中原地区汉族和回族中国人群CYP2C9*3、VKORC1-1639G>A和CYP4F2*3的遗传多态性
2017-12-19齐光照王维杰郑州大学第一附属医院药学部郑州45005黄河科技学院医学院郑州大学第一附属医院肝胆胰外科通讯作者mailcyp4509com
齐光照,薄 惠,王维杰(郑州大学第一附属医院药学部,郑州 45005;黄河科技学院医学院;郑州大学第一附属医院肝胆胰外科;通讯作者,E-mail:cyp4509@6.com)
中原地区汉族和回族中国人群CYP2C9*3、VKORC1-1639G>A和CYP4F2*3的遗传多态性
齐光照1,薄 惠2,王维杰3*
(1郑州大学第一附属医院药学部,郑州 450052;2黄河科技学院医学院;3郑州大学第一附属医院肝胆胰外科;*通讯作者,E-mail:cyp450239@126.com)
目的 探讨参与华法林药物反应的功能性变异体CYP2C9*3、VKORC1-1639G>A和CYP4F2*3在中原地区汉族和回族中国人群的遗传多态性。 方法 提取285例无血缘关系的健康中国志愿者(包括175例汉族和110例回族)外周血DNA进行基因型分析。CYP2C9*3和VKORC1-1639G>A的基因型频率通过多聚酶链反应-限制性片段长度多态性的方法检测,而CYP4F2*3的基因型通过多聚酶链反应-Sanger测序的方法检测。将中原地区汉族和回族CYP2C9*3、VKORC1-1639G>A和CYP4F2*3的等位基因频率与文献已报道的高加索人和非洲人频率进行比较。 结果 汉族和回族的CYP2C9*3等位基因频率分别为3.7%和5.5%。回族CYP2C9*3等位基因频率显著高于已报道的非裔美国人频率(2.0%,P<0.05)。汉族和回族VKORC1-1639A等位基因频率分别为90.3%和87.3%。汉族和回族VKORC1-1639A等位基因频率都显著高于已报道的非裔美国人频率(10.8%,P<0.05)和高加索人频率(40.6%,P<0.05)。汉族和回族CYP4F2*3等位基因频率分别为26.3%和22.3%。汉族和回族CYP4F2*3等位基因频率都显著高于已报道的非裔美国人频率(11.7%,P<0.05),但显著低于已报道的高加索人频率(34.2%,P<0.05)。 结论 中原地区汉族和回族中国人群具有相似的CYP2C9*3、VKORC1-1639G>A和CYP4F2*3的遗传多态性频率,但与文献已报道的非洲人和高加索人频率存在显著差异。
CYP2C9; CYP4F2; VKORC1; 回族; 汉族; 遗传多态性
香豆素类抗凝剂如华法林广泛应用于肺栓塞、深静脉血栓形成、心脏瓣膜手术、中风和房颤等患者[1]。但是华法林治疗窗较窄,在其抗凝治疗期间常发生严重的药物不良反应如出血,因此为了治疗的安全性和有效性确定一个合适的华法林维持剂量显得尤为重要。细胞色素酶P450 2C9(cytochrome P450 2C9,CYP2C9)以及维生素K环氧化物还原酶复合体亚基1(vitamin K epoxide reductase complex subunit 1,VKORC1)和CYP4F2参与华法林的药效学和药动学过程(见图1),多项候选基因研究[2-4]和全基因组关联研究[5-9]发现这3个基因的遗传多态性与华法林治疗反应的个体间和群体间差异相关。回族是我国人口较多的一个少数民族[10],2010年第六次人口普查全国有10 586 087人,其中河南省有957 964人。本研究考察了中原地区汉族和回族中国人群CYP2C9*3、VKORC1-1639G>A和CYP4F2*3的遗传多态性,并与已报道的其他世界人群进行了比较,现将结果报道如下。
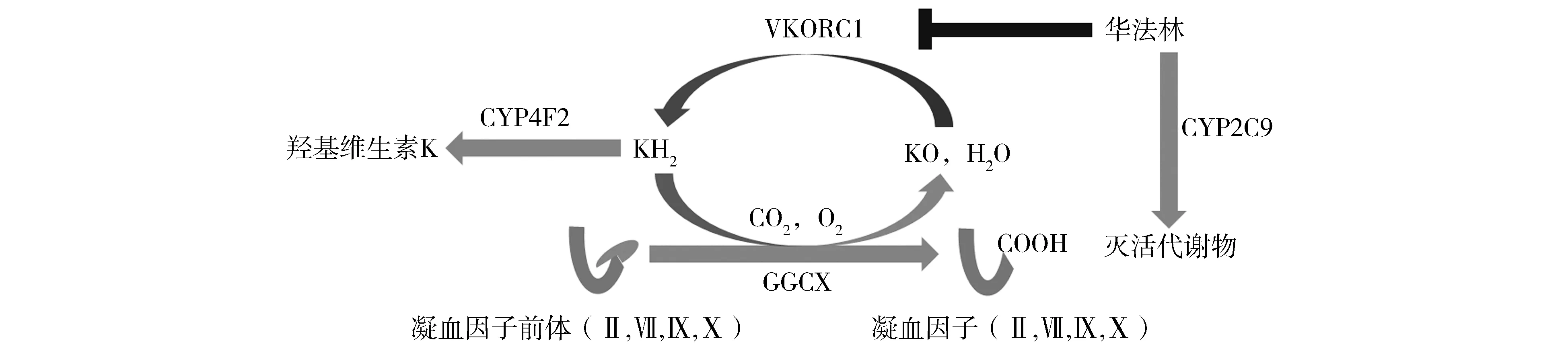
CYP2C9是华法林的代谢酶,催化华法林代谢为无活性代谢产物;VKORC1是华法林的药物作用靶点,催化环氧型维生素K(KO)还原为氢醌型维生素K(KH2);CYP4F2是氢醌型维生素K的代谢酶,催化氢醌型维生素K代谢为羟基化维生素K图1 华法林作用通路中CYP2C9、VKORC1和CYP4F2的作用Figure 1 Role of CYP2C9, VKORC1 and CYP4F2 in the pharmacokinetics and pharmacodynamics procedure of S-warfarin
1 材料与方法
1.1 研究对象
