皂荚种质资源RSAP遗传多样性分析及指纹图谱的构建
2017-12-20范定臣张安世
范定臣,张安世,刘 莹,骆 扬
(1.河南省林业科学研究院,河南 郑州 450008; 2.焦作师范高等专科学校 理工学院,河南 焦作 454000)
皂荚种质资源RSAP遗传多样性分析及指纹图谱的构建
范定臣1,张安世2*,刘 莹2,骆 扬2
(1.河南省林业科学研究院,河南 郑州 450008; 2.焦作师范高等专科学校 理工学院,河南 焦作 454000)
利用RSAP标记技术对18份皂荚种质材料进行了遗传多样性分析。结果表明,从45对RSAP引物中筛选了12对引物进行PCR扩增,共扩增出167个条带,其中多态性条带154个,多态性位点百分率(PPL)为92.22%。各引物Nei’s基因多样性指数(H)和Shannon’s信息指数(I)的平均值分别为0.229 8和0.371 6,18份皂荚种质的遗传相似系数(GS)为0.431 1~0.988 0。 UPGMA聚类分析表明,在GS为0.62处可将18份皂荚种质资源分为3组,其中,野皂荚单独为一组,山皂荚和皂荚-T聚为一组,其他皂荚材料聚为一组。利用6对引物扩增的9个多态性位点,构建了18份皂荚种质资源的DNA指纹图谱,可以将其区分并精准鉴定。
皂荚; RSAP; DNA指纹图谱; 遗传多样性
皂荚(GlediysiasinensisLam.)又叫皂角,属豆科苏木亚科皂荚属(GleditsiaLinn.)植物,是我国重要的乡土树种,广泛分布于我国东北、华北、华东、华南等地,多生长于平原、山谷及丘陵地区,具有悠久的栽培历史。皂荚适应性广[1],抗性强,能抗旱、耐盐、耐高温[2]。皂荚具有多种用途,皂荚、皂刺、皂根、皂叶均可入药,荚果富含皂甙,是重要的化工原料[3],具有很高的药用价值和工业价值[4],杀虫、杀鼠效果也非常好[5-6]。皂荚又是优良的生态树种,具有多种生态属性,广泛用作经济林、用材林、防护林及园林绿化[7]。目前,国内外对皂荚的研究主要集中在皂荚果实与皂刺中活性成分[1,8-9]、引种栽培[10]、生态属性[11]等方面,有关遗传学[12-14]方面的研究还仅限于表型分析。最近,分子标记技术已开始在皂荚研究中应用。邢俊连等[15]利用已公布的皂荚转录组数据设计得到6 494条EST-SSR特异性引物,对6个来源地的12份皂荚材料进行了有效性筛选和多态性分析,并利用筛选出的有效引物对皂荚9个近缘种进行了引物通用性检测,结果证实,这些引物在其近缘种间具有很高的通用性,为以后开展皂荚种质资源的遗传多样性研究提供了有效的分析方法和手段。李伟等[16]利用AFLP分子标记技术对10个南方皂荚群体进行了遗传多样性研究,具体分析了皂荚群体遗传结构形成的原因,并提出了皂荚的保护策略。目前,尚未见到有关皂荚RSAP分子标记研究的报道。
