四种引进椴在大连的春季抗寒机制分析
2017-12-19陈美谕徐程扬李金航
陈美谕,徐程扬,李金航,袁 振
(北京林业大学省部共建森林培育与保护教育部重点实验室,干旱、半干旱地区森林培育和森林生态系统国家林业局重点实验室,北京 100083)
四种引进椴在大连的春季抗寒机制分析
陈美谕,徐程扬,李金航,袁 振
(北京林业大学省部共建森林培育与保护教育部重点实验室,干旱、半干旱地区森林培育和森林生态系统国家林业局重点实验室,北京 100083)
以四种引进椴:欧洲小叶椴Tilia cordata、欧洲大叶椴Tilia platyphylla、银椴Tilia tomentosa和美洲椴Tilia america的幼树枝条为材料,分别于幼树生长的萌芽期和叶片伸展期,采用室内等速降温试验法,通过研究枝条生理指标变化特征以及枝条解剖形态特征与半致死温度的关系,探寻早春时节四种引进椴的抗寒能力差异及抗寒机理。研究结果表明:四种椴树进行抗寒调节的形态和生理指标随处理温度的改变而发生不同变化,是引起其抗寒能力差异的主要原因。萌芽期引进椴对可溶性糖、脯氨酸和丙二醛含量的调控能力是影响植株抗寒力的重要因素,而在展叶期,椴树的抗寒力则主要受结构指标以及丙二醛含量的影响。此外,四种北美引进椴的抗寒能力在不同物候期存在显著差异(p<0.05),其中,银椴在萌芽期的抗寒能力最强,而美洲椴在展叶期的抗寒能力最强。欧洲小叶椴、欧洲大叶椴、银椴和美洲椴在萌芽期的半致死温度分别为-15.1 ℃、-17.4 ℃、-23.0℃和-20.8℃,在展叶期则分别为6.0 ℃、4.7 ℃、5.5 ℃和4.3 ℃。结合大连地区两个物候期时的气候状况,对引种椴早春防寒工作提出如下建议:在萌芽期,引种椴受冻害概率较低,相应防寒措施应重点集中于展叶期进行。
北美椴;抗寒;生理指标;结构指标;差异分析
早春降温造成的低温胁迫是影响植物生长发育的关键因素,能否抵御“倒春寒”的影响决定引种植物的成活率和长势好坏,探究生长期植物的抗寒能力与影响耐寒力的指标对于引种过程中的经营手段有着指导作用。对于落叶树种来说,植物枝条已经经历之前秋冬的低温锻炼,但叶片或未经历低温锻炼或非正常越冬,不具试验参考价值,因此,在早春针对枝条进行降温试验更具现实意义。在遭遇低温胁迫时,短期内植物体内会通过脯氨酸[1]和可溶性糖浓度[2]变化发生渗透调节[3]或通过丙二醛含量变化发生膜质相变[4],严重胁迫时,则致使细胞膜破裂而发生电解质渗透[5];为了长期适应低温胁迫环境,植物通过改变木质部在枝条中的比例而提高抗寒能力[6]。遗传差异对植物抗寒性的影响极大,但环境变化在一定程度上会改变这种现象[7-9]。
金桐力是从北美北部地区引进并成功驯化的椴树属树种的商品名统称[10],包括欧洲小叶椴Tilia cordata、欧洲大叶椴Tilia platyphylla、银椴Tilia tomentosa、美洲椴Tilia america,其优点是树形优美,病虫害少,树龄长。目前,该树种被引种到辽宁大连等地用于城市绿化[11],在当地生长表现良好,但对于其抗寒能力未见系统研究与报道。春季寒流通常会致使抗寒力较差的植物遭受冻害和寒害,而金桐力能够在多大程度上抵御寒潮危害尚不得知。引进种在萌芽期和展叶期的抗寒能力是评价其抗寒性的重要标志[12],然而,目前在该方面的研究报道较少。
因此,为了科学评价金桐力的抗寒能力,本研究以大连地区燕园农业公司引种栽植的3年生欧洲小叶椴、欧洲大叶椴、美洲椴、银椴枝条为材料,分别在春季萌芽期和展叶期,采用人工模拟试验的方法,从形态特征与枝条生理指标变化两个方面对抗寒性进行评价,探讨各生理与结构指标与抗寒性的关系,以期为在我国北方城市栽培推广金桐力提供参考。
1 试验地概况