收集285例无血缘关系的健康中国志愿者(包括175例汉族和110例回族)外周血DNA进行基因型分析。每个受试者至少三代都是同一民族,即其父母及祖父母和外祖父母皆为同一民族。本研究符合《赫尔辛基宣言》相关伦理规定并得到郑州大学第一附属医院伦理委员会的批准。受试者均签署书面知情同意书。
1.2 基因型分析
利用DNA提取试剂盒QIAamp DNA Blood Mini Kits(Qiagen Inc.,Valencia,CA,USA)从乙二胺四乙酸(EDTA)处理的外周血提取基因组DNA。CYP2C9*3和VKORC1-1639G>A的基因型通过多聚酶链反应-限制性片段长度多态性的方法确定[11-14],而CYP4F2*3的基因型通过多聚酶链反应-Sanger测序的方法确定[15]。PCR条件及基因型分析的相关信息见表1。CYP2C9*3和VKORC1-1639G>A基因型分析的电泳图分别见图2和图3,CYP4F2*3的测序峰图见图4。
表1PCR扩增引物和基因型分析条件
Table1SelectedprimersandconditionsforPCRamplificationandgenotypeanalysisofCYP2C9*3,VKORC1-1639G>AandCYP4F2*3
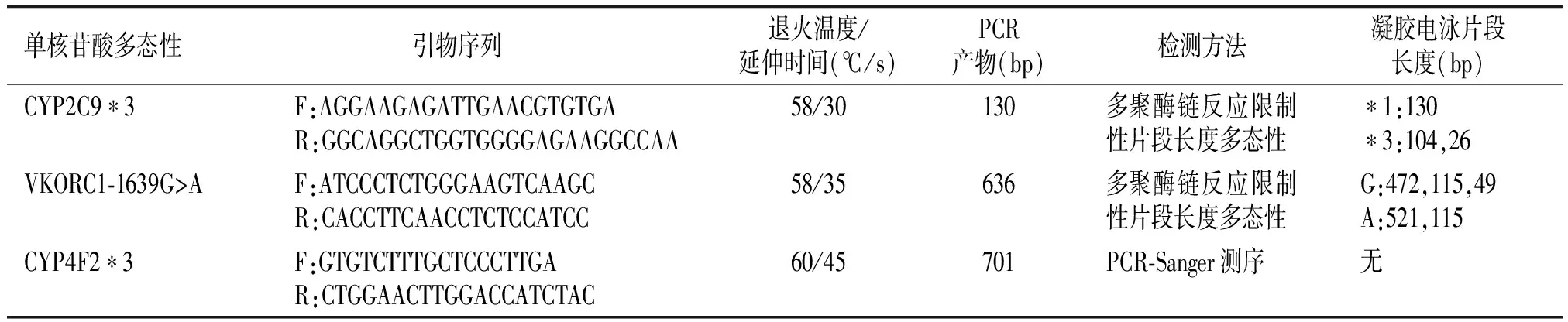
单核苷酸多态性引物序列 退火温度/延伸时间(℃/s)PCR产物(bp)检测方法凝胶电泳片段长度(bp)CYP2C9∗3F:AGGAAGAGATTGAACGTGTGAR:GGCAGGCTGGTGGGGAGAAGGCCAA58/30130多聚酶链反应限制性片段长度多态性∗1:130∗3:104,26VKORC1⁃1639G>AF:ATCCCTCTGGGAAGTCAAGCR:CACCTTCAACCTCTCCATCC58/35636多聚酶链反应限制性片段长度多态性G:472,115,49A:521,115CYP4F2∗3F:GTGTCTTTGCTCCCTTGAR:CTGGAACTTGGACCATCTAC60/45701PCR⁃Sanger测序无
F:上游引物;R:下游引物
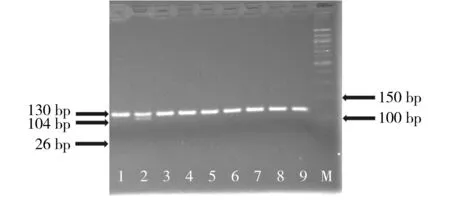
M. 50 bp DNA分子量标准;1,3-9.野生型纯合子CYP2C9*1/*1;2.杂合子CYP2C9*1/*3图2 多聚酶链反应-限制性片段长度多态性的方法对CYP2C9*3基因型分析的电泳图Figure 2 Electrophoresis of CYP2C9*3 genotyping by polymerase chain reaction-restriction fragment length polymorphism analysis(3% agarose)
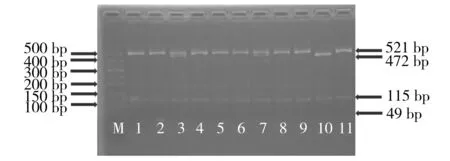
M.50 bp DNA分子量标准;10.野生型纯合子VKORC1-1639GG;1,2,4-6,8,9,11.突变纯合子VKORC1-1639AA;3,7.杂合子VKORC1-1639GA图3 多聚酶链反应-限制性片段长度多态性的方法对VKORC1-1639G>A基因型分析电泳图Figure 3 Electrophoresis of VKORC1-1639G>A genotyping by polymerase chain reaction-restriction fragment length polymorphism analysis (3% agarose)
1.3 单核苷酸多态性数据库等位基因频率资料的收集
从两个数据库(国际HapMap工程和千人基因组工程)收集了CYP2C9*3、VKORC1-1639G>A和CYP4F2*3等位基因频率的资料。国际HapMap工程包括12个人群:欧裔美国人(CEU)、北京汉族中国人(CHB)、无血缘关系汉族中国人(HCB)、美国华裔(CHD)、日本人(JPT)、非裔美国人(ASW)、印第安人(GIH)、肯尼亚非洲人(LWK)、墨西哥人(MEX)、肯尼亚马赛人(MKK)、意大利人(TSI)、尼日利亚人(YRI)。千人基因组工程包括5个人群:非洲人(AFR)、美洲人(AMR)、东亚人(EAS)、欧洲人(EUR)和南亚人(SAS)。
1.4 统计学分析
基因型的Hardy-Weinberg平衡检验采用HAP-LOVIEW 4.1(http://broad.mit.edu/mpg/haploview)软件,利用Fisher精确检验比较不同人群的基因型和等位基因频率。利用R相关软件包进行主成分分析(PCA)以分析人群之间的关系。相关统计分析在R中进行。P<0.05为差异有统计学意义。
