两种连作模式下杨树苗木的气体交换及其生物量分配
2017-12-20黄延军朱文成安小林张鹏远董玉峰
黄延军,朱文成,安小林,任 飞,张鹏远,董玉峰∗
(1.山东省水利科学研究院,山东 济南250013;2.山东省林业科学研究院;3.山东省林木种苗和花卉站)
两种连作模式下杨树苗木的气体交换及其生物量分配
黄延军1,朱文成2,安小林2,任 飞2,张鹏远3,董玉峰2∗
(1.山东省水利科学研究院,山东 济南250013;2.山东省林业科学研究院;3.山东省林木种苗和花卉站)
以I-107杨为试材,采取当年扦插(A)和留根截干(B)两种不同的连作模式,研究两种不同连作方式下杨树苗木的气体交换及生物量分配。结果表明:A模式下,光合速率(Pn)、蒸腾速率(E)及气孔导度(Gs)均随连作代数的增加而降低,水分利用效率(WUE)则相反.B模式下,Pn和WUE随着连作代数的增加而降低,E相反,而Gs则先增加后降低。其中,A模式下1代、2代苗木的Pn、E及Gs均大于B模式;随着连作代数的增加,两种不同连作方式下的苗高、胸径及单株生物量均呈现下降趋势。其中,B连作模式下的苗高、胸径及单株生物量均高于同代的A连作模式;两种连作方式下的叶绿素a、b含量及叶绿素总量均随连作代数增加而降低。其中,A模式下苗木的叶绿素a、b含量及叶绿素总量高于B模式;随着连作代数的增加,A、B两种连作方式下根、茎部分生物量均降低,但两种连作模式下各部分生物量所占比例的变化趋势相反。其中,B连作模式下苗木的根、茎生物量明显高于A模式。
连作代数;杨树;苗木;气体交换;生物量分配
长期以来苗圃承担着壮苗培育、科学试验和示范推广等任务,为造林绿化和林业六大工程建设做出了积极的贡献。但是由于土地资源有限,大多数的苗圃面临着多代连作的问题,加上长期经营管理不善,树种单一等问题使得苗圃地面临着严重的地力衰退现象[1]。苗圃地连作引起地力下降的问题很早就有报道[2-3],且目前仍然是困扰苗木生产的一个大问题。但由于耕地面积的不断减少,连作往往不可避免。连作研究在杉木、桉树等树种方面开展较早,研究的也比较深入[4-6],有报道指出不同立地管理措施会对连作地上杉木林的生长产生较大影响[7,8],那么不同的连作方式是否能减缓或改变苗圃地的这种衰退现象,地力衰退所引起的苗木生长矮小、瘦弱及产量低多与苗木的生物量积累有关,苗木的气体交换过程是生物量积累的重要生理生化过程。为此,笔者以两种不同的连作模式下不同连作代数的I-107杨树为研究对象,对不同模式下各代苗木的气体交换及影响气体交换的外界环境因子及内部光合器官中叶绿素含量,光合产物的分配比例等做了深入的调查研究,以期能了解不同连作模式下各代苗木的气体交换和生物量分配情况,为苗圃地不同模式连作经营提供技术支持和理论依据。
1 材料与方法
1.1 试验地概况
