光唇鱼(Acrossocheilus fasciatus)Leptin基因克隆及禁食-再投喂对其表达的影响*
2017-12-14穆方申李明云侯红红李笑萌徐玉敏
穆方申 苗 亮 李明云 侯红红 李笑萌 徐玉敏
(宁波大学 教育部应用海洋生物技术重点实验室 宁波 315211)
光唇鱼(Acrossocheilus fasciatus)Leptin基因克隆及禁食-再投喂对其表达的影响*
穆方申 苗 亮①李明云①侯红红 李笑萌 徐玉敏
(宁波大学 教育部应用海洋生物技术重点实验室 宁波 315211)
瘦素(Leptin)是在动物摄食和能量代谢调节中具有重要作用的一种蛋白。为研究光唇鱼(Acrossocheilus fasciatus)中Leptin基因(AfLep)的结构和功能, 作者克隆了其cDNA序列全长。结果显示AfLep由1315个核苷酸组成, 含1个长度为516bp的开放阅读框, 预测编码蛋白由171个氨基酸残基组成并具有长度为20aa的信号肽。系统进化树中AfLep与其他鲤科鱼类的Leptin蛋白聚为一簇, 与齐口裂腹鱼(Schizothorax prenanti)进化相关性最高。多重序列比对显示AfLep与齐口裂腹鱼Leptin蛋白相似性最高(86.7%), 与鲤科其他鱼类间的相似性均在 80%以上。AfLep具有脊椎动物Leptin蛋白的4个保守α螺旋结构, 其三级结构与人Leptin相似。AfLep在光唇鱼肝脏中的表达量最高, 其次是脑和肾, 其他组织中仅有微弱表达。实时荧光定量PCR检测显示禁食后光唇鱼肝脏AfLep表达量降低(P<0.05), 禁食1d、3d、7d时的表达量分别比与禁食前降低了58.25%、73.82%和92.05%;禁食1d和3d的鱼经再投喂后肝脏AfLep表达量均显著升高(P<0.05), 但禁食7d的鱼再投喂后AfLep表达量仅略有升高(P>0.05)。上述结果表明AfLep参与了光唇鱼的摄食管理和能量代谢调控。
光唇鱼; Leptin基因; 组织表达; 禁食-再投喂
由于天然饵料的季节性丰度变化以及人工养殖中的限制性投饵, 鱼类无论在野生还是养殖条件下都可能会经历一定的食物匮乏期。饥饿状态导致的鱼体生理生化、代谢变化会影响摄食行为、能量代谢、物质组成及生长发育、生殖等生命活动(王婷等, 2015;贺诗水等, 2016)。Leptin又称瘦素, 是在动物摄食和能量平衡的调节中有重要的作用一种蛋白质激素,并在生殖和发育、免疫、心血管功能调节等方面有重要作用(Klok, 2007)。在哺乳动物中, Leptin可通过下丘脑-垂体轴的反馈调节使动物摄食减少、能量消耗增加(Sahu, 2004)。目前已获得了斑马鱼(Danio rerio)、虹鳟(Oncorhynchus mykiss)、鲤(Cyprinus carpio)、草鱼(Ctenopharyngodon idella)、大西洋鲑(Salmo salar)等多种鱼类的Leptin基因序列, 虽然鱼类的Leptin蛋白有着相似的高级结构, 但各科、目之间氨基酸序列却有较大差异, 关于鱼类中Leptin的功能也存在争议(Copelandet al, 2011)。就摄食调节方面而言, 鲤(C.carpio)和草鱼(C.idella)均为禁食后 Leptin基因表达上调、摄食表达后下调(卢荣华等, 2015), 且对金鱼(Carassius auratus)和虹鳟(O.mykiss)侧脑室注射Leptin蛋白后可以抑制摄食、导致体重降低(de Pedroet al, 2006; Aguilaret al, 2010); 但在点带石斑鱼(Epinephelus coioides)、虹鳟(O.mykiss)中则发现饥饿状态下Leptin基因表达上调(Klinget al, 2009; Zhanget al, 2013)。鱼类中Leptin的功能可能比哺乳动物复杂,要弄清 Leptin在鱼类摄食管理和能量代谢调控中的功能及作用机制还需要进行更加广泛和深入的研究。
光唇鱼(Acrossocheilus fasciatus)俗称淡水石斑鱼,属鲤形目、 鲤科、鲃亚科、光唇鱼属, 不但有较高的营养价值, 还可作为小型观赏鱼(潘娜等, 2015)。近年来光唇鱼的人工繁育和养殖技术逐渐成熟, 养殖规模不断扩大, 但在摄食调控、能量代谢调节等方面鲜有研究报道。本研究克隆光唇鱼Leptin基因(AfLep),并检测禁食期间及恢复投喂后AfLep的表达变化, 以期为研究鱼类 Leptin的功能及其光唇鱼的摄食和能量代谢调节中的作用提供基础资料。