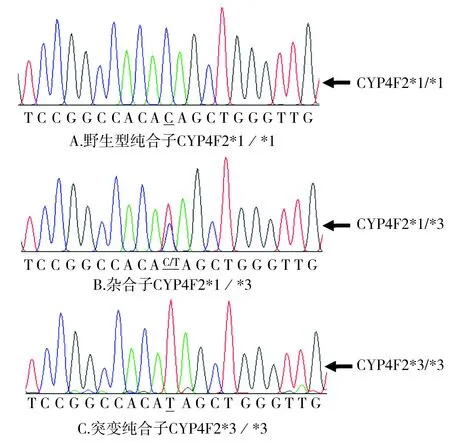
图4 CYP4F2第11外显子碱基17981-18001共21个碱基的测序峰图Figure 4 Sequencing graph of nucleotides 17981 to 18001 in exon 11 of CYP4F2
2 结果
本研究汉族和回族中国人群CYP2C9*3、VKORC1-1639G>A和CYP4F2*3的基因型频率符合Hardy-Weinberg平衡(P>0.05,见表2),说明其为遗传平衡群体。汉族和回族中国人群CYP2C9*3、VKORC1-1639G>A和CYP4F2*3的基因型及等位基因频率与已报道非裔美国人和高加索人的比较[16]见表2。
2.1 汉族和回族CYP2C9*3频率及与已报道世界其他人群的比较
CYP2C9*3突变纯合子和变异等位基因频率在汉族(0.6%和3.7%)、回族(0.9%和5.5%)和整体中国人(合并本研究汉族和回族所得,0.7%和4.4%)无显著差异(P>0.05)。但是回族CYP2C9*3/*3基因型频率显著高于非裔美国人(0,P<0.05),并且回族和整体中国人CYP2C9*3等位基因频率皆显著高于非裔美国人(2.0%,P<0.05)。这些结果说明对于中国人检测CYP2C9*3的基因与汉族比较,*P<0.05;与回族比较,#P<0.05;与整体中国人比较,&P<0.05;整体中国人为合并本研究汉族和回族所得;HWE:Hardy-Weinberg平衡型更重要,而其他变异体如CYP2C9*2在高加索人频率较高而在东亚人中比较罕见。在世界各个人群中[16-25],罗姆人(吉普赛人)CYP2C9*3的等位基因频率最高(15.5%),而美洲土著和尼日利亚人最低(皆为0)。整体来看,CYP2C9*3等位基因频率在人群的分布如下:高加索人(5.7%)>中国人(4.4%)>非裔美国人(2.0%)(见表3)。
表2本研究基因多态性频率及其与已报道非裔美国人和高加索人的比较%(例)
Table2ComparisonofgeneticpolymorphismsbetweenthepresentstudyandthosereportedpreviouslyinAfrican-AmericanandCaucasian%(cases)
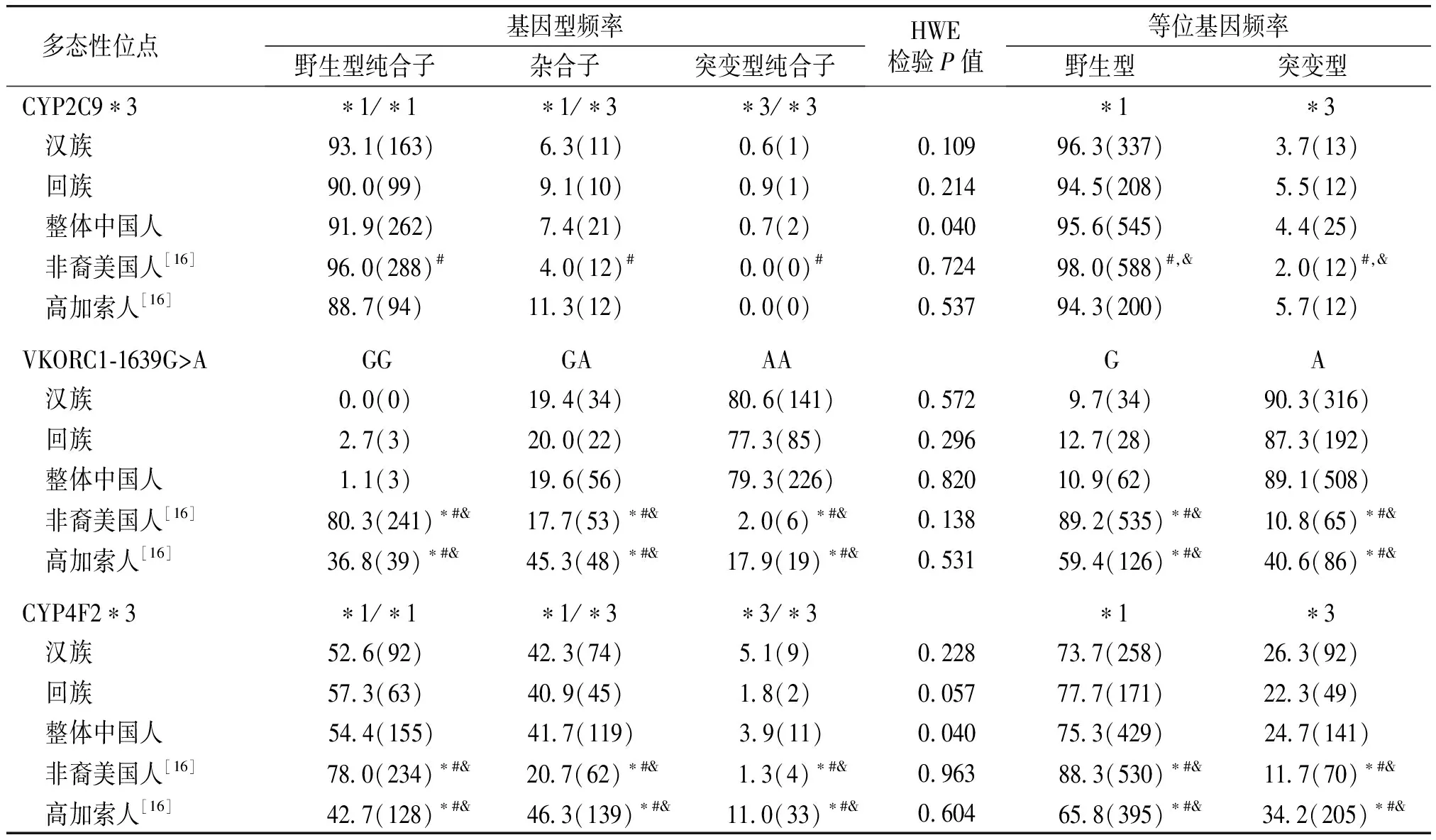
多态性位点 基因型频率野生型纯合子杂合子突变型纯合子HWE检验P值等位基因频率野生型突变型CYP2C9∗3∗1/∗1∗1/∗3∗3/∗3∗1∗3 汉族 93 1(163) 6 3(11)0 6(1)0 10996 3(337)3 7(13) 回族90 0(99) 9 1(10)0 9(1)0 21494 5(208)5 5(12) 整体中国人 91 9(262) 7 4(21)0 7(2)0 04095 6(545)4 4(25) 非裔美国人[16] 96 0(288)# 4 0(12)# 0 0(0)#0 724 98 0(588)#,& 2 0(12)#,& 高加索人[16]88 7(94)11 3(12)0 0(0)0 53794 3(200)5 7(12)VKORC1⁃1639G>AGGGAAAGA 汉族0 0(0)19 4(34) 80 6(141)0 5729 7(34)90 3(316) 回族2 7(3)20 0(22)77 3(85)0 29612 7(28) 87 3(192) 整体中国人1 1(3)19 6(56) 79 3(226)0 82010 9(62) 89 1(508) 非裔美国人[16] 80 3(241)∗#& 17 7(53)∗#& 2 0(6)∗#&0 138 89 2(535)∗#& 10 8(65)∗#& 高加索人[16] 36 8(39)∗#& 45 3(48)∗#& 17 9(19)∗#&0 531 59 4(126)∗#& 40 6(86)∗#&CYP4F2∗3∗1/∗1∗1/∗3∗3/∗3∗1∗3 汉族52 6(92)42 3(74)5 1(9)0 22873 7(258)26 3(92) 回族57 3(63)40 9(45)1 8(2)0 05777 7(171)22 3(49) 整体中国人 54 4(155) 41 7(119) 3 9(11)0 04075 3(429)24 7(141) 非裔美国人[16] 78 0(234)∗#& 20 7(62)∗#& 1 3(4)∗#&0 963 88 3(530)∗#& 11 7(70)∗#& 高加索人[16] 42 7(128)∗#& 46 3(139)∗#& 11 0(33)∗#&0 604 65 8(395)∗#& 34 2(205)∗#&
