虫纹鳕鲈消化道指数及主要消化酶活性的研究
2017-12-26安丽孟庆磊李娴董学飒付佩胜朱永安
安丽,孟庆磊,李娴 董学飒,付佩胜,朱永安
(山东省淡水渔业研究院,山东 济南 250117;山东省淡水水产遗传育种重点实验室,山东 济南 250117)
虫纹鳕鲈消化道指数及主要消化酶活性的研究
安丽,孟庆磊,李娴 董学飒,付佩胜,朱永安
(山东省淡水渔业研究院,山东 济南 250117;山东省淡水水产遗传育种重点实验室,山东 济南 250117)
以体重(1.02±0.34)kg,体长(34.7±5.2)cm的虫纹鳕鲈(Macculochellapeeli)为实验材料,采用常规方法测量其消化道指数并对3种主要消化酶的活性及分布进行了研究。结果表明,虫纹鳕鲈的比肠长、比肠重、比肝脏重和比内脏重分别为0.557±0.028、0.0047±0.0005、0.0157±0.008和0.058±0.005。胃蛋白酶在各消化器官的比活力顺序为胃>肝胰脏>肠;淀粉酶的比活力顺序为肝胰脏>胃>肠;脂肪酶比活力顺序为肝胰脏>肠>胃。研究结果表明,虫纹鳕鲈具有肉食性鱼类的消化道特征,胃为胃蛋白质的主要消化器官,肝胰脏为淀粉的主要消化器官,肝胰脏和肠为脂肪的主要消化器官。
虫纹鳕鲈(Macculochellapeeli);消化道指数;胃蛋白酶;淀粉酶;脂肪酶
虫纹鳕鲈(Macculochellapeeli)俗称澳洲石斑或墨累鳕鱼,隶属于鲈型目(Periformes)鳕鲈科(Maccullochellidae)鳕鲈属(Maccullochella)。原产于澳大利亚,由于产量大、肉质细腻、口味鲜美、营养价值高,是当地重要的淡水养殖品种。国外学者早在20世纪70年代就对虫纹鳕鲈各方面特征[1~5]进行了研究,国内关于虫纹鳕鲈的研究比较晚也比较薄弱,目前仅对其生物学特性[6]、形态结构[7,8]、肌肉营养成分[9]、人工养殖技术[10]、苗种培育技术[11]及遗传多样性[12]有过初步研究,至今未见关于虫纹鳕鲈消化道指数、消化酶活性的研究报道,但这方面的研究结果对于了解虫纹鳕鲈的食性、消化特点都有重要的意义,可进一步丰富虫纹鳕鲈消化生理基础资料,为其人工配合饲料的开发提供有力的科学依据。
1 材料与方法
1.1 材料
实验所用虫纹鳕鲈引自澳大利亚,暂养于山东省淡水渔业研究院,随机选取2龄虫纹鳕鲈(体重(1.02±0.34)kg,体长(34.7±5.2)cm)共30尾用于实验。
1.2 方法
1.2.1消化道指数测定
取样前24h内停止投喂,取样后测定体长、体重后,解剖鱼体,取出内脏,分离消化道和肝胰脏,生理盐水冲净内容物,剥去多余的脂肪和结缔组织,滤纸吸干水分后分别称重(精确到0.01g),测量肠道的长度(精确到0.1cm),后用于酶液的制备。参照尾崎久雄的方法测定虫纹鳕鲈的比肠长(肠长与体长之比)以及比肠重、比肝脏重和比内脏重(分别为肠重、肝脏重、内脏重与体重之比)等消化道指数。
1.2.2酶液的制备
以2尾鱼的同一部位合并为1个样本,将所获样本加入4倍体积(W/V)预冷双蒸水,用玻璃匀浆器在冰盒中匀浆。匀浆液经高速冷冻离心机离心30min(10000r/min),取上清用于酶活力测定,4℃下保存,24h内分析完毕。每个样本设3个重复。
1.2.3酶活力测定
脂肪酶、胃蛋白酶、淀粉酶的活性测定及蛋白浓度的测定均使用南京建成生物工程研究所生产的试剂盒。
酶的活力以比活力表示,单位为酶活力单位/mg蛋白(U/mgprot)。
1.3 数据处理
采用SPSS 16.0对数据进行单因素方差分析及多重检验,显著水平以P<0.05,极显著水平以P<0.01表示。
2 结果与分析
2.1 虫纹鳕鲈的消化道指数