1 材料与方法
1.1 实验动物及禁食-再投喂实验
光唇鱼由浙江省新昌县新渔水产养殖专业合作社提供, 为人工繁育、养殖的12月龄光唇鱼, 共450尾, 平均体重(3.13±1.14)g、平均体长(6.82±0.64)cm。将鱼随机分配到9个容积为50L的塑料桶中, 每桶50尾, 暂养4天后开始实验。暂养期间每天投喂2次通威牌颗粒饲料, 每次投喂量为鱼体重的 2%。每 3个桶为1组, 处理如下: 第1组禁食1d后恢复投喂, 第2组禁食3d后恢复投喂, 第3组禁食7d后恢复投喂。
1.2 样品采集
在禁食前(0d)和禁食后 1d、3d、7d以及再投喂后3h分别采样1次, 每次每桶随机取样5尾。取样时将鱼用丁香酚麻醉后立即解剖, 其中禁食前取肌肉、鳃、心、肝脏、脾、肠、肾、脑等组织, 用于AfLep基因克隆、组织表达分析和表达量检测; 禁食及再投喂后取肝脏, 用于检测AfLep基因表达变化。样品经液氮速冻后–80℃保存, 用 Trizol法提取各组织总RNA, 反转录合成第一链cDNA。
1.3 AfLep基因克隆
检索GenBank数据库中鲤科鱼类Leptin蛋白序列, 经序列比对后根据保守区域设计 3对兼并引物(lep1, lep2和lep3, 见表1)对光唇鱼AfLep基因cDNA序列进行扩增。用 ExTaq®Hot Start Version聚合酶体系(TaKaRa)进行PCR扩增, 50μL反应体系包括: 5U/μL的ExTaqHS 0.25μL, 10×ExTaqBuffer (Mg2+Plus) 5μL,dNTP Mixutre 4μL, 上、下游引物各0.5μL, cDNA模板1μL, dH2O 38.75μL。反应程序为: 94℃预变性5min;94℃变性 30s、退火(引物退火温度见表 1)30s、72℃合成30s, 循环30次; 72℃延伸10min。PCR产物经1.5%琼脂糖凝胶电泳检测后连接入 pMD-19T载体,转化入感受态大肠杆菌, 挑选阳性克隆菌落送测序。引物合成和测序均由生工生物工程(上海)股份有限公司进行。对测得的序列进行拼接后获得光唇鱼AfLep基因核心序列。
根据所获核心序列设计引物 lep-3′-RACE和lep-5′-RACE(表 1), 用 3′-Full RACE Core Set Ver.2.0和5′-Full RACE Kit (TaKaRa)扩增光唇鱼AfLep基因cDNA的3′端和5′端序列, 扩增体系及反应程序均按照说明书进行。扩增产物经 1.5%琼脂糖凝胶电泳检测后连接入pMD-19T载体, 转化入感受态大肠杆菌,挑选阳性克隆菌落送测序, 拼接后获得光唇鱼AfLep基因cDNA序列全长。

表1 引物信息Tab.1 Information of the primers
1.4 AfLep序列分析及系统进化树构建
对获得的AfLep基因cDNA序列, 用在线ORF Finder工具(https://www.ncbi.nlm.nih.gov/orffinder/)分析开放读码框(open reading frame, ORF); 用SingalP 3.0 (http://www.cbs.dtu.dk/services/SignalP)预测信号肽; 用 SWISS-MODEL (http://swissmodel.expasy.org//SWISS-MODEL.html)的 ProMod II功能对 AfLep蛋白进行二级和三级结构预测(以蛋白质数据库中的人 Leptin结构作为参照); 从 GenBank数据库中检索鱼类及其他脊椎动物的 Leptin序列(表 2), 经 Cluastal X1.83软件进行多重比对后用MEGA5.0软件构建基于氨基酸序列的 NJ法(Neighbor-joining)系统进化树, 设置 bootstraps验证次数为1000次。
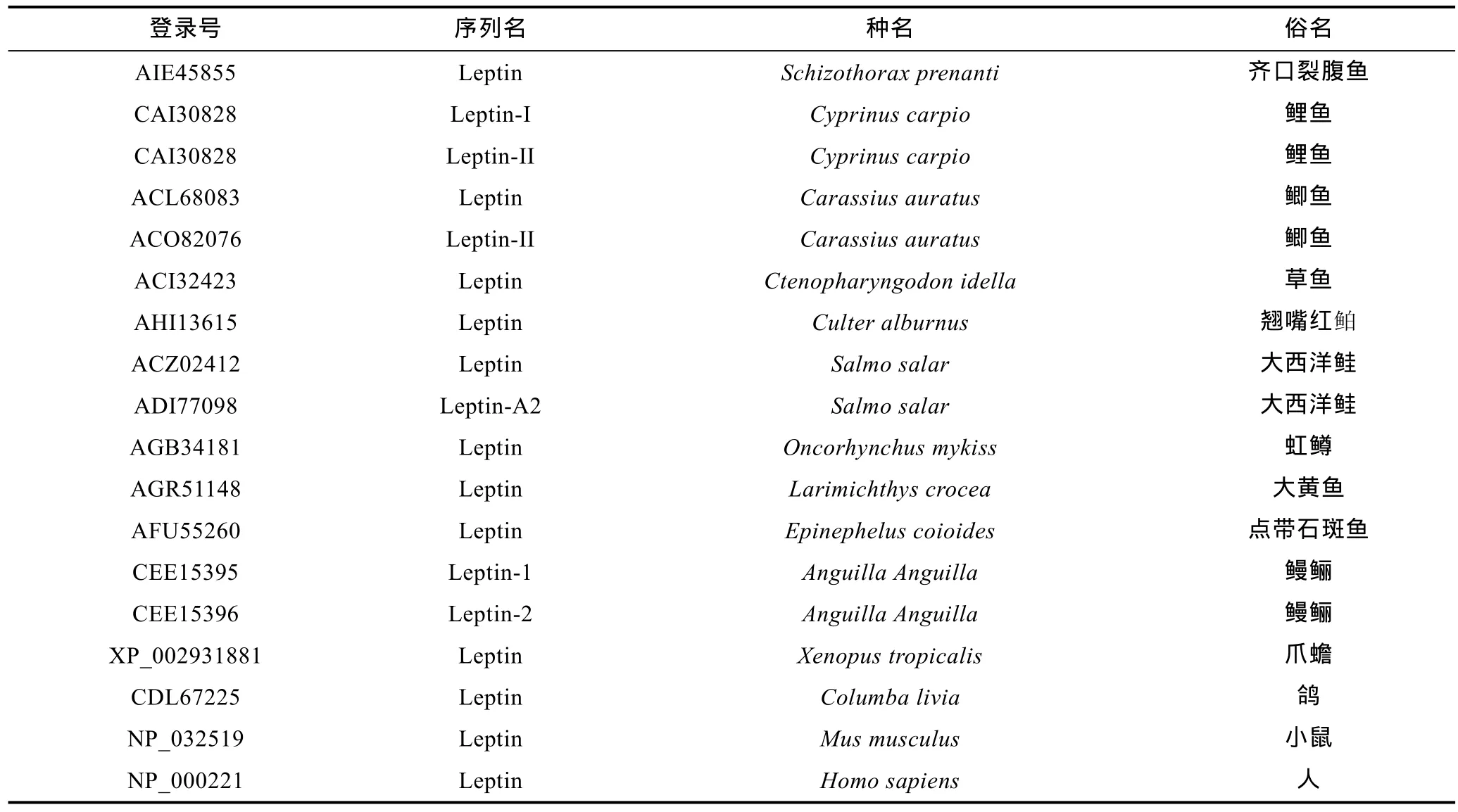
表2 多重序列比对及构建系统进化树所用序列Tab.2 Sequences that used for multiple alignment and phylogenetic tree construction
1.5 AfLep组织表达及禁食-再投喂后表达变化