限制性位点扩增多态性(Restriction site amplified polymorphism,RSAP)是由杜晓华等[17]基于基因组上广泛分布的限制性酶切位点而建立的一种新型分子标记技术。RSAP在PCR扩增程序上参照了SRAP标记的引物设计,借鉴了RAPD标记随机序列的模糊匹配原理,在操作上又避免了AFLP的限制酶切等复杂步骤,具有操作简单、稳定可靠、产率中等特点,已在辣椒[18-19]、南瓜[20]、大花三色堇[21]、龙须菜[22]等植物研究上得到了成功应用。本试验采用RSAP标记对河南省18份皂荚种质资源进行遗传多样性分析,并构建DNA指纹图谱,以期为皂荚种质资源的鉴定、杂交育种亲本的选配等提供科学依据。
1 材料和方法
1.1 材料
供试皂荚材料包括3个种共18份,具体见表1。其中野皂荚、山皂荚和皂荚-T为实生苗,其余均为嫁接苗。
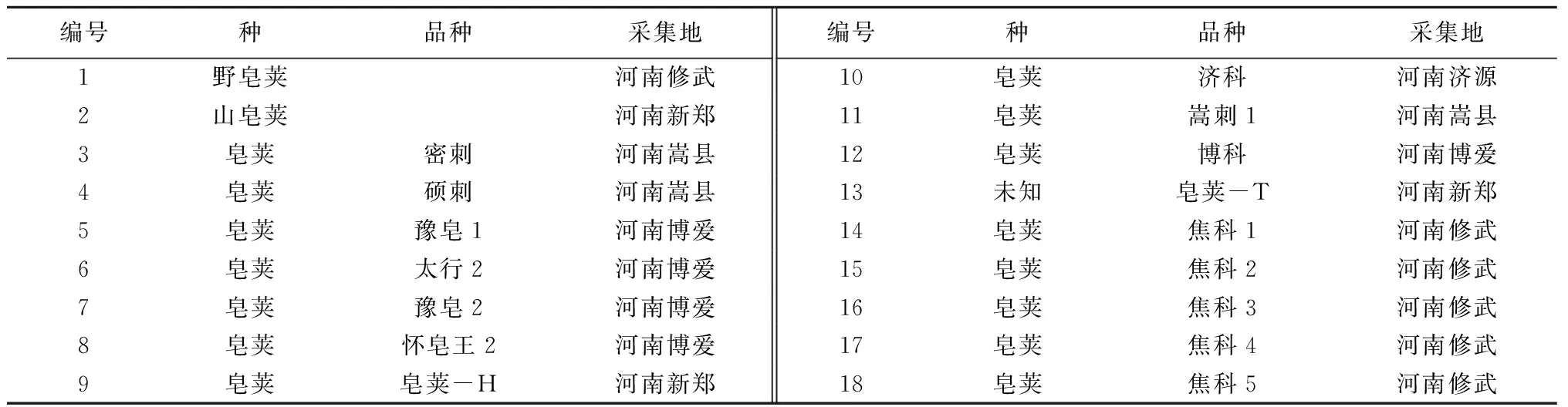
表1 供试材料
1.2 皂荚基因组DNA的提取
采用改良CTAB法[23]提取皂荚基因组DNA,并将模板DNA质量浓度稀释至20 ng/μL,保存于-20 ℃备用。
1.3 RSAP-PCR分析
选用10个RSAP引物(Rs1—Rs10)两两组合共45对RSAP引物对供试材料进行扩增。反应体积为10 μL,包括DNA 1.0 μL、正反引物各 0.5 μL、2×TaqMasterMix 5.0 μL、RNase-Free water 3.0 μL。RSAP-PCR扩增程序为:94 ℃ 5 min;94 ℃ 1 min,35 ℃ 1 min,72 ℃ 1.5 min,5个循环;94 ℃ 1 min,52 ℃ 1 min,72 ℃ 1.5 min,35个循环;72 ℃ 7 min。4 ℃保存。扩增产物用1.5%琼脂糖凝胶分离。
1.4 数据统计与分析
根据电泳结果进行条带的统计分析。电泳图在相同位置上若出现DNA条带记为“1”,无则记为“0”,形成1,0矩阵,将此矩阵导入POPGENE 1.32软件进行多态性位点百分率(PPL)、Nei’s基因多样性指数(H)和Shannon’s信息指数(I)等遗传多样性参数分析,最后通过NTSYS-pc 2.0软件依据UPGMA法进行聚类分析。同时,参照云天海等[24]的方法构建18份皂荚种质材料的DNA指纹图谱。
2 结果与分析
2.1 RSAP引物筛选与多态性分析
以供试的18份皂荚材料对45对RSAP引物进行了筛选,最终筛选出了条带清晰、多态性高的12对引物,所用引物序列及多态性统计结果见表2。12对引物共扩增出167个条带,多态性条带154个,PPL为92.22%,说明皂荚具有较高的RSAP多态性。在被统计的12对引物中,引物组合Rs7-Rs10扩增的位点数最多,为24个,引物组合Rs1-Rs6、Rs1-Rs7、Rs5-Rs10和Rs7-Rs9扩增的位点数最少,为10个,平均每对引物扩增的位点数为13.92个。引物组合Rs6-Rs10的扩增结果见图1。通过POPGENE 1.32软件分析得到皂荚遗传多样性参数(表2),各引物Nei’s基因多样性指数(H)的变化范围为0.148 6~0.302 8,平均值为0.229 8,H值最高的引物组合为Rs2-Rs9(0.302 8),H值最低的引物组合为Rs1-Rs8(0.148 6);各引物Shannon’s信息指数(I)的变化范围为0.256 7~0.461 7,平均值为0.371 6,I值最高的引物组合为Rs2-Rs9 (0.461 7),I值最低的引物组合为Rs1-Rs8(0.256 7)。