30尾虫纹鳕鲈的消化道指数测量结果见表1。鱼体解剖后,消化道完全排列在腹腔中,消化器官中肝脏和胃所占比重较大,其次是肠。肝脏具有3个不完全分叶,且左分叶明显大于其他2个分叶。胃“U”型,壁厚,胃肠之间具幽门盲囊。肠道相对粗短,呈“S” 型弯曲,比肠长为0.557±0.028。

表1 虫纹鳕鲈消化道指数
2.2 虫纹鳕鲈的消化酶活性
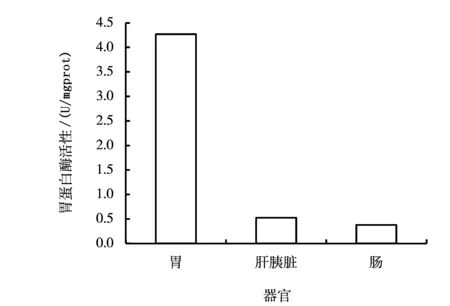
图1 虫纹鳕鲈胃蛋白酶活性比较
虫纹鳕鲈消化道各部分的胃蛋白酶活性见图1。由图1可见,虫纹鳕鲈胃中的胃蛋白酶活性最强,为(4.271±0.403)U/mgprot;肝胰脏和肠道的胃蛋白酶活性较低,分别为(0.525±0.091)U/mgprot和(0.383±0.054)U/mgprot。胃与肝胰脏、胃与肠道的胃蛋白酶活性差异均达到了极显著水平(P<0.01);肝胰脏和肠道的胃蛋白酶活性差异不显著(P>0.05)。
虫纹鳕鲈消化道各部分的淀粉酶活性见图2。由图2可见,虫纹鳕鲈肝胰脏中的淀粉酶活性最强,为(0.285±0.025)U/mgprot;其次为胃和肠道,分别为(0.163±0.008)U/mgprot和(0.082±0.010)U/mgprot。肝胰脏和胃、肠道之间的淀粉酶活性差异均达到了极显著水平(P<0.01);胃和肠道之间的淀粉酶差异达到显著水平(P<0.05)。
虫纹鳕鲈消化道各部分的脂肪酶活性见图3。由图3可见,虫纹鳕鲈肝胰脏脂肪酶活性最强,为(0.365±0.055)U/mgprot;其次为肠道和胃,分别为(0.236±0.038)U/mgprot和(0.084±0.012)U/mgprot。胃和肝胰脏脂肪酶活性差异达到极显著水平(P<0.01);胃和肠道及肝胰脏和肠道脂肪酶活性差异均达到显著水平(P<0.05)。

图2 虫纹鳕鲈淀粉酶活性比较 图3 虫纹鳕鲈脂肪酶活性比较
3 讨论与结论
3.1 虫纹鳕鲈消化道指数与食性
鱼类通过消化系统完成对食物的摄取、营养物质的吸收和消化,进一步获得用于生长、发育、繁殖等一系列生命过程所需要的营养和能量。根据摄食方式和食物种类的不同,鱼类大体分为植食性鱼类、杂食性鱼类和肉食性鱼类。不同食性鱼类因消化食物的特性不同,在消化系统的形态和结构上也存在很大的差异。因此,消化道指数如比肠长、比肠重、比肝重、比内脏重等可谓是鱼类食性最直接的反映指标。一般肉食性鱼类肠长为体长的1/3~1/4;草食性鱼类肠长为体长的2~5倍,甚至达15倍;杂食性鱼类的肠短于草食性鱼类而长于肉食性鱼类[13]。肉食性鱼类的比肠长较短,一般小于1;杂食性和植食性鱼类的比肠长较长,多数都大于1,潘黔生等[14]、毕冰等[15]和林浩然等[16]都通过研究不同鱼类的消化系统结构证明了这一点。肉食性鱼类内脏团较小,比内脏重小于杂食性或草食性比内脏重,肉食性大银鱼的比内脏重为0.038[17],明显小于偏植食性长鳍篮子鱼的比内脏重为0.157[18]。肝脏为鱼体最大的消化腺,其分泌胆汁与胰腺分泌的胰液可用于分解动物性食物,因此肉食性鱼类比肝重较大于杂食性或草食性鱼类的比肝重[19]。王亚楠[20]比较了4种淡水经济鱼类的消化系统,植食性鱼类黄尾密鲴和鲢鱼的比肝重为0.017和0.020,均大于肉食性鱼类翘嘴红鲌和黄鳝的比肝重(0.0066和0.0122)。虫纹鳕鲈的平均比肠长为0.557小于1,平均比内脏重为0.058,平均比肝重为0.0157,各消化道指数与肉食性鱼类相接近,显示了其肉食性鱼类的消化道特征。
3.2 虫纹鳕鲈消化酶活性比较
3.2.1胃蛋白酶活性分布特点