根据获得的AfLep基因cDNA序列设计1对表达检测引物q-lep、并以β-actin作为内参(表1), 检测AfLep基因在光唇鱼各组织中的表达情况及禁食-再投喂后的表达变化。
以提取的正常投喂健康光唇鱼各组织 cDNA为模板, 通过半定量RT-PCR检测各组织中AfLep基因的表达情况。20μL反应体系包括: 2×TaqPremix-Dye酶 10μL, 正反向引物各 1μL, 模板 cDNA 1μL, dH2O 7μL。程序为: 94℃预变性 5min: 94℃变性30s、58℃退火30s、72℃延伸30s, 循环30次; 72℃延伸10min。PCR产物用1.5%琼脂糖凝胶电泳检测后凝胶成像。
对禁食期间及再投喂后光唇鱼肝脏中 AfLep基因的表达变化进行实时荧光定量 PCR (real-time quantitative PCR, RT-qPCR)检测。反应使用 SYBR®PremixEx TaqTM试剂盒(TaKaRa), 20μL反应体系包括: SYBR®Premix ExTaq(Tli RNaseH Plus) 10μL,正、反向引物各1μL, cDNA模板 1μL, ddH2O 7μL。反应程序为: 94℃预变性2min; 94℃变性15s、58℃退火15s、72℃延伸30s, 循环40次并记录CT值。反应完成后, 系统在 65—94℃、每次升温 0.5℃生成溶解曲线。以β-actin 为内参、用 2–ΔΔCt法(Livaket al, 2001)计算AfLep基因相对表达量。用Excel2007软件对数据进行统计, 计算平均数和标准差; 用 SPSS16.0软件对不同时间点的基因相对表达量进行单因素方差(One way ANOVA)分析, 以P<0.05为具有显著性差异。
2 结果
2.1 AfLep序列特征
经测序和拼接, 获得 AfLep基因序列全长1315bp, 其中 5′端 175bp, 3′端 699bp; 该序列含有 1个长度为516bp的开放阅读框(ORF), 编码171个氨基酸; 经SignalP4.1预测, 编码蛋白有长度为20aa的信号肽(图1)。
基于各种动物 Leptin氨基酸序列构建的系统进化树中(图 2), 鱼类 Leptin独立于两栖类、鸟类、哺乳类等其他脊椎动物而聚为一个大簇; 在鱼类Leptin中鲤科、鳗鲡科、石首鱼科、鲑科鱼类均单独聚为一个小簇; AfLep位于鲤科鱼类的分支中, 优先与齐口裂腹鱼 Leptin相聚。多重序列比对显示AfLep与齐口裂腹鱼 Leptin相似性最高(86.7%), 与鲤科其他鱼类 Leptin的相似性均在 80%以上, 与非鲤科鱼类Leptin的相似性较低(30%左右), 而与两栖类、鸟类和哺乳类等其他脊椎动物Leptin相似性仅约20%。

图1 AfLep基因cDNA核苷酸序列及预测编码的氨基酸序列Fig.1 The nucleotide sequences of AfLep cDNA and deduced amino acid sequence
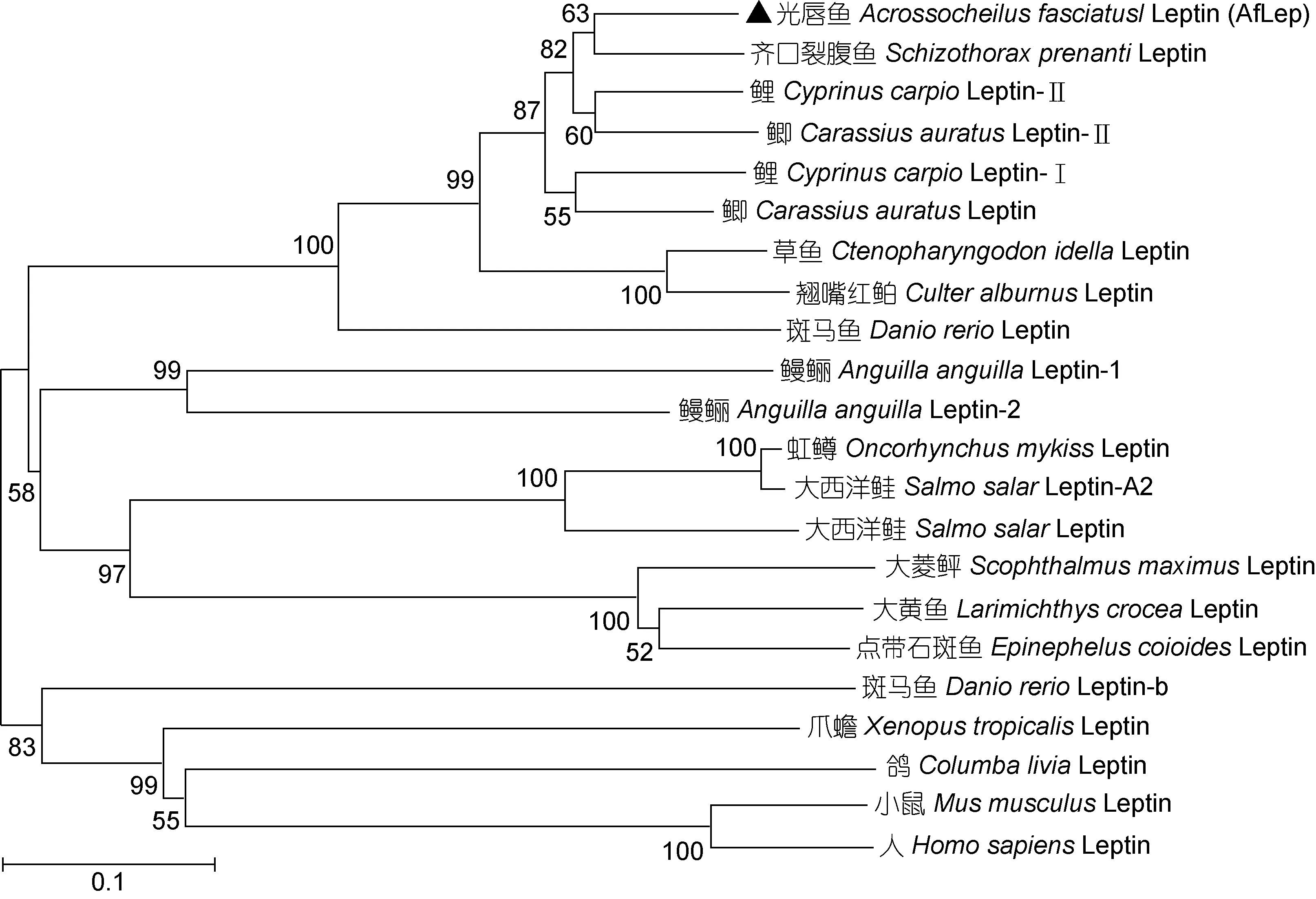
图2 Leptin系统进化树(NJ法, bootstraps=1000)Fig.2 The phylogenetic tree of Leptin (neighbor-joining method, bootstraps=1000)
序列比对(图 3)显示光唇鱼、齐口裂腹鱼、鲤等鲤科鱼类的 Leptin氨基酸序列较为保守, 而与非鲤科鱼类以及其他脊椎动物的Leptin序列差异较大; 但各种动物Leptin蛋白的二级结构高度保守, 均有4个α螺旋(Helix A-D), 且具有长度约 20aa的信号肽。预测的AfLep蛋白三级空间结构也与人Leptin蛋白相似(图4)。

图3 AfLep与其他生物Leptin氨基酸序列比对Fig.3 Amino acid sequence alignment among AfLep and Leptin of other species

图4 Leptin蛋白三级结构Fig.4 The tertiary structures of Leptin protein
2.2 组织表达特征
半定量RT-PCR检测显示了AfLep基因在光唇鱼肝脏中的表达量最高, 其次是脑和肾, 在肌肉、脾、心脏、鳃和肠等组织中仅有微弱表达(图5)。
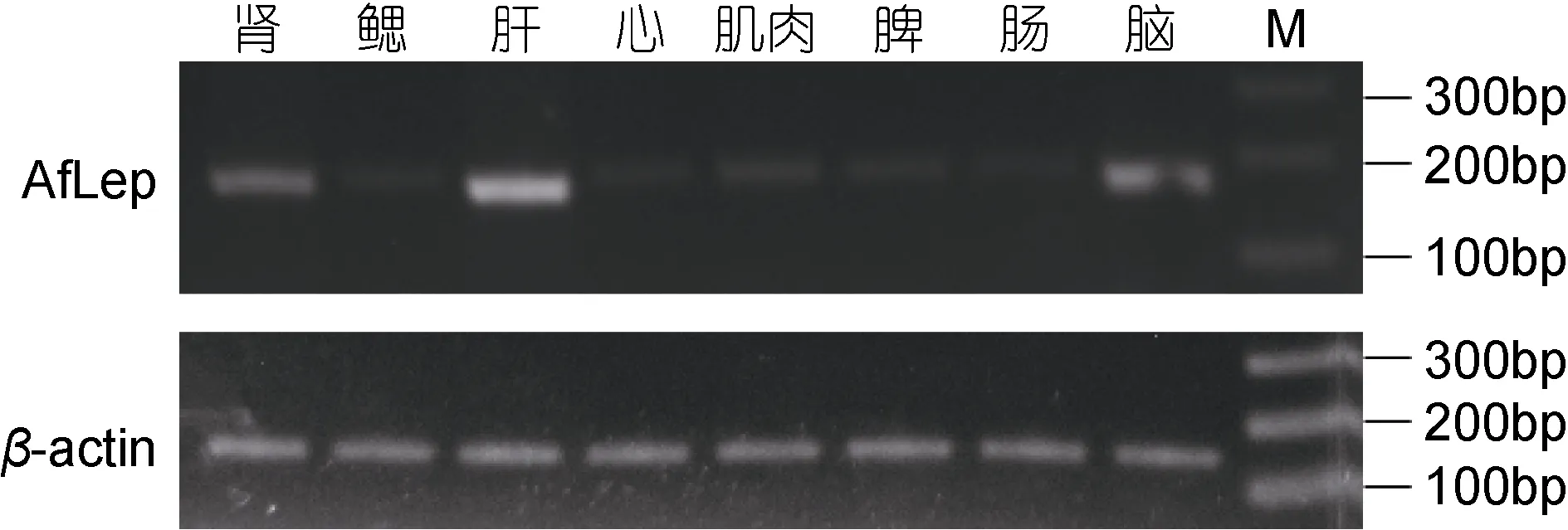