表2 RSAP引物及多态性分析
注:T为位点总数; N为多态性位点数。

M:标准分子质量; 1~18:材料编号同表1,下同
2.2 皂荚种质资源间的遗传相似性和聚类分析
利用NTSYS-pc软件计算皂荚种质材料间的遗传相似系数(GS),结果表明,18份皂荚种质材料间的GS为0.431 1~0.988 0,平均值为0.756 6,变幅为0.556 9,说明供试材料间存在较大的遗传差异。其中焦科4和焦科5的GS最大(0.988 0),亲缘关系最近,豫皂1和皂荚-T的GS最小(0.431 1),亲缘关系最远。
利用NTSYS-pc软件对18份皂荚种质材料进行聚类分析(图2),结果表明,在GS为0.62处可将18份皂荚种质材料分为3组。第1组为野皂荚;第2组为山皂荚和皂荚-T;第3组共有15份皂荚材料:密刺、豫皂2、怀皂王2、博科、皂荚-H、硕刺、焦科1、焦科3、豫皂1、太行2、济科、嵩刺1、焦科2、焦科4和焦科5。该聚类结果与皂荚的传统分类基本一致。
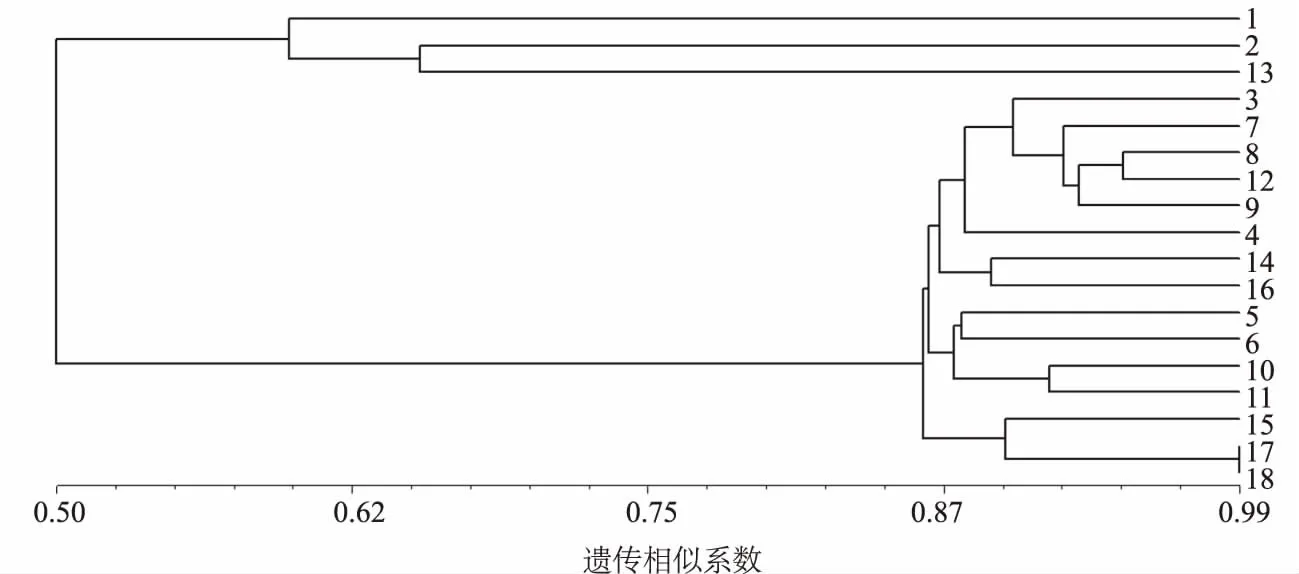
图2 18份皂荚种质资源的聚类分析结果
2.3 皂荚种质资源DNA指纹图谱的构建
通过分析筛选的12对引物对18份皂荚种质材料的扩增结果,选取其中Rs2-Rs9、Rs4-Rs7、Rs4-Rs8、Rs5-Rs10、Rs6-Rs10和Rs7-Rs9六对引物扩增的9个多态性位点构建了18份皂荚种质的DNA指纹图谱(图3)。每份材料都有唯一的指纹图谱,可以将18份皂荚种质材料区分并准确鉴定。

图3 18份皂荚种质资源的DNA指纹图谱
3 结论与讨论
3.1 皂荚种质资源的遗传多样性分析
与形态学标记相比,分子标记能直接反映基因组DNA的遗传变异信息,有助于了解种质资源之间的遗传差异,对于种质资源的合理利用具有重要意义。在本研究中,利用RSAP标记就DNA扩增的总位点数、多态性位点数、多态性位点百分率、Nei’s 基因多样性指数、 Shannon’s 信息指数等多项指标进行了分析。结果表明,12对引物共扩增出167条带,其中多态性条带154条,多态性位点百分率高达92.22%,Nei’s 基因多样性指数、Shannon’s 信息指数分别为0.229 8和0.371 6,说明18份皂荚种质资源具有较高的RSAP多态性,遗传变异也较为丰富。上述研究结果可以为皂荚优良品种的选育和皂荚种质资源的鉴定提供理论指导。
3.2 皂荚种质资源的聚类分析