虫纹鳕鲈胃中胃蛋白酶活性极显著高于肝胰脏和肠道,肝胰脏和肠道中的胃蛋白酶活性很低,两者之间几乎无差别。鱼类消化道中存在酸性蛋白酶(胃蛋白酶、组织蛋白酶等)和碱性蛋白酶(胰蛋白酶、胰凝乳蛋白酶等),酸性蛋白酶在酸性条件下活性较高,主要存在于胃中[21,22],碱性蛋白酶在碱性条件下活性较高,主要存在于肠道中[23]。 有研究表明[24,25],先以不具有活性的酶原颗粒储存在胃的细胞中,待到有酸或有活性的蛋白酶作用下转变为具有活性的胃蛋白酶,胃蛋白酶是有胃鱼类胃中作用最强的消化酶;肝胰脏主要分泌蛋白酶原而未被激活,导致蛋白酶活性较弱;肠道分泌肠致活酶,能激活蛋白酶原,可对食物中的蛋白质进行消化吸收,但是它需要在碱性条件下表现出较高的碱性蛋白酶活性,因此只在胃中检测出较高胃蛋白酶活性。
3.2.2淀粉酶活性分布特点
淀粉酶在几乎所有的鱼类中都有存在,在消化道的分布及活性根据鱼的种类和食性的不同存在着较大的差异。杨金海等[18]对长鳍篮子鱼的消化酶研究结果表明,淀粉酶活性肠道最高,幽门盲囊次之,胃较低,肝脏则很少。马燕梅等[26]对鳜的消化酶研究结果表明,肝脏淀粉酶活性最高,幽门盲囊次之,胃较低,肠道则几乎没有。李凤杰等[27]对长薄鳅幼鱼的消化酶研究结果表明,肝胰脏的淀粉酶活性极显著高于其余各器官,胃淀粉酶活性显著低于肝胰脏和肠道的淀粉酶活性。虫纹鳕鲈淀粉酶活性从肝胰脏、胃、肠呈递减的趋势,三者之间差异都达到显著水平,可推测出虫纹鳕鲈肝胰脏、胃和肠在淀粉酶的消化中都起一定的作用,其中肝胰脏为淀粉的主要消化器官。
3.2.3脂肪酶活性分布特点
有研究表明,脂肪酶活力的大小与鱼类的食性无明显的相关性,与其摄食的食物组成有一定的相关性[28],其分泌的部位器官也存在差异。Mankura等[29]研究的7种海水鱼脂肪酶的活性都以幽门盲囊最高,陈苏维等[30]研究发现泥鳅的肝胰脏脂肪酶活性最高,且与其他部位存在显著差异,吴仁协等[31]研究发现中华乌塘鳢肠脂肪酶活性显著高于食道、胃和肝脏。虫纹鳕鲈肝胰脏脂肪酶活性最强,其次为肠道,胃较低,且三者之间差异均达到显著水平,三者均参与食物中脂肪的消化,肝胰脏和肠为虫纹鳕鲈脂肪的主要消化器官。
3.3 结论
对虫纹鳕鲈消化道不同部位脂肪酶、胃蛋白酶和淀粉酶活性的测定,结果显示脂肪酶活性顺序为:肝胰脏>肠道>胃,肝胰脏和肠道对脂肪的分解消化具有重要的作用;胃蛋白酶活性顺序为:胃>肝胰脏>肠道,且胃中胃蛋白酶活性极显著的高于肝胰脏和肠道,则说明胃可能是虫纹鳕鲈蛋白质消化的主要场所;淀粉酶活性顺序为:肝胰脏>胃>肠道,肝胰脏中淀粉酶活性极显著高于胃和肠道,说明肝胰脏是虫纹鳕鲈生成淀粉酶的的重要器官。
鱼类消化酶的组成特点与其食性有密切的关系,不同的学者[32,33]在研究肉食性、草食性和杂食性鱼类各消化酶活性时发现肉食性鱼类脂肪酶活性和蛋白酶活性较高,草食性鱼类淀粉酶活性较高。Biesio等[34]认为可采用A/P值(淀粉酶比活力/蛋白酶比活力的值)做为鱼类食性的指标,A/P值>1时,鱼类为草食性或偏草食性,A/P值<1时,鱼类为肉食性或偏肉食性。本研究结果表明,虫纹鳕鲈的A/P值<1,符合虫纹鳕鲈为肉食性鱼类的结论。
[1]Gooley G J, Anderson T A, Appleford P. Aspects of the reproductive cycle and gonadal development of Murray cod,Maccullochellapeeliipeelii(Mitchell) (Percichthyidae), in Lake Charlegrark and adjacent farm ponds, Victoria, Australia[J]. Marine and Freshwater Research,1995,46: 723~728.