试验地在位于辽宁省大连市保税区二十里堡镇的燕园农业苗木基地 (39.20 °N,121.80 °E)。试验地土壤类型为黄棕壤,土壤全磷、全氮、全钾、有机质含量分别为1.50、0.58、0.33、19.26 g/kg。年平均气温10 ℃左右,年极端高温和极端低温分别约35 ℃和-20 ℃,无霜期180 ~ 200 d。年平均降水量550 ~ 950 mm,全年降水多集中于夏季,夜雨量多于日雨量。夏季盛行偏南季风,冬季盛行偏北季风,6级及6级以上大风30~40 d,多发生于春、冬两季。
2 材料和方法
2.1 试验材料
分别于2015年3月30日和4月15日,选择树势中等的3年生欧洲小叶椴、欧洲大叶椴、美洲椴、银椴幼树各4株,于每株树冠外围剪取高度一致、东南朝向的一年生枝条若干,用湿报纸和无纺布对枝条分别进行内包裹和外包装后带回实验室[13]。先后利用清水和去离子水将枝条样品清洗干净,待轻轻吸干枝条表面残存水分后,迅速用石蜡对枝条两端进行封口,并用保鲜膜将新鲜枝条包裹以防止其大量失水,包裹完毕后,将所有样品置于8 ℃冰箱中保存备用。
低温试验开始前,取出保鲜膜内的枝条并迅速将其表面水分吸干,随后将其置于低温冰箱内进行冷冻处理。待经过低温处理的枝条解冻后,避开芽眼,将其剪成2 ~ 3 cm长的小段以备各指标的测定。
2.2 研究方法
2.2.1 试验设计
大连地区树木春季萌芽期和展叶期时的平均气温分别约6.82 ℃和10.31 ℃。由于受西伯利亚寒流的影响,北方在这两个阶段的气候变化通常比较剧烈,因而萌芽期和展叶期为北方树木遭受冻害最严重的两个时期[14]。本试验采用室内控制模拟降温的方法,可以较好模拟早春植物遭遇寒流来袭的响应状态,在萌芽期和展叶期分别对4个引进种枝条进行控制降温处理,其中,萌芽期的温度设置为0 ℃、-5 ℃、-10 ℃、-15 ℃、-20 ℃、-25 ℃,降温速度为3℃/h,展叶期的温度设置为6 ℃、4 ℃、2 ℃、0 ℃、-2 ℃、-4 ℃,降温速度为 3℃ /h。
2.2.2 半致死温度测定
准确称取经过低温处理的枝条0.50 g,加入20 mL去离子水并在室温下浸提8 h后,利用电导仪(CON510)测定溶液电导率值,再将浸提液高温煮沸1 h,于室温冷却后再次测定溶液电导率值[15]。相对电导值(REC)的计算如下公式:
相对电导值(REC)=(室温电导值-蒸馏水电导值)/(煮沸后电导值-蒸馏水电导值)。
参照郭海林等[16]的方法,利用相对电导率拟合逻辑斯蒂克方程Y=K/(1+ae-bt),其中:K为相对电导率的饱和值,a、b为方程参数,t为设定试验温度。确定参数拟合方程后,检验拟合度R2的显著程度,若结果显著,则利用LT50=-ln(a/b)计算半致死温度 (LT50)。
2.2.3 生理指标测定
脯氨酸含量的测定采用酸性茚三酮法[17],可溶性糖含量的测定采用蒽酮显色法[18],丙二醛含量的测定采用TCA显色法[19]。
2.2.4 结构观察方法
利用杨捷频改良的石蜡切片法[20]对枝条样品横切解剖结构进行观察。经过FAA固定液固定、脱水、透蜡、包埋、修块、切片、展片、贴片、染色、干燥处理后,将制成的枝条石蜡切片置于OLYMPUS-DP72显微镜下观察。
2.3 数据处理与计算
利用Microsoft Of fi ce Excel 2010制作图表,SPSS 12.0 (SPSS for Windows, Chicago, USA)进行相关性分析和因子分析,G × T (Genotype ×Trait)和GGE(G × GE)双标图利用GGEbiplot 7.9 (Copyright © 严威凯)进行绘制。
为了计算不同温度梯度区间生理指标变化速率,将萌发期温度变化划分成五个温度变化区间分为 0 ℃ ~ -5 ℃(TB1)、-5 ℃ ~ -10 ℃(TB2)、-10 ℃ ~ -15 ℃(TB3)、-15 ℃ ~ -20 ℃(TB4)、-20 ℃ ~ -25 ℃(TB5),相应地将叶片展叶期温度变化划分成五个温度变化区间分为6 ℃ ~ 4 ℃(TL1)、4 ℃ ~ 2 ℃(TL2)、2 ℃ ~ 0 ℃(TL3)、0 ℃ ~ -2 ℃(TL4)、-2 ℃ ~ -4 ℃(TL5)。