试验地位于山东省兖州国有苗圃,地处鲁西南平原的东北部 (116°44′-45′E,35°35′-37′N),海拔49 m。成土母质为山前冲洪积物,由古汶河和泗河冲积形成,表层全部为第四纪松散岩覆盖,土壤黏质,通透性较差。该处属暖温带东亚季风区,年平均气温13.6℃,极端最高气温为41℃,极端最低气温为-19℃。平均无霜期为203 d,最长232 d,最短182 d,平均日照时数2 610.7 hr。平均年辐射总量为124.04 K.cm-2。年平均降雨量725.9mm,最多降雨量为1 179mm,最少降雨量为406mm
1.2 试验地选择
分别选取地理位置和立地条件均一致,不连作、连作1代和连作2代的苗圃地作为试验样地,采取两种不同的连作方式:当年扦插连作方式(A连作模式):即在未连作,连作1次,连作2次的试验地上采取扦插的方法进行育苗,苗木、插穗及根系均为第1年生;留根截干连作方式(B连作模式):即在第1年扦插育苗,收获时采取截干措施,收获地上部分,地下部分保留,等待第2年抽条生长,其中插穗、根系为两年生,茎干为当年生。试验中采用随机区组设计,两种连作方式下的其他育苗和抚育管理措施都按生产常规,但力求一致与可比。
1.3 测定方法及数据分析
(1)生长量调查 采用机械抽样方法,每小区50样株,重复3次,分别调查各试验地内杨树苗木的苗高、胸径。
(2)气体交换 光合作用的测定采用CIRAS-2型光合仪直接野外活体测定,测量在野外天气晴朗的上午8:00—11:30之间进行,外界光强的变化范围在900~1200μmo L m-2·s-1,确保了在饱和光强之下。每小区随机抽取6株,每株选择3片完全展开的健康叶片测定其光合速率(Pn)、蒸腾速率(E)、气孔导度(Gs)等气体交换参数,同时测定空气相对湿度(Rh),大气温度(Tc),光合有效辐射(Par),大气CO2浓度(Ca)等环境参数。使用开放气路,空气流速为0.5 L· min-1。水分利用效率(WUE)= 净光合速率(Pn)/蒸腾速率(E)。
(3)叶绿素的测定 收集光合作用测定后的新鲜叶片,每小区共18片,做标记后立即放入冰壶内带回实验室用于叶绿素的测定。叶绿素的测定采用80%丙酮法黑暗中提取24 h,然后用UV-2500型紫外分光光度计测定.
(4)生物量的测定 每小区选取6样株,地上部分采取分层切割法,地下部分采用全挖法[9],即当根挖至水平分布和垂直分布的边缘时,按根径大小分粗根和细根连同插穗一并收集。最后所有样品分成根、茎、叶、插穗等不同部分在105℃烘箱中连续烘干至恒重。
2 结果与分析
2.1 两种不同连作模式下各代苗木的气体交换参数。
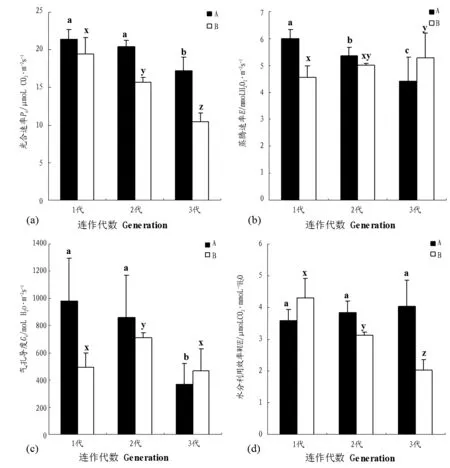
图1 两种连作模式下不同连作代数气体交换参数。Fig1 Gas exchange in different generations under two continuous cropping types.