[2] Anderson J R, Morison A K, Ray D J. Age and growth of Murray cod,Maccullochellapeelii(Perciformes : Percichthyidae), in the Lower Murray-Darling Basin, Australia, from thin-sectioned otoliths[J]. Australian Journal of Marine and Freshwater Research,1992, 43: 983~1013.
[3]Silva S, Silva S S D, Gunaseera R M,etal. Digestibility and amino acid availability of three protein-rich ingredient-incorporated diets by Murray codMaccullochellapeeliipeelii(Mitchell) and the Australian shortfin eel Anguilla australis Richardson[J]. Aquaculture Research,2000,32:195~205.
[4] Robert E K, Melissa A K. The status of Murray cod in the Murray-darling basin[D]. Canberra: Applied Ecology Research Group,University of Canberra, 2001.
[5] Rowland S J. Overview of the history, fishery, biology and aquaculture of Murray cod (Maccullochellapeelii) [A] .Menagement of Murray cod in the Murray-darling Basin-canberra[C].Canberra:Murray-Darling Basin commission,2004:38~61.
[6] 韩茂森.澳洲虫纹鳕鲈的生物学特性及引养前景[J].淡水渔业,2003,33(4):50~52.
[7] 王波,张艳华,韩茂森.虫纹麦鳕鲈的形态和生物学性状[J].水产科技情报,2003,30(6):266~267.
[8]安丽,董学飒,张延华,等.虫纹鳕鲈外形特征及内部消化系统结构的研究[J].长江大学学报(自然科学版),2013,10(17):29~32.
[9]宋理平,冒树泉,胡斌,等.虫纹鳕鲈肌肉营养成分分析与品质评价[J].饲料工业, 2013, 34 (16): 42~45.
[10]李娴,朱永安,钟君伟,等.虫纹鳕鲈的生物学特性及人工养殖技术研究[J].湖北农业科学,2013,52(9):2114~2115.
[11]左瑞华,汪学军.虫纹鳕鲈苗种培育影响因素初步分析[J].安徽农学通报,2001,7(3):57~59.
[12]张龙岗,杨玲,李娴,等.利用mtDNACOI基因序列分析引进的澳洲虫纹鳕鲈群体遗传多样性[J].水产学杂志,2013,26(2):14~18.
[13]冯昭信,曹启华,李星颉,等.鱼类学[M]. 北京:中国农业出版社,1979.
[14]潘黔生,郭广全,方之平,等.6种有胃真骨鱼消化系统比较解剖的研究[J].华中农业大学学报(自然科学版), 1996,15(5):463~469.