(1)脯氨酸含量相对变化量(Pr) =(第r温度区间上降温后脯氨酸含量 - 第r温度区间上降温前脯氨酸含量)/ 第r温度区间上降温前脯氨酸含量;
(2)可溶性糖含量相对变化量(SSr)=(第r温度区间上降温后可溶性糖含量 - 第r温度区间上降温前可溶性糖含量) / 第r温度区间上降温前可溶性糖含量;
(3)丙二醛含量相对变化量(Mr)=(第r温度梯度上降温后丙二醛含量 - 第r温度梯度上降温前丙二醛含量) / 第r温度梯度上降温前丙二醛含量。
(4)木质部宽+韧皮部宽(L1)= lm+ lr;
(5)才皮比((木质部+韧皮部)宽度/皮部宽度,C1) = (lm+ lr)/ lp;
(6)木质部宽/枝条直径径(C2) = lm/lb;
(7)木质部面积/枝条断面积(C3)=lm2/lb2;
(8)韧皮部宽/枝条直径(C4) = lr/ lbs;
其中,lp为皮部厚度(mm)、lm为木质部半径(mm)、lr为韧皮部半径(mm)、lb为枝条半径(mm)。
3 结果分析
3.1 引进椴树春季抗寒性及其种间差异
各椴树品种枝条在萌芽期的抗寒能力明显高于展叶期。在同一物候期,不同椴树品种的枝条抗寒力也有显著差异(表1)。其中,银椴在萌芽期的抗寒力最高,半致死温度分别比欧洲小叶椴、欧洲大叶椴和美洲椴高出52.32%、32.18%、10.58%;美洲椴在展叶期的抗寒力最高,半致死温度分别比小叶椴、欧洲大叶椴和银椴高出27.87%、8.33%、21.43%。
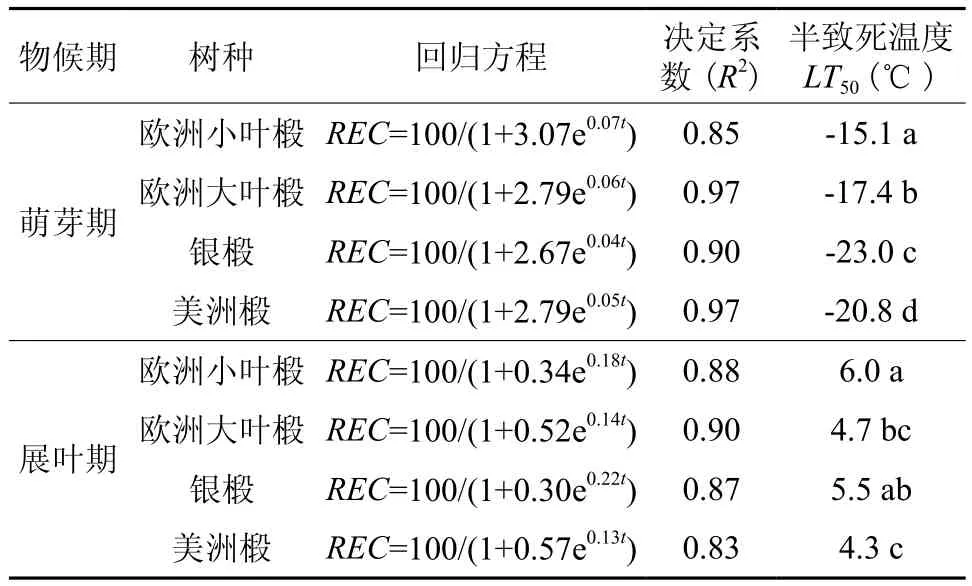
表1 两个物候期四种引进椴枝条电导率模型和半致死温度†Table 1 The electrical conductivity models and half lethal temperature of branch of four linden species in two phenological phases
3.2 引进椴春季抗寒性的枝条结构与生理机理
3.2.1 枝条解剖结构与抗寒性
在萌芽期,银椴枝条L1、C1、C2、C3指标值最大,且C1较其他三个引进种分别高出6.50%、17.70%和6.60%,展叶期,美洲椴枝条的以上指标略高于银椴,四种引进椴枝条解剖结构指标在两个物候期期间均未达显著差异(图1)。因此,枝条解剖结构并非造成四种引进椴抗寒性差异的主导因素。
3.2.2 可溶性糖、脯氨酸和丙二醛含量变化与抗寒性
在萌芽期,四种引进椴枝条中可溶性糖含量变化幅度较大;展叶期,枝条中的脯氨酸含量和丙二醛含量变化更剧烈(图2)。在萌芽期,枝条处理温度在TB3阶段时,脯氨酸和丙二醛的含量增速显著升高,欧洲小叶椴增速最大,分别达到22.00%和230.00%;可溶性糖含量呈现先降后升趋势,在TB4时,美洲椴变化率达到250.00%。在展叶期枝条的生理指标发生明显增加的主要温度梯度在TL1阶段,在这一阶段银椴脯氨酸变化率达到150.00%,丙二醛变化率为373.00%;可溶性糖含量没有出现先降后升的趋势,欧洲小叶椴在TL3阶段可溶性糖相对变化量达最大,仅为39.00%。
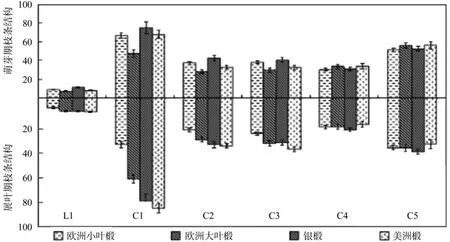
图1 两个物候期枝条解剖结构指标的变化Fig.1 Change of structure indices of sapling branches in two periods