随着连作代数的增加,两种连作方式下的杨树苗木的Pn均呈下降趋势,其中A连作模式下,1代与2代之间苗木的Pn没有显著差异 (图1a,P>0.05),但到了第3代苗木的Pn与第1代、第2代相比差异显著 (图1a,P<0.05)。在B连作模式下,3代苗木Pn之间差异均达到显著水平(图1a,P<0.05)。随着连作代数的增加,当年苗的E呈下降趋势,而B模式下苗木的E呈上升趋势。在A模式下,3代苗木的E差异显著(图1b,P<0.05),而在B模式下,仅第1代与第3代苗木的E差异显著(图1b,P<0.05)。当年苗的Gs也呈现下降趋势,其中1代与3代,2代与3代苗木之间差异显著(图1c,P<0.05),1代与2代之间差异不显著(图1c,P>0.05)。而B模式下苗木的Gs则是先增大后减小,即该连作方式下第2代苗木的Gs最大,且与1代、3代苗木之间差异显著(图1c,P<0.05)。A模式下苗木的WUE随着连作代数的增加而升高,B模式下苗木的WUE则呈现出相反的规律。其中在A连作模式下,各代苗木之间的WUE没有显著差异(图1d,P>0.05),而B连作模式下各代之间WUE差异均显著 (图1d,P<0.05)。
2.2 两种不同连作模式下各代苗木气体交换参数对几个主要环境因子的响应

表1 光合,蒸腾,气孔导度,水分利用效率等气体交换参数对环境因子的逐步回归Table 1 Stepwise regression of photosynthesis rate(Pn),transpirationrate(E),stomatal conductance(Gs)and water use efficiency(WUE)to the environmental factors under the two continuous cropping types
苗木的气体交换过程易受到环境因子变化的影响,利用气体交换参数(Pn、E、Gs、WUE)与空气相对湿度(Rh),大气温度(Tc),光合有效辐射(Par)及大气CO2浓度(Ca)进行逐步回归分析。分析结果表明,在不同的连作模式和连作代数下,环境因子均有被剔除的现象(表1),表中系数均为标准化系数,可以表示各环境因子的影响程度。从模拟方程可以看出,不同连作模式和连作代数下苗木气体交换参数受外界环境因子的交互影响较大,且因子影响程度的差异也较大。这说明连作地上苗木的气体交换除了受连作方式和连作代数影响之外,同时受外界环境因子的交互影响,且影响程度差异较大。
根据模拟方程中的因子数量及标准化系数,两种连作模式比较,Pn在第1代、第2代时,A连作模式容易受环境因子的交互影响,且Rh的影响最大。到了第3代,则是B模式下交互影响强烈;E在第1代,A模式下的苗木对环境因子响应没有达到显著水平,而B模式受Tc变化的影响显著,到了第2代两种模式下的苗木受两种环境因子不同程度的交互影响,第3代则是A连作模式下苗木更容易受环境因子的交互影响,且Ca影响最大;对于Gs,第1代、第3代A模式下苗木易受环境因子的交互影响,而第2代时B连作模式下受影响较大。对于WUE,第1代B模式下苗木受环境因子的交互影响强烈,到第3代,A模式则更容易受到环境因子的交互影响。
2.3 两种连作模式下各代苗圃地内杨树苗木的生长状况
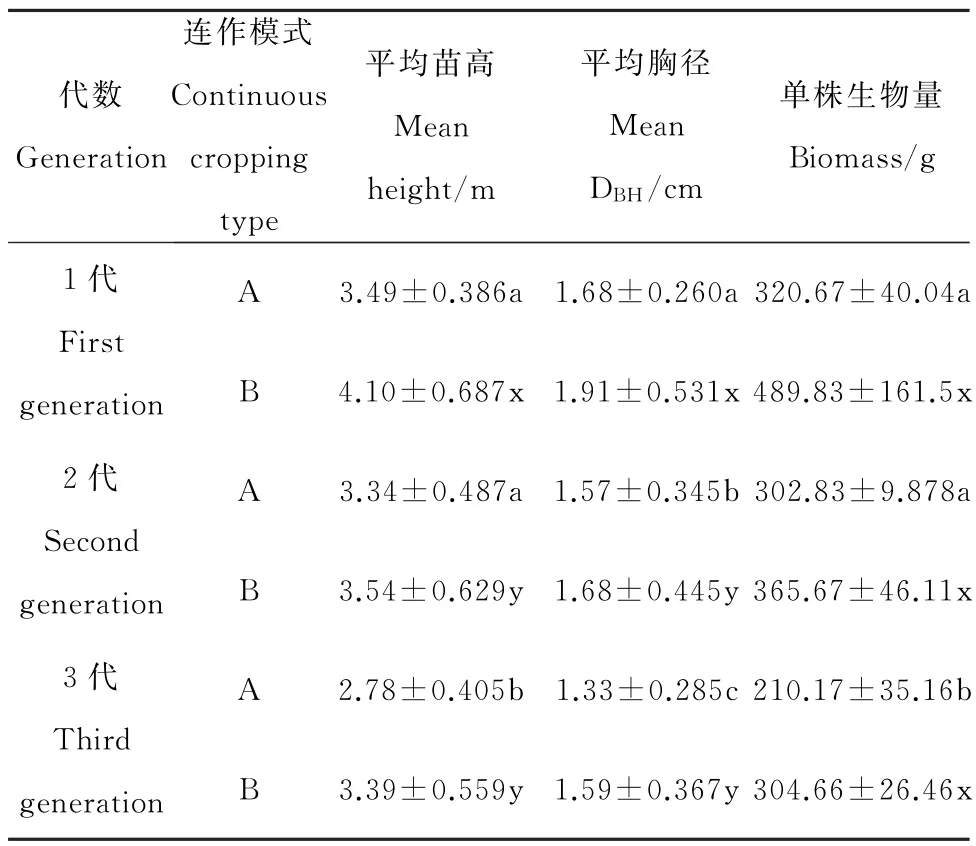