2.2 汉族和回族VKORC1-1639G>A频率及与已报道世界其他人群的比较
VKORC1-1639G>A突变纯合子和变异等位基因频率在汉族(80.6%和90.3%)、回族(77.3%和87.3%)和整体中国人(79.3%和89.1%)之间无显著差异(P>0.05)。但是非裔美国人(2.0%和10.8%)和高加索人(17.9%和40.6%)VKORC1-1639G>A突变纯合子和变异等位基因频率皆显著低于汉族、回族和整体中国人(P<0.05)。在世界各个人群中[16,18-26],CHB的VKORC1-1639A等位基因频率最高(95.1%),而YRI最低(2.2%)。整体来看,VKORC1-1639A等位基因频率在人群的分布如下:中国人(89.1%)>高加索人(40.6%)>非裔美国人(10.8%)(见表4)。
2.3 汉族和回族CYP4F2*3频率及与已报道世界其他人群的比较
CYP4F2*3突变纯合子频率在汉族(5.1%)和回族(1.8%)之间无显著差异(P>0.05)。CYP4F2*3等位基因频率在汉族(26.3%)和回族(22.3%)之间亦无显著差异(P>0.05)。但是汉族、回族和整体中国人CYP4F2*3突变纯合子和变异等位基因频率显著高于非裔美国人(1.3%和11.7%,P<0.05),但是显著低于高加索人(11.0%和34.2%,P<0.05)。河南省目前进行华法林基因检测的医院主要检测CYP2C9和VKORC1,本研究结果显示汉族和回族具有较高的CYP4F2*3频率,因此将CYP4F2*3纳入华法林基因检测也是必要的。在世界各个人群中[16,19-25,27],GIH的CYP4F2*3等位基因频率最高(43.2%),而美洲土著人最低(4.4%)。整体来看,CYP4F2*3等位基因频率在人群的分布如下:高加索人(34.2%)>中国人(24.7%)>非裔美国人(11.7%)(见表5)。
2.4 汉族和回族及与其他世界人群的遗传关系
通过主成分分析考察了本研究汉族和回族及其与其他世界人群的遗传关系。汉族和回族与东亚人(CHB、CHD、HCB、Bai Chinese、JPT、Tibetan Chinese、Indonesian、Asian、Korean)距离较近,而与其他人群距离较远(见图5)。2个主成分共计达到CYP2C9*3、VKORC1-1639G>A和CYP4F2*3遗传变异的93.2%。主成分分析中所用的等位基因频率数据见表3-5。
表3世界人群中CYP2C9*3的等位基因频率
Table3CYP2C9*3allelefrequenciesinworldwidepopulations
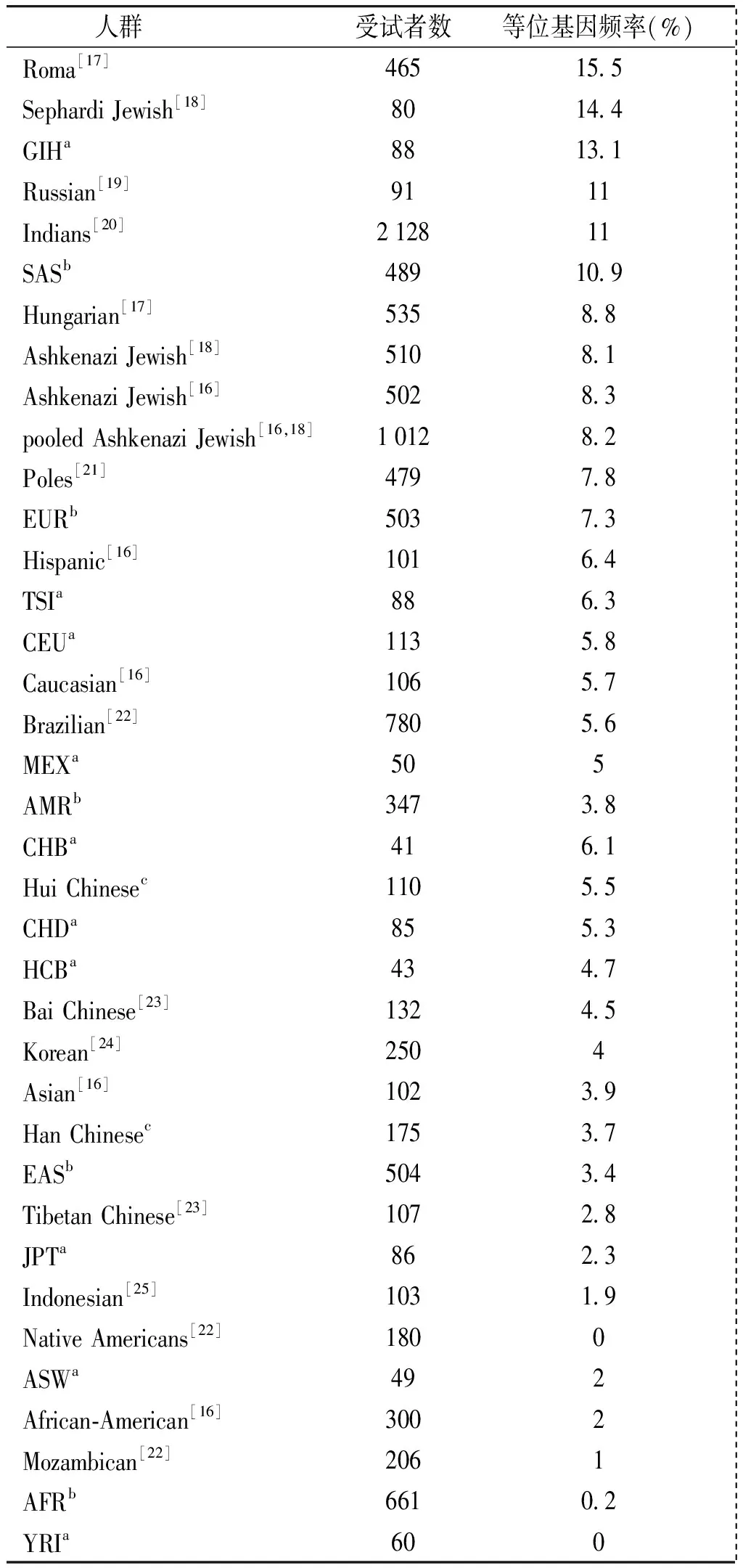
人群受试者数等位基因频率(%) Roma[17]46515 5 SephardiJewish[18]8014 4 GIHa8813 1 Russian[19]9111 Indians[20]212811 SASb48910 9 Hungarian[17]5358 8 AshkenaziJewish[18]5108 1 AshkenaziJewish[16]5028 3 pooledAshkenaziJewish[16,18]10128 2 Poles[21]4797 8 EURb5037 3 Hispanic[16]1016 4 TSIa886 3 CEUa1135 8 Caucasian[16]1065 7 Brazilian[22]7805 6 MEXa505 AMRb3473 8 CHBa416 1 HuiChinesec1105 5 CHDa855 3 HCBa434 7 BaiChinese[23]1324 5 Korean[24]2504 Asian[16]1023 9 HanChinesec1753 7 EASb5043 4 TibetanChinese[23]1072 8 JPTa862 3 Indonesian[25]1031 9 NativeAmericans[22]1800 ASWa492 African⁃American[16]3002 Mozambican[22]2061 AFRb6610 2 YRIa600
a国际HapMap工程;b千人基因组工程;c本研究