在萌芽期,可溶性糖变化动态与种间抗寒性之间并非完全同步,其中,半致死温度最高的欧洲小叶椴枝条中可溶性糖含量,随着温度的快速降低并未呈现出其他三个种的剧烈降低趋势;温度从0 ℃降至-15 ℃,欧洲小叶椴枝条中可溶性糖浓度仅改变了9.75 mg/g,而其余三个种则降低了43.75 ~ 83.33%。在展叶期,半致死温度最低的美洲椴则在丙二醛含量上呈大幅度提高趋势;温度从6 ℃降到2 ℃,美洲椴枝条丙二醛含量提高了400.00%,而其余三种则仅提高了18.00 ~170.00%不等(图2)。
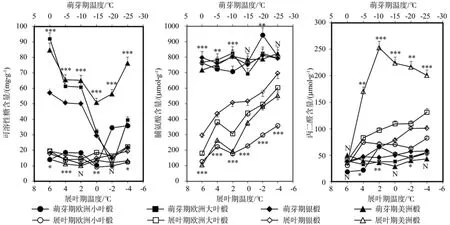
图2 两个物候期椴树枝条生理指标变化动态Fig.2 Dynamic of branches physiological indices during bud bursting period and leave outspreading period
3.2.3 引进椴抗寒性的综合分析
在萌芽期,枝条的抗寒反应与脯氨酸、可溶性糖以及丙二醛含量变化均有密切联系;而在展叶期,枝条的抗寒反应则与丙二醛含量变化的关系更加密切,且更多受结构指标的影响(表2)。萌芽期引进椴在TB2与TB5两个温度梯度上,脯氨酸相对变化量与LT50呈极显著负相关;在TB3与TB4温度梯度上,可溶性糖含量相对变化量与抗寒能力达到显著相关水平;在TB2与TB3两个温度梯度上,MDA含量的相对变化量与抗寒能力达到极显著负相关水平。展叶期枝条在TL2阶段的脯氨酸含量相对变化量,TL1、TL4可溶糖含量的相对变化量以及TL1、TL3、TL4温度梯度上丙二醛含量的相对变化量与抗寒性呈显著相关,此外,L1、C1、C4、C5与半致死温度均成显著负相关。
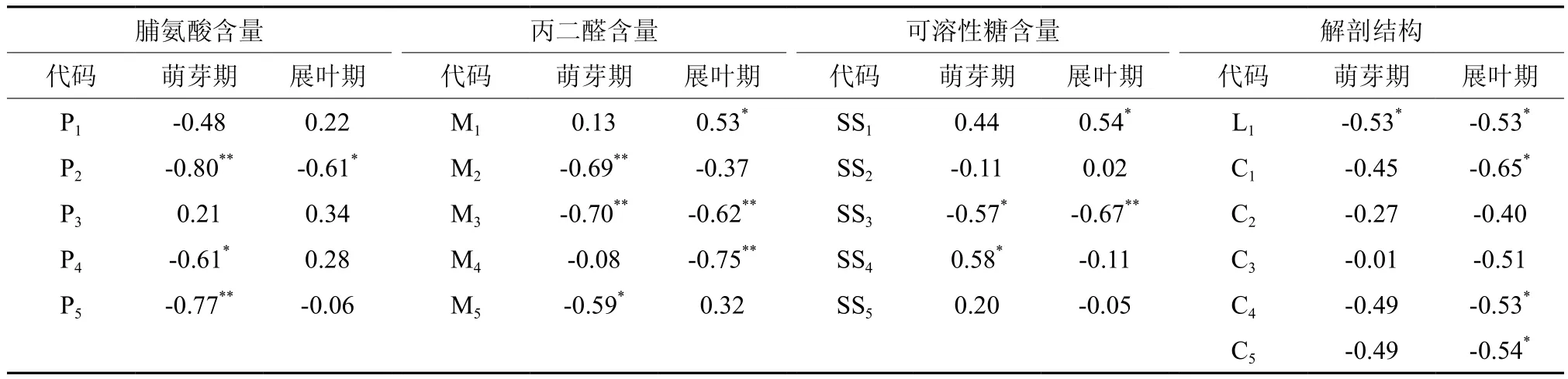
表2 枝条各指标相对变化量与半致死温度(LT50)的简相关性分析†Table 2 Correlation between the increments of physiological indices and LT50
3.3 引进椴树木春季抗寒对策的种间差异
选取与抗寒能力显著(p< 0.05)相关的生理指标(表3)绘制主成分分析双标图的结果表明,四个引进椴种间春季抗寒的对策明显不同。在萌芽期,TB3时的可溶性糖含量变化率、TB5和TB2时的脯氨酸含量变化率对银椴抗寒能力贡献较大,TB5时的脯氨酸含量变化率、TB2和TB3的丙二醛含量变化率对美洲椴抗寒能力贡献较大,TB2和TB3的丙二醛含量变化率更好地解释了欧洲大叶椴的抗寒能力,而TB5的丙二醛含量变化率、TB4的可溶性糖和脯氨酸含量变化率更好地反映了欧洲小叶椴的抗寒能力(图3A)。在展叶期,TL3时的丙二醛含量变化率可很好地解释美洲椴的抗寒能力,TL3和TL4时的丙二醛含量变化率可解释美洲和欧洲大叶椴的抗寒能力,TL1时的可溶性糖、丙二醛和脯氨酸含量变化率较好地反映了银椴的抗寒能力,而TL3的可溶性糖含量变化率以及TL4的丙二醛含量变化率则是对欧洲小叶椴抗寒能力贡献较高的指标(图3B)。
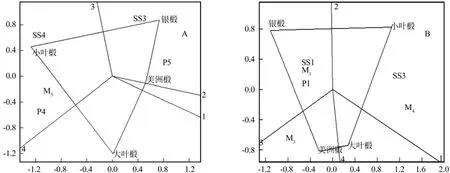
图3 萌芽期(A)和展叶期(B)不同引进椴品种与不同生理指标含量变化率之间的关系Fig. 3 Relationship between species and physiological indexes change rate during bud bursting period and leave outspreading period
这表明:在萌芽期,面对急速降温环境,银椴主要采取了可溶性糖和脯氨酸含量变化的短期响应对策,丙二醛含量变化是欧洲大叶椴应对急速降温环境的核心响应对策,脯氨酸含量和丙二醛含量变化是美洲椴短期响应急速降温环境的主要对策,欧洲小叶椴则是采取了丙二醛、可溶性糖和脯氨酸协同变化是短期响应急速降温环境的对策。同样,在展叶期急速降温环境过程中,美洲椴和欧洲大叶椴针对急速降温主要通过丙二醛含量变化进行短期响应的,银椴是通过丙二醛、可溶性糖与脯氨酸含量三者协同变化响应,而欧洲小叶椴的主要响应对策则是可溶性糖含量与丙二醛含量的协同变化。