图5 AfLep基因在光唇鱼各组织中的表达Fig.5 The expression of AfLep gene in various tissues of A.fasciatus
2.3 禁食及再投喂后光唇鱼肝脏AfLep基因的表达变化
经实时荧光定量PCR检测(图6), 禁食后光唇鱼肝脏中 AfLep基因表达显著下调(P<0.05), 且表达水平随禁食时间延长而逐渐降低: 禁食 1d、3d、7d后AfLep基因表达量分别比禁食前(0d)降低了58.25%、73.82%和92.05%。禁食-再投喂后AfLep基因表达上调, 其中禁食1d组和禁食3d组分别显著升高了1.79倍和2.65倍(P<0.05), 而禁食7d组的表达水平仅略有升高、差异不显著(P>0.05)。
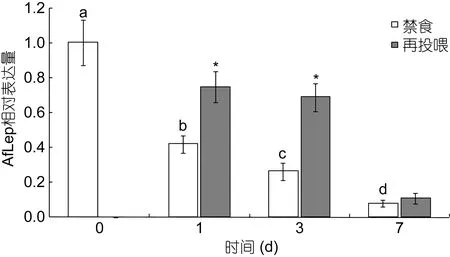
图6 禁食及再投喂后光唇鱼肝脏中AfLep基因表达变化Fig.6 Changes in expression of AfLep in liver of A.fasciatus during fasting and after refeeding
3 讨论
自Zhang等(1994)率先在小鼠中得到Leptin基因以来, 已在包括鱼类在内的脊椎动物各类群中都发现有与哺乳类Leptin同源的基因存在。在哺乳动物及非洲爪蟾的基因组中Leptin基因仅有1个拷贝、编码1种蛋白, 而在某些鱼类中则发现存在多个拷贝以及不同的Leptin蛋白亚型(Kurokawaet al, 2009; Angotziet al, 2013; 卢荣华等, 2015), 这可能与鱼类在进化过程中发生过基因组复制有关(周莉等, 2006)。另外,鱼类中Leptin基因的起源进化关系也比较复杂, 例如鲤(C.carpio)2个Leptin基因拷贝所编码的蛋白相似性较高(84%), 而斑马鱼(D.rerio)中2个Leptin蛋白亚型的相似性却仅有 24%(卢荣华等, 2015)。本研究通过同源克隆从光唇鱼中获得了一个 Leptin基因(AfLep), 但不能确定光唇鱼基因组中是否还存在该基因的其他拷贝。
在本研究构建的Leptin系统进化树中, 包括光唇鱼AfLep在内的各种鲤科鱼类Leptin聚为一簇, 其氨基酸序列相似性在80%以上, 而与非鲤科鱼类Leptin的相似性仅 30%左右。鱼类不同科、目之间 Leptin氨基酸序列有较大差异, 这可能与鱼类种系发生和系统进化关系复杂有关; 但研究显示在各种脊椎动物基因组中Leptin基因有着相似的线性排列, 表明它们可能有相同的起源; 并且各类动物Leptin蛋白二级结构高度保守, 均含2个半胱氨酸和形成4个α螺旋(卢荣华等, 2015; 张沛等, 2016); 本研究预测的光唇鱼AfLep蛋白的空间结构也与人类Leptin蛋白相似。结构上的保守可能对 Leptin蛋白的稳定性和活性有重要影响, 而氨基酸序列的低相似性则反映了Leptin基因在物种进化中的差异, 这种差异可能导致该基因在不同生物中功能的多样化。
小鼠等哺乳动物中 Leptin基因主要在脂肪组织表达(Trayhurnet al, 1995), 两栖类Leptin则是脑和心脏中表达量较高。在光唇鱼肝脏中AfLep基因高表达,斑马鱼(D.rerio)、花鲈(Lateolabrax maculatus)、鳜鱼(Siniperca chuatsi)、点带石斑鱼(E.coioide)等鱼类中也是肝脏中 Leptin基因表达量最高(Gorissenet al,2009; Zhanget al, 2013; 张沛等, 2016; Yuanet al,2016); 但也有些鱼类在其他组织中出现 Leptin的高表达, 如团头鲂(Megalobrama amblycephala)肾脏、鳃、肠、性腺中 Leptin的表达量远均高于肝脏(赵鸿昊等, 2016), 大西洋鲑(S.salar)Leptin则是在脑中表达量最高、肌肉中也有较高表达(Ronnestadet al,2010); Leptin基因组织表达的这种差异性提示该基因在鱼类中可能有多种功能, 并且在不同鱼类中的功能可能有较大差异。