表4世界人群中VKORC1-1639A的等位基因频率
Table4VKORC1-1639Aallelefrequenciesinworldwidepopulations
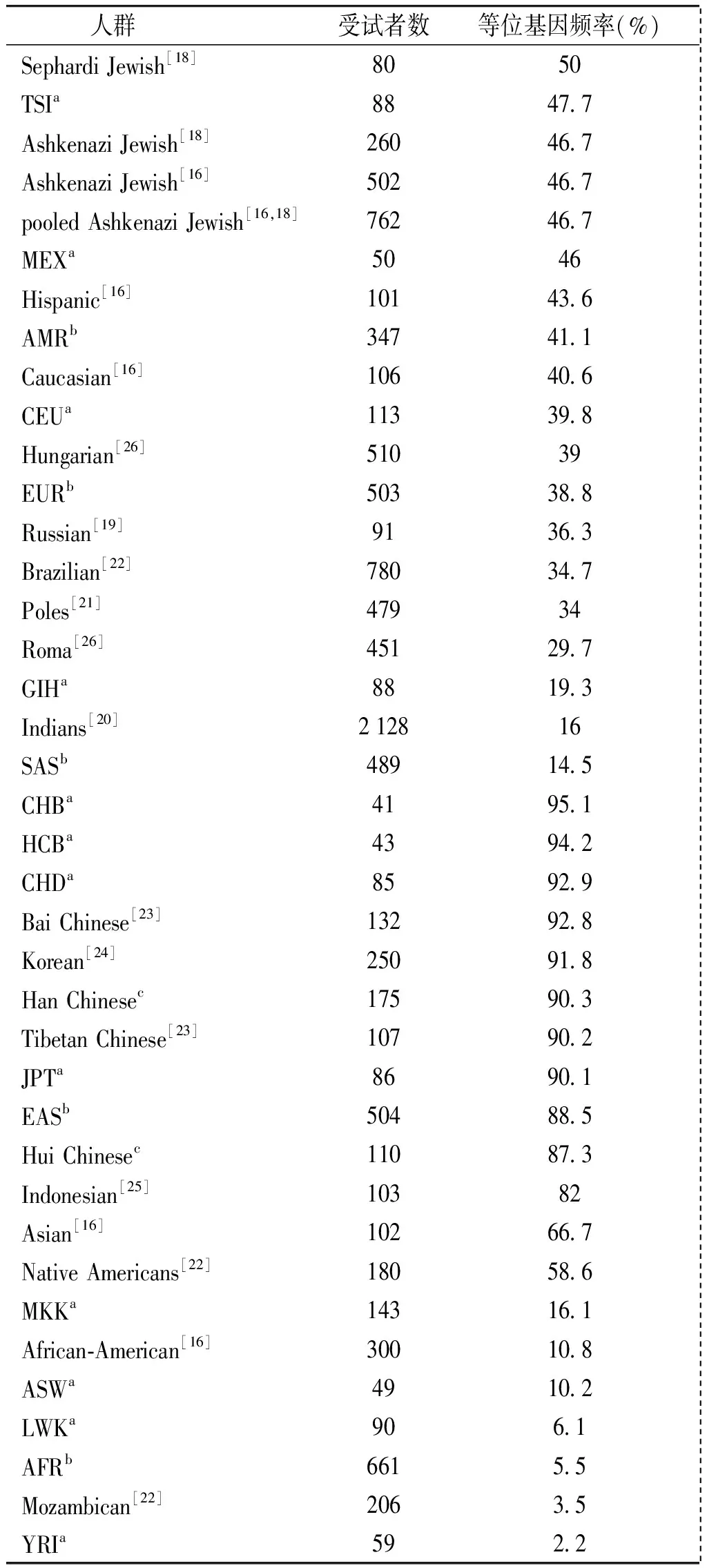
人群受试者数等位基因频率(%) SephardiJewish[18]8050 TSIa8847 7 AshkenaziJewish[18]26046 7 AshkenaziJewish[16]50246 7 pooledAshkenaziJewish[16,18]76246 7 MEXa5046 Hispanic[16]10143 6 AMRb34741 1 Caucasian[16]10640 6 CEUa11339 8 Hungarian[26]51039 EURb50338 8 Russian[19]9136 3 Brazilian[22]78034 7 Poles[21]47934 Roma[26]45129 7 GIHa8819 3 Indians[20]212816 SASb48914 5 CHBa4195 1 HCBa4394 2 CHDa8592 9 BaiChinese[23]13292 8 Korean[24]25091 8 HanChinesec17590 3 TibetanChinese[23]10790 2 JPTa8690 1 EASb50488 5 HuiChinesec11087 3 Indonesian[25]10382 Asian[16]10266 7 NativeAmericans[22]18058 6 MKKa14316 1 African⁃American[16]30010 8 ASWa4910 2 LWKa906 1 AFRb6615 5 Mozambican[22]2063 5 YRIa592 2
a国际HapMap工程;b千人基因组工程;c本研究
3 讨论
华法林主要经肝脏CYP2C9代谢,其编码基因有60余变异等位基因位点。其中CYP2C9*3(c.1075A>C,rs1057910)是位于第7外显子的一个单核苷酸多态性位点,突变之后引起第359氨基酸残基异亮氨酸变为亮氨酸,使得CYP2C9代谢华法林的酶活性下降。因此,此变异体携带者(CYP2C9*1/*3和CYP2C9*3/*3)血浆华法林浓度增加,变得对华法林敏感从而需要减少其维持剂量[11]。VKORC1通过将维生素K环氧化物(KO)还原为还原型维生素K(KH2)而参与体内维生素K的循环,华法林结构与维生素K类似可竞争性作用于VKORC1,后者因而是华法林的药物作用靶点[13]。VKORC1基因启动子区一个单核苷酸多态性位点(VKORC1 g.-1639G>A,rs9923231)引起启动子活性下降,因此突变等位基因携带者(-1639GA和-1639AA)VKORC1活性下降,需减少华法林维持剂量[12]。本研究结果显示CYP2C9*3等位基因频率在中原地区汉族和回族之间无显著差异,且与高加索人无显著差异,但都显著高于非裔美国人。而中原地区汉族和回族VKORC1-1639A等位基因频率显著高于高加索人和非裔美国人,这可以解释临床上观察到的东亚人华法林剂量普遍高于西方人的现象。
表5世界人群中CYP4F2*3的等位基因频率
Table5CYP4F2*3allelefrequenciesinworldwidepopulations
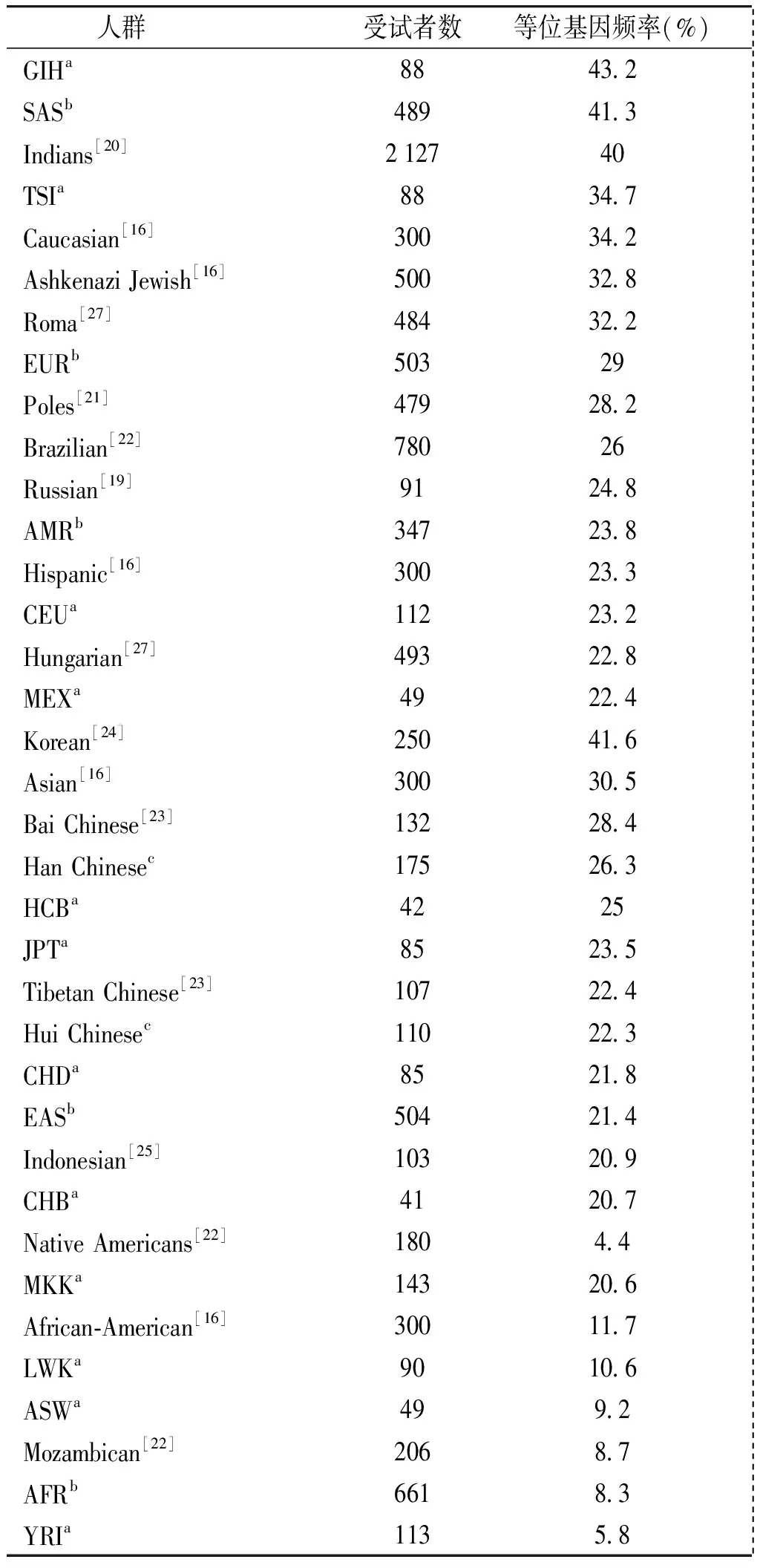
人群受试者数等位基因频率(%) GIHa8843 2 SASb48941 3 Indians[20]212740 TSIa8834 7 Caucasian[16]30034 2 AshkenaziJewish[16]50032 8 Roma[27]48432 2 EURb50329 Poles[21]47928 2 Brazilian[22]78026 Russian[19]9124 8 AMRb34723 8 Hispanic[16]30023 3 CEUa11223 2 Hungarian[27]49322 8 MEXa4922 4 Korean[24]25041 6 Asian[16]30030 5 BaiChinese[23]13228 4 HanChinesec17526 3 HCBa4225 JPTa8523 5 TibetanChinese[23]10722 4 HuiChinesec11022 3 CHDa8521 8 EASb50421 4 Indonesian[25]10320 9 CHBa4120 7 NativeAmericans[22]1804 4 MKKa14320 6 African⁃American[16]30011 7 LWKa9010 6 ASWa499 2 Mozambican[22]2068 7 AFRb6618 3 YRIa1135 8