本研究涉及野皂荚、山皂荚和皂荚3个种共18份材料。聚类结果表明,在GS为0.62处可将18份皂荚种质材料分为3组。其中,野皂荚单独为一组,山皂荚和皂荚-T聚为一组,其余15份皂荚嫁接苗聚为一组。从整体结果来看,该聚类结果与皂荚的传统分类基本吻合。在上述第3组的15份皂荚种质材料中,焦科4和焦科5是以皂荚荚果为目的选育的品种,其余均以皂刺为目的选育。焦科4和焦科5的接穗均为荚果较大的不同皂荚品种,聚类结果显示,两者在所有供试材料中亲缘关系最近,与预期相符。济科是以嵩刺1为接穗、山皂荚为砧木的嫁接品种,两者首先聚在一起,具有很近的亲缘关系,也与实际情况相吻合。另外,皂荚-T是从土耳其引进种子的3年生播种苗,起初曾被作为皂荚品种,但其3年生播种苗无论从叶的形态、皮孔大小及形状以及嫩枝的颜色都与山皂荚极为相似,因此认为该种子在引进时可能鉴定有误。本研究结果显示,山皂荚和皂荚-T聚为一组,两者具有很近的亲缘关系,因此,应将皂荚-T归为山皂荚而非皂荚品种,同时还要结合其他分子标记技术及其以后的生长过程中所表现出的其他形态学特征进行进一步的验证。
3.3 RSAP标记技术在种质资源鉴定上的应用分析
分子标记以其高效、快捷、准确度高、信息量大、适合数字化等优点,已经成为植物种质鉴定的有效方法。乔利仙等[25]利用RSAP标记技术从扩增的408条多态性条带中选取其中10条稳定且重复性好的条带构建了15个紫菜系的DNA 指纹图谱,并将此图谱实现了计算机化,可以验证这些紫菜系的身份。刘泽发等[20]利用SRAP和RSAP 2种标记方法,对印度南瓜品种杂交种子进行了纯度鉴定试验,选用引物 E1M5 和 R1R7能够较好地鉴定红栗二号、红栗、锦栗二号和锦栗南瓜,并将引物R1R7检测的锦栗杂交种子纯度与田间检测结果进行了对比,其吻合率达到 90%。本研究利用RSAP技术,选用6对引物扩增的9个多态性位点构建了18份皂荚种质材料的DNA指纹图谱,为皂荚种质资源的鉴别提供了重要的科学依据。
[1] 张宏利,简利茹,韩崇选,等.皂荚果实中皂苷O的分离鉴定及毒性研究[J].西北植物学报,2013,33(6):1234-1238.
[2] 姚永胜,马秀琴.银川地区皂角引种及造林试验[J].宁夏农林科技,1998(6):34-35.
[3] 杨洋,菅红磊,徐永霞,等.皂荚多糖胶酶解制备低聚糖[J].食品科学,2011,32 (18):138-141.
[4] 朱莉伟,蒋建新.皂荚豆和日本皂荚豆氨基酸及脂肪酸组成分析[J].中国野生植物资源,2001,20(4):40-41.
[5] 邵则夏,陆斌,杨卫明,等.多功能树种滇皂荚及开发利用[J].中国野生植物资源,2002,21(3):33-34.
[6] 张一贞,韩崇选,张宏利,等.两种皂荚提取物的杀鼠活性研究[J].西北林学院学报,2007,22(1):106-108.
[7] 顾万春,兰彦平,孙翠玲.世界皂荚(属)的研究与开发利用[J].林业科学,2003,39(4):127-133.
[8] 林雪娇,周自圆,菅红磊,等.皂荚半乳甘露聚糖胶分级醇沉及表征[J].中国野生植物资源,2013,32(4):20-23.
[9] 李建军,尚星晨,任美玲,等.皂荚实生苗与嫁接苗皂刺单株质量及药用成分含量比较[J].河南农业科学,2017,46(8):107-110.