在人、小鼠等哺乳动物中的实验显示饥饿后Leptin基因表达量和血浆Leptin蛋白水平显著降低、进食后升高(Kolaczynskiet al, 1995; Saladinet al,1995; Trayhurnet al, 1995; Weigleet al, 1997)。本研究中 AfLep在光唇鱼肝脏中的表达也表现为禁食后显著降低、恢复喂食后升高, 提示该基因有降低食欲的作用, 在草鱼(C.idella)、金鱼(C.auratus)、鲈鱼(Morone saxatilis)、虹鳟(O.mykiss)等鱼类中也得到了相似的结果(卢荣华等, 2015), 但在点带石斑鱼(E.coioides)和虹鳟(O.mykiss)中则发现禁食后Leptin基因表达上调、血浆Leptin蛋白含量升高(Klinget al,2009; Zhanget al, 2013)。这可能与不同鱼类在摄食和糖、脂代谢管理方式上有着很大的差异有关, 例如通常情况下机体处于饥饿状态时会分解脂肪提供能量,但真鲷(Pagrus major)幼鱼经过短期饥饿后却出现了脂肪含量的升高(Kaneko, 2016); 而肉食性和植食性鱼类在糖代谢上也有很大不同(Moon, 2001)。另外,本研究中禁食1d和3d的光唇鱼在摄食后肝脏AfLep基因表达量均显著升高, 提示此时鱼体的能量储备可能消耗不大; 而禁食7d组摄食后肝脏AfLep基因表达仍处于较低水平, 提示鱼体脂肪消耗过多、需摄取更多食物进行补充。贺诗水等(2016)也发现鲫鱼(Carassius auratus)饥饿 4d后脂肪含量仅下降了约10%, 而饥饿6d后已减少了50%以上。虽然本研究表明AfLep基因参与了光唇鱼的摄食管理, 但要弄清其作用机制以及在能量代谢中的功能仍需进一步研究。
王 婷, 刘利平, 陈桃英等, 2015.鳗鲡(Anguilla japonica)性腺发育和饥饿胁迫下生物学指标及体内蛋白质与氨基酸含量变化.海洋与湖沼, 46(6): 1373—1379
卢荣华, 孙君君, 梁旭方等, 2015.鱼类leptin的生物学特性及功能.水生生物学报, 39(3): 583—589
张 沛, 温海深, 迟美丽等, 2016.花鲈瘦素基因的克隆及低盐度条件下调控表达分析.大连海洋大学学报, 31(1):13—18
周 莉, 汪 洋, 桂建芳, 2006.鱼类特异的基因组复制.动物学研究, 27(5): 525—532
赵鸿昊, 李学华, 曾 聪等, 2016.leptin基因在团头鲂成鱼各组织和早期发育中的表达分析.华中农业大学学报, 35(1):92—98
贺诗水, 王洪凯, 蒋万祥等, 2016.上市前短期饥饿对鲫鱼肌肉品质的影响.食品工业科技, 37(1): 334—337
潘 娜, 苗 亮, 李明云等, 2015.浙江新昌光唇鱼(Acrossocheilus)COII和D-loop基因克隆及系统发育分析.海洋与湖沼, 46(2): 381—388
Aguilar A J, Conde-Sieira M, Polakof Set al, 2010.Central leptin treatment modulates brain glucosensing function and peripheral energy metabolism of rainbow trout.Peptides,31(6): 1044—1054
Angotzi A R, Stefansson S O, Nilsen T Oet al, 2013.Molecular cloning and genomic characterization of novel leptin-like genes in salmonids provide new insight into the evolution of the leptin gene family.General and Comparative Endocrinology, 187: 48—59
Copeland D L, Duff R J, Liu Qet al, 2011.Leptin in teleost fishes: an argument for comparative study.Frontiers in Physiology, 2: 26
de Pedro N, Martínez-Álvarez R, Delgado M J, 2006.Acute and chronic leptin reduces food intake and body weight in goldfish (Carassius auratus).Journal of Endocrinology,188(3): 513—520