4 讨 论
4.1 两个生长期结构指标与生理指标的抗寒机理差异
植物在对寒冷环境的长期适应过程中形成一系列结构特征[21],同时,环境温度急速下降时,低温会诱导体内生理指标含量改变以抵御寒冷[22]。结构特征与生理指标含量的变化均可以反映植株对寒冷的抵抗能力,但在植株生长的不同时期,二者对抗寒能力的作用大小并不相同。
生理指标发生显著变化的时期往往集中在早春和初秋[23],此时环境温度忽然降低,植株进行低温锻炼,通过渗透调节改变细胞内渗透势,并改变膜质结构以抵御寒冷胁迫[24]。而在其他时期,生理指标对于温度改变响应不敏感,抗寒力主要由枝条结构体现[23]。
本试验中萌芽期(早春时期)生理指标的相对变化是衡量抗寒能力的主要指标,结构指标中只有木质部与韧皮部的半径之和(L1)与半致死温度呈显著相关;虽然采样间隔较短,但在展叶期,枝条处于解除低温驯化状态[25],并进入生长模式,植物体内养分运输较快,叶片生长需要相当多的有机质和糖类物质,生理指标对寒冷胁迫的渗透调节能力较弱,枝条结构指标(L1、C1、C4、C5)成为衡量抗寒能力的主要因素[26]。
结构指标对抗寒力的作用体现在木质化程度上[27]。枝条皮部由活细胞组成,对寒冷胁迫较敏感,而木质部和韧皮部是由死细胞构成的导管组成的,结构较硬,所以木质化可以有效防止因低温产生的细胞膜破裂和原生质变形[28]。枝条结构的形成是一个长期过程,主要由树木品种决定[29]。故本试验中结构指标在两个生长期差异不大,种间也无显著差异。但细微的结构变化都可对抗寒性造成显著影响[30]。美洲椴的各项结构指标在两个时期均略高于其他品种,综合两个时期,其抗寒能力也较强。
4.2 两个生长期生理指标变化的差异
在不同生长时期,生理指标变化对抗寒力的影响也存在差异。可溶性糖调控在抗寒生理中起到重要作用,在植株的休眠期,其调节功能强于脯氨酸[31]。在初始萌芽期,可溶性糖可以通过低温诱导合成,一方面,通过合成反应释放热量以提升细胞内温度,另一方面可溶性糖含量的增加可以提高细胞内渗透势[32]。当植株进入生长期,可溶性糖转而用于营养生长[33],对细胞渗透势的调控作用下降。因此本试验中,在展叶期各品种可溶性糖含量相对变化趋于平缓,相对变化量与半致死温度的相关性也较弱。银椴品种是以可溶性糖含量变化为主要抗寒响应指标的树种,在萌芽期,由于可溶性糖变化较活跃,银椴抗寒能力在四个品种中最强,但进入展叶期后,随可溶性糖变化趋于平缓,其抗寒能力也相应减弱。
当可溶性糖的调节能力减弱,脯氨酸与丙二醛的作用趋于显著。脯氨酸对寒冷较敏感,通常在降温初期,脯氨酸率先响应[27],这解释了两个时期,与半致死温度显著相关的脯氨酸含量相对变化区间均发生在寒冷胁迫的初期,展叶期发生在4 ~ 2 ℃时期,萌芽期由于形成低温胁迫的温度较低,所以-20 ~ -25℃梯度上的脯氨酸含量变化率与半致死温度显著相关。丙二醛是膜质过氧化反应的产物,反映细胞膜的损伤程度[29],膜质氧化反应在一定阶段是可逆的,抗寒能力强的品种清除丙二醛的能力更强,丙二醛含量与抗寒能力呈负相关关系[34]。本试验各品种在萌芽期遇低温时丙二醛变化存在显著差异,其中抗寒能力强的品种如美洲椴及欧洲大叶椴,丙二醛调控作用更显著,主要表现时期在-5 ℃到-15 ℃之间。在展叶期,美洲椴温度降到2 ℃之前,出现了丙二醛含量大幅上升的现象,这与丙二醛对抗寒能力的调控规律不一致,可能由于在这一阶段反映植株抗寒程度的主要指标为可溶性糖变化与枝条结构,2 ℃之后丙二醛开始对美洲椴抗寒能力起调节作用。