[10] 范定臣,马群智,杨伟,等.中原地区皂荚栽培技术[M].郑州:黄河水利出版社,2015.
[11] 杨海东.皂荚的多种功效及其绿化应用[J].贵州农业科学,2003,31(4):73-74.
[12] 李伟,林富荣,郑勇奇,等.皂荚南方天然群体种实表型多样性[J].植物生态学报,2013,37(1):61-69.
[13] 李伟,林富荣,郑勇奇,等.皂荚天然群体间种实表型特性及种子萌发的差异分析[J].植物资源与环境学报,2013,22(4):70-75.
[14] 兰彦平,顾万春.北方地区皂荚种子及荚果形态特征的地理变异[J].林业科学,2006,42(7):47-51.
[15] 邢俊连,孟艳琼,林富荣,等.皂荚EST-SSR分子标记开发与分析评价[J].植物遗传资源学报,2017,18(1):149-155.
[16] 李伟,林富荣,郑勇奇,等.10个南方皂荚群体遗传多样性的AFLP分析[J].林业科学研究,2017,30(1):46-52.
[17] 杜晓华,王得元,巩振辉.一种新型 DNA标记技术——限制性位点扩增多态性(RSAP)的建立与优化[J].西北农林科技大学学报(自然科学版),2006,34(6):45-49,54.
[18] 杜晓华,王得元,巩振辉.基于 RSAP和SSR 的辣椒优良自交系间遗传距离的估计与比较[J].西北农林科技大学学报(自然科学版),2007,35(7):97-102.
[19] 左泽彦,吴拥军,罗熹,等.贵州省加工型辣椒资源RSAP分析[J].河南农业科学,2012,41(7):116-119.
[20] 刘泽发,孙小武,董亚静,等.SRAP/RSAP 标记鉴定印度南瓜种子纯度的方法[J].西北农业学报,2011,20(4):124-128.
[21] 李小梅,杜晓华,穆金燕,等.基于RSAP标记的大花三色堇遗传多样性分析[J].西北植物学报,2015,35(10):1989-1997.
[22] 王津果,隋正红,周伟,等.龙须菜RSAP分析及其SCAR标记的转化[J].中国海洋大学学报,2014,44(4):47-53.
[23] 张安世,邢智峰,刘永英,等.苔藓植物DNA不同提取方法的比较分析[J].河南科学,2009,27(5):559-562.
[24] 云天海,郑道君,谢良商,等.中国南瓜海南农家品种间的遗传特异性分析和 DNA 指纹图谱构建[J].植物遗传资源学报,2013,14(4):679-685.
[25] 乔利仙,翁曼丽,孔凡娜,等.RSAP 标记技术在紫菜遗传多样性检测及种质鉴定中的应用[J].中国海洋大学学报,2007,37(6):951-956.
Genetic Diversity and Fingerprints of Glediysia sinensis Germplasms Based on RSAP
FAN Dingchen1,ZHANG Anshi2*,LIU Ying2,LUO Yang2
(1.Henan Academy of Forestry,Zhengzhou 450008,China; 2.School of Science,Jiaozuo Teachers College,Jiaozuo 454000,China)
Genetic diversity of 18Glediysiasinensisgermplasms was analyzed by the RSAP markers.The results showed that 12 primers were screened out of 45 primer pairs and 167 bands were obtained,including 154 polymorphic bands,with the percentage of polymorphic loci(PPL) of 92.22%.The average Nei’s gene diversity(H) and Shannon’s information index(I) were 0.229 8 and 0.371 6,and the genetic similarity coefficients among the tested samples ranged from 0.431 1 to 0.988 0.UPGMA analysis showed that 18Glediysiasinensiswere clustered into 3 groups with the genetic similarity coefficient of 0.62.Glediysiaheterophyllaformed the first group,Glediysiamelanacanthaand Zaojia-T were clustered into the second group,and the others were clustered into the third group.In total of 18Glediysiasinensisgermplasm DNA fingerprints were constructed from 9 polymorphic loci amplified by 6 primer pairs and these materials could be distinguished and identified accurately.
Glediysiasinensis; RSAP; DNA fingerprint; genetic diversity
S718.46
A
1004-3268(2017)11-0103-05
2017-06-20
河南省科技攻关项目(14210221100850);2015年河南省林业厅科技兴林项目
范定臣(1967-),男,河南沁阳人,高级工程师,本科,主要从事园林绿化及林木育种研究。
E-mail:fdclky@139.com
*
张安世(1965-),男,河南博爱人,教授,硕士,主要从事植物分子生物学研究。E-mail:aszhang1212@163.com