a国际HapMap工程;b千人基因组工程;c本研究
CYP4F2通过氧化维生素K侧链而限制体内可利用维生素K的量,其编码基因一个变异体(CYP4F2*3,c.1297G>A,rs2108622)引起第433氨基酸残基缬氨酸突变为甲硫氨酸,突变之后CYP4F2活性下降使得体内可利用维生素K量增加,因此此变异体携带者(CYP4F2*1/*3和CYP4F2*3/*3)需增加华法林的维持剂量[15]。本研究结果显示中原地区汉族和回族CYP4F2*3等位基因频率介于高加索人和非裔美国人之间,考虑到CYP4F2*3与华法林剂量调整的相关性,应将其纳入华法林药物基因检测。
此外,将本研究结果与已报道世界其他人群进行了比较,并在世界范围内利用主成分分析确定了中原地区汉族和回族之间及其与其他人群的遗传距离。结果提示中原地区汉族和回族与东亚人群距离较近。有意思的是,我们的分析还可发现欧洲的罗姆人(吉普赛人)可能起源于南亚,因其与南亚人群距离较近,这些结果也与早期文献报道一致[28,29]。本研究的分析也发现美洲土著人与其他人群距离较远,这可能反应了其与欧亚人群分离后的遗传孤立现象[30,31]。
综上所述,中原地区汉族和回族CYP2C9*3、VKORC1-1639G>A和CYP4F2*3遗传多态性分布频率相似但与高加索和非洲人群存在显著差异。在中国人群香豆素类抗凝剂药物基因检测中应该将CYP2C9*3、VKORC1-1639G>A和CYP4F2*3遗传多态性都包含进去以利于此类药物剂量的调整,促进中国人群抗凝药物治疗的个体化和精准化。
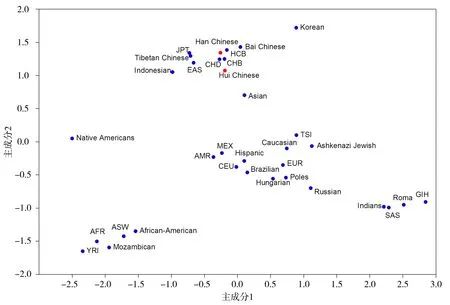
利用34个人群的等位基因频率进行主成分分析,将前2个主成分即主成分1和主成分2作点图,其中汉族和回族用红色实心圆表示,而其他32个文献已报道的人群用蓝色实心圆表示图5 通过主成分分析对汉族和回族及其与世界其他人群的遗传关系的分析Figure 5 Genetic relationships between the Han and Hui Chinese populations and their genetic relations to 32 other world populations by principal component analysis
[1] Husted SE, Ziegler BK, Kher A. Long-term anticoagulant therapy in patients with coronary artery disease[J]. Eur Heart J, 2006, 27(8): 913-919.
[2] Ghozlan MF, Foad DA, Darwish YW,etal. Impact of CYP2C9 and VKORC1 genetic polymorphisms upon warfarin dose requirements in Egyptian patients with acute coronary syndrome[J]. Blood Coagul Fibrinolysis, 2015,26(5):499-504.
[3] de Oliveira Almeida VC, Ribeiro DD, Gomes KB,etal. Polymorphisms of CYP2C9, VKORC1, MDR1, APOE and UGT1A1 genes and the therapeutic warfarin dose in Brazilian patients with thrombosis: a prospective cohort study[J]. Mol Diagn Ther, 2014,18(6):675-683.
[4] Nahar R,Saxena R,Deb R,etal.CYP2C9,VKORC1,CYP4F2,ABCB1 and F5 variants: influence on quality of long-term anticoagulation[J]. Pharmacol Rep,2014,66(2):243-249.