表2 两种连作模式下各代杨树苗木生长状况Table 2 The cuttings growth of different generations under two continous cropping types
苗木的气体交换过程,特别是光合作用过程是生物量累积的过程,林分生长量是生物量累积最直观的指标,通过对同一品种、同一立地条件下不同连作地杨树生长状况的调查表明,连作后苗木的苗高、胸径、单株生物量与不连作相比都有不同程度的降低。从表2可以看出,A连作模式下2代、3代的苗木与1代的苗木相比,平均胸径分别下降6.55%,20.83%;平 均 苗 高 分 别 下 降 4.30%,20.34%;单株生物量分别减少5.56%,34.46%。其中,1代与2代苗木的苗高,单株生物量没有显著性差异(表2,P>0.05),但第3代与第2代、第1代的苗高、单株生物量差异均达到显著水平(表2,P<0.05);而平均胸径在各代之间差异均达到显著水平(表2,P<0.05)。B连作模式下,2代、3代苗木与1代相比,平均胸径分别下降12.04%,16.75%;平均苗高分别下降13.66%,17.32%;单株生物量分别减少25.35%,37.80%。其中,1代苗木的平均苗高、胸径与2代、3代差异均显著(表2,P<0.05),但是第2代、第3代之间差异不显著(表2,P>0.05);平均单株生物量在1代、2代之间差异不显著(表2,P>0.05),但都与第3代差异显著(表2,P<0.05)。两种连作方式相比,B连作模式下苗木的平均苗高、胸径及单株生物量均大于A连作模式下的苗木。
2.4 两种不同连作模式下杨树苗木的叶绿素含量
叶绿素在光合作用过程中起着十分关键的作用,叶绿素a具有收集和转化光能的作用,叶绿素b具有收集光能的作用,若叶绿素a,b的含量降低,植物收集和转化光的能力下降,将影响苗木正常的光合作用过程。
从表3中看出,A连作模式下,与1代相比,2代和3代苗木的叶绿素a含量分别下降了10.38%和22.54%;叶绿素b含量分别下降了6.20%和16.30%;叶绿素总量分别下降了9.43%和21.12%;叶绿素a/b的值分别下降4.45%和7.46%。其中各代之间的叶绿素a、b含量及叶绿素总量差异均显著(表3,P<0.05),而叶绿素a/b差异不显著(表3,P>0.05)。B连作模式下,与1代相比,2代和3代杨树苗木的叶绿素a含量分别下降了30.26%、47.43%;叶绿素b的含量分别下降了31.24%、47.86%;叶绿体色素的含量分别下降了30.49%、47.53%;而叶绿素a/b先增大后减小,即2代杨树苗木的叶绿素a/b值最大。其中叶绿素a、b含量及叶绿体色素总量仅在1代与3代之间差异显著(表3,P<0.05),1代和2代、2代和3代之间差异不显著(表3,P>0.05);而叶绿素a/b在1代和3代、2代和3代之间差异显著(表3,P<0.05),1代和2代之间叶绿素a/b差异不显著(表3,P>0.05)。
两种不同的连作模式相比,叶绿素a、b含量及叶绿体色素总量均表现为A连作模式高于B连作模式;叶绿素a/b在第1代时A连作模式高于B连作模式,但第2代、第3代苗木则相反。

表3 两种连作模式下各代杨树苗木叶绿素含量Table 3 Chlorophyll of poplar cuttings in different generations under the two continuous cropping types
2.5 两种不同连作模式下杨树苗木的生物量分配
