality in tropical shrimp culture ponds [J]. Marine Pollution Bulletin, 2008, 56(7): 1343–1352.
[25] 杨慧君, 谢从新, 何绪刚, 等. 循环水池塘与非循环水池塘中浮游生物的群落结构及其动态研究[J]. 淡水渔业, 2010, 40(3): 28–35.
[26] 朱晓荣. 基于潜流湿地的池塘循环水养殖系统净化效能研究[D]. 苏州: 苏州大学, 2013.
[27] 董双林. 养殖水域生态学[M]. 北京: 中国农业出版社, 2004: 189–201.
[28] 郭永坚, 朱长波, 阴晓丽. 凡纳滨对虾—鲻网围分隔混养池塘浮游植物群落结构特征的研究[J]. 南方水产科学, 2015,11(1): 45–54.
[29] BRIAN C P, JOHN W D, DUBRAVKO J, et al. Nitrogen and phosphorus transport between Fourleague Bay, LA, and the Gulf of Mexico: the role of winter cold fronts and Atchafalaya River discharge[J]. Estuarine, Coastal and Shelf Science, 2003: 1065–1078.
[30] REDFIELD A C, KETCHUM B H, RICHARDS F A. The influence of organisms on the composition of seawater// HILL M N. The Sea, vol. II, 1963: 26–77.
李俊伟, 朱长波, 颉晓勇, 等. 池塘分区养殖系统对水质和浮游植物群落结构的影响[J]. 生态科学, 2017, 36(6): 114-124.
LI Junwei, ZHU Changbo, XIE Xiaoyong, et al. Characteristics of water quality and phytopankton community in the partitioned aquaculture system[J]. Ecological Science, 2017, 36(6): 114-124.
Characteristics of water quality and phytopankton community in the partitioned aquaculture system
LI Junwei1, ZHU Changbo1, XIE Xiaoyong1,*, GUO Yongjian1, CHEN Suwen1, Chang Hua2, ZHANG Ning2, ZENG Lin2, HU Ruiping3
1. Guangdong Provincial Key Laboratory of Fishery Ecology and Environment, Key Laboratory of South China Sea Fishery Resources Exploitation & Utilization of Ministry of Agriculture of China, South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Guangzhou 510300, China 2.Haida group co., LTD, Guangzhou 511400, China 3. Haiwei aquaculture technology, LTD, Guangzhou 510300, China
The effects of a partitioned aquaculture system on the water quality and phytoplankton were studied using 3 partitioned units and 3 control ponds. The partitioned aquaculture system consisted of shrimp pond and fish pond, and the two ponds were connected by piping and pump. Three other isolated shrimp ponds were designated as the C group, whereas the fish pond was used for the culturing of silver carp, crucian carp, grass carp andand three partitioned aquaculture system were designated as the R group (R). Throughout 4 sampling times (30d), the total particulate matter (TPM) and the particulate organic matter (POM) in the R group were 36%-45% and 15%-20% lower than in the C group. The results suggest that the system can improve the water quality in the shrimp ponds, the PO4–P content of the water column was higher in the R group than in the C group (<0.05), and NH4–N was lower than in the C group (<0.05) late in the experiment. During the experiment, the phytoplankton species in the R group was more than that in the C group, and the trend of the different species in the two groups was Chlorophyta>Cyanophyta>Bacillariphyta>Euglenophyta. The biomass of Cyanobacteria in the R group was less than that in the C group, and there was higher density ofin the C group. At the end of the experiment, the Simpson index of Chlorophyta in the R group was higher than that in the C group (<0.05), and the Shannon index of phytoplankton in the R group was a little more than that in the C group (>0.05). During the experiment, the richness index of phytoplankton in the two groups increased gradually, and the value in the R group was higher than that in the C group on the 20thday and 30thday (<0.05). In the present study, the partitioned aquaculture system could be easily implemented in an aquaculture zone, and its application can reduce the POM and increase the release of PO4–P. The running water and low N/P value may be the main reason for the dominance of Chlorophyta and weakness of Cyanophyta, therefore the partitioned aquaculture system may play an important role in the improvement of the diversity of phytoplankton.
; polyculture; partitioned aquaculture system; phytoplankton
10.14108/j.cnki.1008-8873.2017.06.016
S96
A
1008-8873(2017)06-114-11
2016-10-10;
2016-12-08
广东省海洋渔业科技推广项目(B201601-01&B201500B04); 中央级公益性科研院所基本科研业务费专项资金(中国水产科学研究院南海水产研究所) ( 2014ZD01); 广东省省级科技计划项目(2013B090500081&2013B040500020)
李俊伟(1982—), 男, 河北故城人, 博士, 助理研究员, 从事水产生态养殖技术研究, E-mail: lijunwei303@163.com
颉晓勇, 副研究员, 水产养殖技术研究, E-mail: xiexiaoyongsh@sina.com