Gorissen M, Bernier N J, Nabuurs S Bet al, 2009.Two divergent leptin paralogues in zebrafish (Danio rerio) that originate early in teleostean evolution.Journal of Endocrinology,201(3): 329—339
Kaneko G, Shirakami H, Yamada Tet al, 2016.Short-term fasting increases skeletal muscle lipid content in association with enhanced mRNA levels of lipoprotein lipase 1 in lean juvenile red seabream (Pagrus major).Aquaculture, 452:160—168
Kling P, Rønnestad I, Stefansson S Oet al, 2009.A homologous salmonid leptin radioimmunoassay indicates elevated plasma leptin levels during fasting of rainbow trout.General and Comparative Endocrinology, 162(3): 307—312
Klok M D, Jakobsdottir S, Drent M L, 2007.The role of leptin and ghrelin in the regulation of food intake and body weight in humans: a review.Obesity Reviews, 8(1): 21—34
Kolaczynski J W, Considine R V, Ohannesian Jet al, 1995.Responses of leptin to short-term fasting and refeeding in humans: a link with ketogenesis but not ketones themselves.Diabetes, 45(11): 1511—1515
Kurokawa T, Murashita K, 2009.Genomic characterization of multiple leptin genes and a leptin receptor gene in the Japanese Medaka,Oryzias latipes.General and Comparative Endocrinology, 161(2): 229—237
Livak K J, Schmittgen T D, 2001.Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCTmethod.Methods, 25(4): 402—408
Moon T W, 2001.Glucose intolerance in teleost fish: fact or fiction? Comparative Biochemistry and Physiology Part B:Biochemistry and Molecular Biology, 129(2—3): 243—249
Ronnestad I, Nilsen T O, Murashita Ket al, 2010.Leptin and leptin receptor genes in Atlantic salmon: cloning, phylogeny,tissue distribution and expression correlated to long-term feeding status.General and Comparative Endocrinology,168(1): 55—70
Sahu A, 2004.Minireview: a hypothalamic role in energy balance with special emphasis on leptin.Endocrinology, 145(6):2613—2620