[5] Perera MA, Cavallari LH, Limdi NA,etal. Genetic variants associated with warfarin dose in African-American individuals: a genome-wide association study[J]. Lancet, 2013,382(9894):790-796.
[6] Cha PC, Mushiroda T, Takahashi A,etal. Genome-wide association study identifies genetic determinants of warfarin responsiveness for Japanese[J]. Hum Mol Genet, 2010,19(23):4735-4744.
[7] Teichert M, Eijgelsheim M, Rivadeneira F,etal. A genome-wide association study of acenocoumarol maintenance dosage[J]. Hum Mol Genet, 2009,18(19):3758-3768.
[8] Takeuchi F, McGinnis R, Bourgeois S,etal. A genome-wide association study confirms VKORC1, CYP2C9, and CYP4F2 as principal genetic determinants of warfarin dose[J]. PLoS Genet, 2009,5(3):e1000433.
[9] Cooper GM, Johnson JA, Langaee TY,etal. A genome-wide scan for common genetic variants with a large influence on warfarin maintenance dose[J]. Blood, 2008,112(4):1022-1027.
[10] Guo H, Yan J, Jiao Z,etal. Genetic polymorphisms for 17 Y-chromosomal STRs haplotypes in Chinese Hui population[J]. Leg Med (Tokyo), 2008,10(3):163-169.
[11] Steward DJ, Haining RL, Henne KR,etal. Genetic association between sensitivity to warfarin and expression of CYP2C9*3[J]. Pharmacogenetics, 1997,7(5):361-367.
[12] Rieder MJ, Reiner AP, Gage BF,etal. Effect of VKORC1 haplotypes on transcriptional regulation and warfarin dose[J]. N Engl J Med, 2005,352(22):2285-2293.
[13] Tie JK, Stafford DW. Structural and functional insights into enzymes of the vitamin K cycle[J]. J Thromb Haemost, 2016,14(2):236-247.
[14] Spreafico M, Lodigiani C, van Leeuwen Y,etal. Effects of CYP2C9 and VKORC1 on INR variations and dose requirements during initial phase of anticoagulant therapy[J]. Pharmacogenomics, 2008,9(9):1237-1250.
[15] Alvarellos ML, Sangkuhl K, Daneshjou R,etal. PharmGKB summary: very important pharmacogene information for CYP4F2[J]. Pharmacogenet Genomics, 2015,25(1):41-47.