[15]毕冰,孙中武,毛天强,等.鲤、鲢、鳙、草鱼消化道结构与食性的研究[J].水产学杂志, 2011, 24(1):26~29.
[16] 林浩然. 五种不同食性鲤科鱼的消化道[J]. 中山大学学报(自然科学版), 1962,3:65~78.
[17]钱云霞,杨文鸽.路开宏.大银鱼消化道指数和消化酶的初步研究[J].水利渔业,2001,21(5):8~9.
[18]杨金海,章龙珍,庄平,等.人工养殖长鳍篮子鱼消化道指数及3种消化酶活性分布[J].海洋科学, 2009,33(7):43~48.
[19]熊洪林,戴婧婧,陈嶙.中国少鳞鳜消化系统解剖结构研究[J].安徽农业科学,2011,39(7):4015~4017.
[20]王亚楠.四种淡水经济鱼类消化系统的比较研究[D].长沙:湖南农业大学,2008.
[21] Einarsson S, Davis P S. On the localization and ultrastructure of pepsinogen, trypsinogen and chymotrypsin secreting cells in the Atlantic salmon,SalnosalarL[J].Comp Biochem physio, 1996,114:295~301.
[22] Chiu S T, Pan B S. Digestive protease activities of juvenile and adult eel (Anguilljaponica) fed with floating feed [J]. Aquaculture,2002,205:141~156.
[23] Kawai S, Ikeda S. Studies on digestive enzymes of fish Ⅱ.Effect of dietary change on the activities of digestive enzymes in carp intestine[J]. Bulletin of the Japanese Society of Scientific Fisheries,1972,38:265~270.
[24]周景祥,陈勇,黄权,等.鱼类消化酶的活性及环境条件的影响[J].北华大学学报,2001,2(1):70~73.
[25] Das K M, Tripathi S D. Studies on the digestive enzymes of grass carp, Ctenopharyngodon idella[J].Aquaculture,1991,92:21~30.
[26]马燕梅,梅景良,林树根.鳜胃肠道和肝脏主要消化酶活性的研究[J].江西农业大学学报, 2004, 26 (4):584~588.
[27]李凤杰,曾令清,黄小铭,等.长薄鳅幼鱼消化道指数和消化酶活性研究[J].淡水渔业, 2012, 42 (3):10~13.
[28]路明龙,朱希,赵卫红.鲈鱼、草鱼和鲤鱼的消化酶活性比较研究[J].上海农业科技,2008,(5):58~59.
[29] Nakamura M, Kayama M, Saito S.Wax ester hydrolysis by lipolytic enzymes in pyloric caeca of various fishes[J].Nippon Suisan Gakkaishi,1984,50:2127~2131.
[30]陈苏维,吉红,朱文东,等.泥鳅消化道指数及消化酶活性分布的研究[J].水产科学, 2009,28(5):272~275.
[31]吴仁协,洪万树,张其永,等.中华乌塘鳢成鱼消化酶活性的研究[J].厦门大学学报(自然科学版),2007,46(1):118~122.
[32]Agrawal V P, Sastry K V, Kaushab S K. Disgestive enzymes of three teleost fishes[J]. Acta Physiol Acad Sci Hung,1975,46:93~98.
[33]吴婷婷,朱晓鸣.鳜鱼、青鱼、草鱼、鲤、鲫、鲢消化酶活性的研究[J].中国水产科学,1994,1(2):10~17.
[34]Biesiot P M, Capuzzo J M. Change in digestive enzyme activities during early development of the American lobsterHomarusamaricanus, IW[J]. Mar Biol Ecol,1990,136:107~122.
2017-06-16
农业部“948”项目(2011-z41);国家科技基础条件平台项目(2016DKA30470);山东省现代农业产业技术体系鱼类创新团队(SDAIT-12-01)。
安丽(1982-),女,硕士,助理研究员,主要从事淡水鱼类遗传育种研究。通信作者:朱永安,zhuyongan1965@163.com。
[引著格式]安丽,孟庆磊,李娴,等.虫纹鳕鲈消化道指数及主要消化酶活性的研究[J].长江大学学报(自科版),2017,14(22):32~35.
S917
A
1673-1409(2017)22-0032-05
[编辑] 余文斌