Saladin R, De Vos P, Guerre-Millo Met al, 1995.Transient increase inobesegene expression after food intake or insulin administration.Nature, 377(6549): 527—528
Trayhurn P, Thomas M E A, Duncan J Set al, 1995.Effects of fasting and refeeding onobgene expression in white adipose tissue of lean and obese (ob/ob) mice.FEBS Letters, 368(3):488—490
Weigle D S, Duell P B, Connor W Eet al, 1997.Effect of fasting,refeeding, and dietary fat restriction on plasma leptin levels.The Journal of Clinical Endocrinology and Metabolism,82(2): 561—565
Yuan X C, Li A X, Liang X Fet al, 2016.Leptin expression in mandarin fishSiniperca chuatsi(Basilewsky): regulation by postprandial and short-term fasting treatment.Comparative Biochemistry and Physiology Part A: Molecular &Integrative Physiology, 194: 8—18
Zhang H X, Chen H P, Zhang Yet al, 2013.Molecular cloning,characterization and expression profiles of multiple leptin genes and a leptin receptor gene in orange-spotted grouper(Epinephelus coioides).General and Comparative Endocrinology, 181: 295—305
Zhang Y Y, Proenca R, Maffei Met al, 1994.Positional cloning of the mouseobesegene and its human homologue.Nature,372(6505): 425—432
CLONING AND EXPRESSION OF LEPTIN GENE INACROSSOCHEILUS FASCIATUSDURING FASTING AND REFEEDING
MU Fang-Shen, MIAO Liang, LI Ming-Yun, HOU Hong-Hong, LI Xiao-Meng, XU Yu-Min
(Key Laboratory of Applied Marine Biotechnology,Ministry of Education,Ningbo University,Ningbo315211,China)
Leptin plays an important role in regulation of food intake and energy expenditure in animals.To study the structure and function of Leptin gene in commercial fishAcrossocheilus fasciatus, we cloned the cDNA sequence of Leptin gene inA.fasciatus, named AfLep.The full-length of AfLep cDNA sequence was 1315bp, contained an open reading frame(ORF) of 516bp encoding 171 amino acids; and the initiative 20 amino acid consisted of signal peptide.In the