[16] Scott SA, Khasawneh R, Peter I,etal. Combined CYP2C9, VKORC1 and CYP4F2 frequencies among racial and ethnic groups[J]. Pharmacogenomics, 2010,11(6):781-791.
[17] Sipeky C, Lakner L, Szabo M,etal. Interethnic differences of CYP2C9 alleles in healthy Hungarian and Roma population samples: relationship to worldwide allelic frequencies[J]. Blood Cells Mol Dis, 2009,43(3):239-242.
[18] Scott SA, Edelmann L, Kornreich R,etal. Warfarin pharmacogenetics: CYP2C9 and VKORC1 genotypes predict different sensitivity and resistance frequencies in the Ashkenazi and Sephardi Jewish populations[J]. Am J Hum Genet, 2008,82(2):495-500.
[19] Ivashchenko D, Rusin I, Sychev D,etal. The frequency of CYP2C9, VKORC1, and CYP4F2 polymorphisms in Russian patients with high thrombotic risk[J]. Medicina (Kaunas), 2013,49(12):517-521.
[20] Giri AK, Khan NM, Grover S,etal. Genetic epidemiology of pharmacogenetic variations in CYP2C9, CYP4F2 and VKORC1 genes associated with warfarin dosage in the Indian population[J]. Pharmacogenomics, 2014,15(10):1337-1354.
[21] Wypasek E, Branicka A, Awsiuk M,etal. Genetic determinants of acenocoumarol and warfarin maintenance dose requirements in Slavic population: a potential role of CYP4F2 and GGCX polymorphisms[J]. Thromb Res, 2014,134(3):604-609.
[22] Vargens DD, Damasceno A, Petzl-Erler ML,etal. Combined CYP2C9, VKORC1 and CYP4F2 frequencies among Amerindians, Mozambicans and Brazilians[J]. Pharmacogenomics, 2011,12(6):769-772.
[23] Zeng WT, Zheng QS, Huang M,etal. Genetic polymorphisms of VKORC1, CYP2C9, CYP4F2 in Bai, Tibetan Chinese[J]. Pharmazie, 2012,67(1):69-73.
[24] Kim KA, Song WG, Lee HM,etal. Multiplex pyrosequencing method to determine CYP2C9*3, VKORC1*2, and CYP4F2*3 polymorphisms simultaneously: its application to a Korean population and comparisons with other ethnic groups[J]. Mol Biol Rep, 2014,41(11):7305-7312.
[25] Rusdiana T, Araki T, Nakamura T,etal. Responsiveness to low-dose warfarin associated with genetic variants of VKORC1, CYP2C9, CYP2C19, and CYP4F2 in an Indonesian population[J]. Eur J Clin Pharmacol, 2013,69(3):395-405.
[26] Sipeky C, Csongei V, Jaromi L,etal. Vitamin K epoxide reductase complex 1 (VKORC1) haplotypes in healthy Hungarian and Roma population samples[J]. Pharmacogenomics, 2009,10(6):1025-1032.
[27] Sipeky C, Weber A, Melegh BI,etal. Interethnic variability of CYP4F2 (V433M) in admixed population of Roma and Hungarians[J]. Environ Toxicol Pharmacol, 2015,40(1):280-283.
[28] Rai N, Chaubey G, Tamang R,etal. The phylogeography of Y-chromosome haplogroup h1a1a-m82 reveals the likely Indian origin of the European Romani populations[J]. PLoS One, 2012,7(11):e48477.
[29] Moorjani P, Patterson N, Loh PR,etal. Reconstructing Roma history from genome-wide data[J]. PLoS One, 2013,8(3):e58633.
[30] Rasmussen M, Sikora M, Albrechtsen A,etal. The ancestry and affiliations of Kennewick Man[J]. Nature, 2015,523(7561):455-458.
[31] Raghavan M, Steinrücken M, Harris K,etal. Genomic evidence for the Pleistocene and recent population history of Native Americans[J]. Science, 2015,349(6250):aab3884.
GeneticpolymorphismsofCYP2C9*3,VKORC1-1639G>AandCYP4F2*3amongHanandHuiChinesepopulationsofcentralChina
QI Guangzhao1,BO Hui2,WANG Weijie3*
(1DepartmentofPharmacy,FirstAffiliatedHospitalofZhengzhouUniversity,Zhengzhou450052,China;2SchoolofMedicine,HuangheScienceandTechnologyCollege;3DepartmentofHepatobiliaryandPancreaticSurgery,FirstAffiliatedHospitalofZhengzhouUniversity;*Correspondingauthor,E-mail:cyp450239@126.com)
ObjectiveTo investigate the genetic polymorphisms of CYP2C9*3, VKORC1-1639G>A and CYP4F2*3 among Han and Hui Chinese populations of central China.MethodsTwo hundred and eighty-five unrelated Chinese healthy subjects(175 Han and 110 Hui) were used for genotyping. Frequencies of CYP2C9*3 and VKORC1-1639G>A were determined by the polymerase chain reaction-restriction fragment length polymorphism(PCR-RFLP) method and frequencies of CYP4F2*3 were determined by PCR-Sanger’ sequencing. Allele frequencies of CYP2C9*3, VKORC1-1639G>A and CYP4F2*3 in Han and Hui Chinese populations were compared to those published previously in Caucasian and African.ResultsThe allele frequencies of CYP2C9*3 in Han and Hui Chinese were 3.7% and 5.5%, respectively. The allele frequency of CYP2C9*3 in Hui Chinese was significantly higher than that reported previously in African-American(2.0%,P<0.05). The frequencies of VKORC1-1639A in Han and Hui were 90.3% and 87.3%, respectively. The allele frequencies of VKORC1-1639A in both Han and Hui Chinese were significantly higher than those reported previously in African-American(10.8%,P<0.05) and Caucasian (40.6%,P<0.05). The allele frequencies of CYP4F2*3 in Han and Hui Chinese were 26.3% and 22.3% respectively. The allele frequencies of CYP4F2*3 in both Han and Hui Chinese were significantly higher than that reported previously in African-American (11.7%,P<0.05), but lower than that in Caucasian (34.2%,P<0.05).ConclusionThe frequencies of CYP2C9*3, VKORC1-1639G>A and CYP4F2*3 genetic polymorphisms between Han and Hui Chinese populations are similar but significantly different from those in Caucasian and African.
CYP2C9; CYP4F2; VKORC1; Hui nationality; Han nationality; genetic polymorphism
齐光照,男,1987-12生,博士,主管药师,E-mail:guangzhao-qi@qq.com
2017-07-31
R181
A
1007-6611(2017)12-1265-08
10.13753/j.issn.1007-6611.2017.12.014