试验中萌芽期的可溶性糖含量变化幅度较大,除欧洲小叶椴外,各品种均呈现先降后升的趋势,上升区间在-20 ℃之后。这是由于在遭遇低温初期,处于生长初期的树木会在低温的诱导下向休眠过渡[35],植物体内可溶性糖开始向合成淀粉方向进行,并导致在临界温度前可溶性糖含量先降低[36]。同时,由于枝条已经萌芽,部分可溶性糖用于生长发育[37],这也是导致可溶性糖含量降低的原因之一。当处理温度足够低,引发抗寒机制,细胞内开始诱导可溶性糖合成,一方面,通过合成反应释放热量提升细胞内温度,另一方面可溶性糖含量的增加可以增加细胞内渗透势[32]。所以,欧洲小叶椴作为在该时期抗寒能力最差的树种,在降温初始未出现可溶性糖含量下降,是因为该温度已经构成寒冷胁迫。在试验中,对展叶期各品种枝条降温过程中并未出现可溶性糖含量下降的现象,也是因为设置的初始温度已在半致死温度附近,所以直接启动了抗寒机制,导致可溶性糖合成。
萌芽期可溶性糖含量出现先降后升的趋势,其对于抗寒能力的作用发生在-20℃之后,这一现象可以通过生长期的可溶性糖部分用于调节细胞渗透势从而提升抗寒能力、部分用于合成淀粉等大分子以促进植株生长来解释,但可溶性糖具体如何变化尚未研究,因此,未来可以开展枝条内淀粉含量与各种酶活性关系的研究,这对植物春季抗寒机理与生长机理研究的进一步完善有着重要作用。
4.3 同一时期种间抗寒机理差异
4个引进椴品种的结构指标不存在显著差异,造成种间抗寒差异的因素主要为各品种对寒冷的响应机理。种间存在的抗寒机理差异一部分受品种间遗传因素的影响,另一方面受环境因素影响。在萌芽期,银椴主要采取了可溶性糖和脯氨酸含量变化的短期响应对策,丙二醛含量变化是欧洲大叶椴应对急速降温环境的核心响应对策,脯氨酸和丙二醛含量变化是美洲椴短期响应急速降温环境的主要对策,欧洲小叶椴则是采取了丙二醛、可溶性糖和脯氨酸含量协同变化是短期响应急速降温环境的对策。在展叶期,美洲椴针对急速降温主要是通过丙二醛含量变化来响应的,银椴的降温对策是通过丙二醛、可溶性糖与脯氨酸含量三者协同反映来实现的,欧洲小叶椴的主要对策则是可溶性糖含量与丙二醛含量的协同变化。这也可以解释试验中出现品种抗寒能力与生理物质相对变化不一致的情况,植物的抗寒能力是多项指标共同作用的结果,不可能用单一指标来评价。
5 结 论
(1)品种间由于本身的遗传特性与环境因素共同作用形成抗寒力的差异:在萌芽期,银椴的抗寒能力最强,其次是美洲椴、欧洲大叶椴和欧洲小叶椴,半致死温度分别为-23.0 ℃、-20.8℃、-17.4 ℃和-15.1 ℃;在展叶期,抗寒能力从强到弱依次是美洲椴、欧洲大叶椴、银椴、欧洲小叶椴,各品种半致死温度分别为4.3 ℃、4.7 ℃、5.5 ℃和6.0 ℃。
(2)枝条的结构指标,生理指标变化包括可溶性糖含量变化、丙二醛含量变化以及脯氨酸含量变化均可很好地反映其抗寒能力。不同生长期衡量引进椴抗寒能力的指标存在差异,相同时期种间抗寒能力也存在显著差异。
(3)随着枝条逐渐进入生长状态,生理指标的影响作用逐渐减弱,结构指标对抗寒能力的影响逐渐增强。在萌芽期,其主要调控作用的因子为可溶性糖,另外,脯氨酸与丙二醛的协同作用、枝条结构也可反映其抗寒力大小;展叶期可溶性糖的调控作用减弱,脯氨酸与丙二醛的影响作用增加,在这一时期,生理调控作用的影响程度低于枝条本身的结构指标的影响。
(4)引进椴在萌芽期的半致死温度在-15 ℃ ~-23 ℃区间,此时大连地区的最低温度不低于0 ℃,所以萌芽期树种受低温胁迫的可能性小;展叶期的半致死温度在4 ℃ ~ 6 ℃区间,在此时期应该加强对温度变化的关注,及时采取树种防寒措施。
[1]Buchner O, Neuner G. Winter frost resistance of Pinus cembra measured in situ at the alpine timberline as affected bytemperature conditions[J]. Tree Physiology, 2014, 31(11):1217-1227.