phylogenetic tree, AfLep gathered with Leptin of other Cyprinidae fish and was most closely related to Leptin ofSchizothorax prenanti.The multiple sequence alignment showed that AfLep shared the highest amino acid sequence identify (86.7%) with Leptin ofS.prenanti, and had more than 80% sequence identity to other Cyprinidae fish.The predicted protein of AfLep has four α helices regions, which is conserved in vertebrates.The predicted tertiary structure of AfLep is similar to human Leptin.AfLep gene had the highest expression level in liver, followed by brain and kidney, and weakly expressed in other tissues.Real-time fluorescent quantitative PCR (RT-qPCR) showed that the expression of AfLep decreased significantly in liver after being fasted (P<0.05), and reduced by 58.25%, 73.82%, and 92.05% after fasting 1d, 3d, and 7d, respectively,compared to pre-fasting.When the fish were refed 1d and 3d after fasting, the expression level of AfLep in liver significantly increased (P<0.05), but in the group of fasting 7d the AfLep expression was slightly elevated after refeeding(P>0.05).All these results indicate that AfLep participates in the adjustment of feeding regulation and energy metabolism.
Acrossocheilus fasciatus; Leptin gene; tissue expression; fasting and refeeding
Q346
10.11693/hyhz20170100023
* 浙江省海洋与渔业项目, 浙海渔计[2012]83号; 国家星火计划项目, 2011GA701001号。穆方申, E-mail: mufangshen@126.com
① 通讯作者: 苗 亮, 博士, 硕士生导师, E-mail: miaoliang@nbu.edu.cn; 李明云, 教授, 博士生导师, E-mail:limingyun@nbu.edu.cn
2017-01-26, 收修改稿日期: 2017-03-03
猜你喜欢
杂志排行

海洋与湖沼的其它文章
- 瓦氏黄颡鱼(Pelteobagrus vachellii)脂肪酸去饱和酶FAD2和延伸酶ELOVL5的克隆及表达分析*
- 基于de novo高通量测序的曼氏无针乌贼(Sepiellajaponica)ESTs中微卫星位点筛选与特征分析*
- 泥蚶(Tegillarca granosa)重组铁蛋白富集重金属离子的特性及化学传感器的研究*
- 厚壳贻贝(Mytilus coruscus)金属硫蛋白MT-10:cDNA克隆、结构分析及铜离子胁迫下的表达*
- 舟山海洋沉积物及沿海沉积物流动相石油污染的电阻率特性及其影响因素研究*
- 迟缓型爱德华氏菌(Edwardsiella tarda)诱导牙鲆(Paralichthys olivaceus)TLR1及TLR2基因的表达分析*