苗圃地连作影响了苗木的单株生物量,对其生物量的结构也有着直接的影响。由表4知,A连作模式下,每一代林分各部分生物量所占比例均表现为茎>叶>插穗>根,随着连作代数的增加茎生物量所占比例呈下降趋势,叶、根系生物量所占比例则有所上升,但总体上地上部分生物量所占比例是下降。有研究指出根系生物量所占比例升高与连作土壤中养分和水分供应能力下降,刺激林木强化其吸收器官从而使其所占比重增大有关[10]。而B连作模式下,随着苗圃地连作次数的增加茎生物量所占比例呈上升趋势,叶、根系及插穗生物量所占比例则有所下降。

表4 两种连作模式下杨树苗木的生物量分配及各部分所占比例Table4 The biomass of different parts in different generations under the two continuous cropping types
两种不同的苗圃地连作模式相比,各代苗木在B连作模式下茎和根的生物量明显高于A连作模式。因此在土地利用受到限制,而采取连作方式经营的苗圃地采取留根截干模式进行苗木经营往往会加大生物量向茎干部分分配的比例。从而使苗木质量提高,更增加经济收入。
3 讨论
杨树是我国主要造林树种之一,因其速生丰产、适应性强、用途广泛等特点而深受人们的欢迎。杨树在保持水土、防风固沙、保护生态环境、促进地区经济发展方面也发挥了巨大的经济、生态和社会效益[11]。近年来,由于培育管理制度及育苗技术不当,引起了苗圃地力严重衰退,特别是杨树苗圃地连续育苗,导致的苗木扦插成活率低、生长势弱、病虫害发生严重等现象,对苗圃生产和造林质量产生了严重影响。本研究对两种连作模式下不同连作代数苗圃地上苗木的生长调查证实了这种衰退现象。优质种苗生产是实现我国高效生态林业建设目标最基本的物质基础[12],因此杨树壮苗培育显得益发重要。该研究中杨树苗木的气体交换参数在不同连作模式和不同连作代数之间表现出较大的差异,同时受到外界环境因子的交互作用明显,因此在耕地面积减少,不可避免进行苗圃连作时,我们应该加强对苗圃地的管理,除了按照适地适树的原则合理布置育苗结构外,重点应放在苗圃地的管理上,如合理的连作模式,合理的灌溉、施肥[12]等措施,通过这些措施可以适当的改善连作地上的微环境,从而改善连作地苗木的气体交换和生物量累积,实现壮苗培育。
苗木的气体交换和生物量累积密切相关,本研究中不同连作模式和不同连作代数苗木的生物量累积变化较大,各部分生物量的分配比例也会发生较大的变化。两种连作模式下各部分生物量及分配比例的变化说明,留根截干方式更利于在连作苗圃地上的经营,特别是到了连作后期,留根截干模式下,苗木根、茎部分的生物量大于当年扦插模式,根是吸收养分和矿物质的主要器官,是与苗木生长密切相关的,根系生物量的增加,有利于苗木忍耐连作地上恶劣的土壤环境。而以收获干材为主的苗木生产,留根截干更是加大了生物量向茎干的分配比例,实现壮苗培育。但这并非绝对,两种模式下苗木的生长情况也与插穗的生物量及土壤状况有关,虽然两种连作方式开始时插穗的规格和土壤养分状况基本一致,但2年根1年干模式下的插穗经过了1年的生长积累了大量的营养物质有利于第2年抽条生长,加上不同连作模式下土壤环境的影响,使得两种模式下的比较变得更加复杂,需要进一步的研究证实。
[1]潘玉明,杨占英.苗圃地力问题的探讨与对策[J].青海农林科技,2001,(3):40-41
[2]刘福德,姜岳忠,刘颜泉,等.重茬对杨树苗期生长特性影响的研究[J].江西农业大学学报,2004,26(5):681-685.