[2]黄桂华, 梁坤南, 周再知, 等. 柚木无性系苗期抗寒生理评价与选择[J].东北林业大学学报,2015, 43(9): 12-17.
[3]徐呈祥. 提高植物抗寒性的机理研究进展[J]. 生态学报,2012, 32(24): 7966-7980.
[4]Takahashi D, Li B, Nakayama T, et al. Plant plasma membrane proteomics for improving cold tolerance[J]. Frontiers in Plant Science. 2013, 4(3): 4-7.
[5]Rajashekar C B, Panda M. Water stress is a component of cold acclimation process essential for inducing full freezing tolerance in strawberry[J]. Scientia Horticulturae, 2014, 174(174): 54-59.
[6]何开跃, 李晓储, 黄利斌, 等. 3种含笑耐寒生理机制研究[J].南京林业大学学报(自然科学版), 2004, 28(4):62-64.
[7]郭海林, 高雅丹, 薛丹丹, 等. 结缕草属植物抗寒性的遗传分析[J]. 草业学报, 2009, 18(3): 53-58.
[8]Trischuk R G, Schilling B S, Low N H, et al. Cold acclimation,de-acclimation and re-acclimation of spring canola, winter canola and winter wheat: The role of carbohydrates, cold-induced stress proteins and vernalization[J]. Environmental and Experimental Botany, 2014, 106(1): 156-163.
[9]Rashid N R, Ali M E, Hamid S B, et al. A suitable method for the detection of a potential fraud of bringing macaque monkey meat into the food chain[J]. Food Additives & Contaminants Part A Chemistry Analysis Control Exposure & Risk Assessment, 2015,32(7): 1-10.
[10]焦自龙. 绿化彩化主力军“金桐力” [N].中国花卉报, 2015-12-17(4).
[11]武建勇, 薛达元, 赵富伟. 欧美植物园引种中国植物遗传资源案例研究[J]. 资源科学, 2013, 35(7): 1499-1509.
[12]胡春霞, 王 丽, 汤 杰. 低温对南果梨的生理生化指标的影响[J]. 沈阳农业大学学报, 2009, 40(3): 349-352.
[13]张友焱, 周泽福, 党宏忠,等. 毛乌素沙地油蒿不同生长期的枝条水势和叶片气体交换特性[J]. 植物资源与环境学报,2010, 19(3):17-22.
[14]郁万文. 银杏抗寒机理及种质资源抗寒性评定的初步研究[D].南京: 南京林业大学, 2008.
[15]许 瑛, 陈发棣. 菊花8个品种的低温半致死温度及其抗寒适应性[J]. 园艺学报, 2008, 35(4): 559-564.
[16]郭海林, 高雅丹, 薛丹丹, 等. 结缕草属植物抗寒性的遗传分析[J]. 草业学报, 2009, 18(3): 53-58.
[17]蔡 能,王晓明,曾慧杰,等. 3个金银花新品种逆境条件下脯氨酸含量初步分析[J]. 中南林业科技大学学报,2012,32(5):161-165.
[18]杨 惠,翟梅枝,李 丽,等. 陕西核桃栽培品种优系抗寒性评价[J]. 中南林业科技大学学报,2013,33(4):50-55.
[19]赵 广,朱万泽,谢 静,等. 不同海拔川滇高山栎丙二醛和脯氨酸含量及其影响因子[J]. 中南林业科技大学学报,2015,35(7):69-73.
[20]杨捷频. 常规石蜡切片方法的改良[J]. 生物学杂志, 2006,2(1): 45-46.
[21]李 姗. 三个树种木质部结构与耐旱性关系研究[D]. 杨凌:西北农林科技大学, 2013.
[22]郁万文. 银杏抗寒机理及种质资源抗寒性评定的初步研究[D].南京: 南京林业大学, 2008.
[23]Ershadi A, Karimi R, Mahdei K N. Freezing tolerance and its relationship with soluble carbohydrates, proline and water content in 12 grapevine cultivars[J]. Acta Physiologiae Plantarum, 2016,38(1): 1-10.
[24]王一静. 不同外源激素处理对库尔勒香梨抗寒生理变化的研究[D]. 乌鲁木齐: 新疆农业大学, 2015.
[25]Yann V, Armando L, Christian K. Theinteractionbetweenfreez ingtoleranceandphenologyintemperatedeciduoustrees[J]. Plant Science, 2014, 5(541): 541-541.
[26]Glauciana D M, Eduardo L B. Enzymatic activity in Braúna seeds subjected to thermal stress [J]. Biology, 2016, 46(6): 1044-1049.
[27]Sha fi A, Dogra V, Gill T, et al. Simultaneous over-expression of PaSOD and RaAPX in transgenic Arabidopsis thaliana confers cold stress tolerance through increase in vascular ligni fi cations.[J]. Plos One, 2014, 9(10):1-14.
[28]Macdonald M T, Lada R R, Veitch R S. Postharvest needle abscission resistance of balsam fi r is modi fi ed by harvest date[J].Canadian Journal of Forest Research, 2014, 44(11): 1394-1401.
[29]王召元, 张立莎, 常瑞丰, 等. 桃枝条组织结构与抗寒性的关系研究[J]. 河北农业科学, 2014(4): 29-33.
[30]吴晓星, 刘凤栾, 房义福, 等. 36个欧美观赏海棠品种(种)应用价值的综合评价[J]. 南京林业大学学报(自然科学版),2015, 39(1):93-98.
[31]王 娜, 王奎玲, 刘庆华, 等. 四种常绿阔叶树种的抗寒性[J].应用生态学报, 2016, 27(10): 3114-3122.
[32]王 丹, 宣继萍, 朱小晨. 假俭草抗寒性与体内碳水化合物、脯氨酸、可溶性蛋白含量的关系[J]. 草地学报, 2010, 18(6):816-822.
[33]曾炳山,黄永芳,杨懋勋,等. 柚木嫩枝扦插生根过程中营养物质的研究[J]. 中南林业科技大学学报,2013, 33(2): 1-4.
[34]张学贤. 白菜型冬油菜的抗寒性分析及抗寒性相关的分子标记[D]. 杨凌: 西北农林科技大学, 2014.
[35]王红宁, 贾林光, 邵建柱. SH40矮化中间砧苹果幼树抗寒性分析[J]. 北方园艺, 2013, 17: 16-19.
[36]Arumugam T, Rajasekaran L, et al. Vulnerability of low temperature induced needle retention in balsam fir (Abies balsamea L. ) to vapor pressure de fi cits [J]. Scandinavian Journal of Forest Research , 2015, 31(1): 1-7.
[37]陈 容, 张 丽, 曹 颖, 等. 低温胁迫下梁山慈竹再生植株叶绿素荧光特性和耐寒转录因子的表达[J]. 南京林业大学学报(自然科学版), 2014, 38(4):39-44.
Analysis on cold-resistance mechanisms of four exotic Linden species during spring in Dalian
CHEN Meiyu, XU Chengyang, LI Jinhang, YUAN Zhen
(Key Laboratory for Forest Silviculture and Conservation of Ministry of Education,Key Laboratory for Silviculture and Forest Ecosystem in Arid and Semi-arid of State Forestry Administration Beijing Forest University, Beijing 100083, China)
The objective of our research is to evaluate the cold tolerance differences and the coldresistance mechanism of four exotic Tilia species (Tilia cordata, Tilia platyphylla, Tilia america and Tilia tomentosa) in early spring. Branches at the bud bursting period and the leave outspreading period were used under the laboratory cooling test. And the relationships between LT50and physiological and anatomical characteristics were investigated. Results showed that the anti-cold regulating factors altered as the temperature changed,which was the main cause of the cold hardiness differences among the four genotypes. The regulation of the content of soluble sugar,proline and MDA was the major factor in fl uencing the cold resistance at the bud bursting period. While at the leave outspreading period,cold hardiness was mainly in fl uenced by ligni fi cation indicators and MDA relative variation. Moreover, there was a signi fi cant difference(p<0.05) in the cold hardiness among the four genotypes in both periods. Tilia tomentosa was the most resistant genotype during the bud bursting period while Tilia america performed a stronger anti-cold ability at the later period. At the bud bursting period, the chilling injury temperature of Tilia cordata, Tilia platyphylla, Tilia america and Tilia tomentosa was -15.1 ℃、-17.4 ℃、-23.0 ℃、-20.8℃ ,respectively, and 6.0 ℃、4.7 ℃、5.5 ℃ and 4.3 ℃ , respectively at the leave outspreading period. Considering the temperature situation at the two periods in Dalian, it is suggested that, there may not be freezing injury events at the bud bursting period, and measurements for preventing Linden trees from freezing or cold-induced injury should be implemented before the leave outspreading period.
North American Lindens; cold resistance; physiological index; structural index;variation analysis
S718.43;S687.1
A
1673-923X(2017)06-0024-08
10.14067/j.cnki.1673-923x.2017.06.004
2016-11-14
科学研究与研究生培养共建项目“北京典型城市森林结构合理性与木质材料高效加工利用技术”
陈美谕,硕士研究生
徐程扬,教授,博士生导师;E-mail:xyxu@bjfu.edu.cn
陈美谕,徐程扬,李金航,等. 四种引进椴在大连的春季抗寒机制分析[J].中南林业科技大学学报,2017, 37(6): 24-31.
[本文编校:吴 彬]