[3]刘福德,姜岳忠,刘颜泉,等.连作I-107杨树无性系苗圃地的土壤酶活性特征[J].中国水土保持科学,2005,3(2):119-124.
[4]范少辉,马祥庆,陈绍栓,等.多代杉木人工林生长发育效应的研究[J].林业科学,2000,36(4):9-15.
[5]朱宇林,温远光,曹福亮,等.短周期尾巨桉连栽林分生产力的研究[J].江西农业大学学报,2006,28(1):90-94
[6]Jing Quan Yu and Yoshuhisa Matsui.Phytotoxic Substances in Root Exudutes of Cucumber[J].Journal of Chemical Ecology,1994,20(1):21-34
[7]何宗明,范少辉,卢镜铭,等.立地管理措施对2代6年生杉木林生长的影响[J],林业科学,2006,42(11):47-51.
[8]范少辉,何宗明,卢镜铭,等.立地管理措施对2代5年生杉木林生长影响[J],林业科学研究,2006,19(1):27-31.
[9]陈婷,温远光,孙永萍,等.连栽桉树人工林生物量和生产力的初步研究[J].广西林业科学,2005,34(1):8-12.
[10]Comeau PG..Above-and-bellow-ground biomass and production of lodgpole pine on sites with differing soil moisture regimes[J].Canada Journal of Forest Research.1989,19:477-454.
[11]方升佐.杨树定向培育[M].合肥:安徽科学技术出版社,2004.
[12]方升佐,田野,袁发银.扦插密度对杨树无性系苗木生长及质量的影响[J].南京林业大学学报,2004,28(4):1-5.
[13]Gert N,Ekblad A,Roland B,et al.Short-term patterns of carbon and nitrogen mineralisation in a fallow field amended with green manures from agroforestry[J].Biology and Fertility of Soil,2002,36(1):18-15.
Gas exchange and biomass distribution of poplar cuttings(I-107,Populus×Euramercana‘Neva’)under two continuous cropping types
HUANG Yanjun1,ZHU Wencheng2,AN Xiao lin2,REN Fei2,ZHANG Peng yuan3,DONG Yufeng2∗
(1.Shandong Provincial Water Resource Research Institute,Jinan 250013;2.Forestry Science Academy of Shandong,Jinan 250014)
Gas exchange and biomass distribution of I-107(Populus×Euramercana‘Neva’)poplar cutting under two continuous cropping types(A represent one year cutting;B represent two years cutting shoot and one year cutting trunk)were studied.Result showed that,under A type,photosynthesis rate(Pn),transpiration rate(E)and stomatal conductance(Gs)decreased with generation increased,but water use efficiency(WUE)increased.Under B type,PnandWUEdecreased with generation increased,butEincreased,Gsincreased firstly,then decreased.Therein,Pn、EandGsof 1stand 2ndgenerations under A type were higher than B type;As continuous generation increased,height(H),diameter at breast height(DBH)and total biomass under the two types both decreased.Therein,H,DBHand biomass of poplar cuttings under B type were higher than A type in the same generation;Under two types,the chlorophyll a(chlⅡa)content,chlorophyll b(chlⅡb)content and total chlorophyll(chlⅡ)content decreased with generation increased.Therein,the three parameters under A type were higher than B type;As the generation increased,root biomass and stem biomass of poplar cuttings under A and B types both decreased,but variation of the ratio of each part biomass under the two types showed the reverse trend.Therein,root biomass and stem biomass of poplar cuttings under B type were higher than A type.
generation;poplar;cutting;gas exchange;biomass distribution
S792.11
A
1002-2724(2016)05-0016-07
2016-09-03
国家重点专项“杨树工业资源材高效培育技术研究”(2016YFD0600400),十二五科技支撑“阔叶树速生丰产林定向培养技术”(2015BAD09B02),山东省中青年科学家奖励基金(BS2015NY013)。
∗通讯作者。
