小麦品种东农冬麦2号根中 TaEXPA7部分同源基因的克隆及表达特性分析
2017-12-11董佳敏徐永清彭丽娜姚树宽赵巧芩李凤兰胡宝忠
董佳敏,徐永清,彭丽娜,冯 旭,姚树宽,赵巧芩,李凤兰,胡宝忠,2
(1.东北农业大学生命科学学院,黑龙江哈尔滨 150030; 2.哈尔滨学院,黑龙江哈尔滨 150086)
小麦品种东农冬麦2号根中 TaEXPA7部分同源基因的克隆及表达特性分析
董佳敏1,徐永清1,彭丽娜1,冯 旭1,姚树宽1,赵巧芩1,李凤兰1,胡宝忠1,2
(1.东北农业大学生命科学学院,黑龙江哈尔滨 150030; 2.哈尔滨学院,黑龙江哈尔滨 150086)
膨胀素(expansin)是植物生长发育过程中诱导细胞壁松弛和伸展的蛋白,包括EXPA、EXPB、EXLA、EXLB四个基因家族,对植物根系的形成及快速发育具有重要作用。本试验以寒地冬小麦品种东农冬麦2号两叶一心期的根为材料,克隆了 TaEXPA7基因的3个CDS全长,其氨基酸序列同源性较高,仅信号肽处有两个氨基酸不同,其核酸序列长度均为777 bp,编码258个氨基酸,分别命名为 TaEXPA7-A、 TaEXPA7-B、 TaEXPA7-D,且分别定位于2AL、2BL、2DL染色体。蛋白均含有DPBB-1和Pollen-allerg-1两个保守结构域,分子量为27 670.74 Da,等电点为8.09,均为疏水性蛋白;与节节麦、二穗短柄草的相似性分别为99%和92%。采用两叶一心期的根进行qRT-PCR分析发现, TaEXPA7-A/B/D三个基因在根尖中的表达量均较高,伸长区和成熟区次之;对冬小麦根进行低温、干旱和激素处理后,这三个基因均下调表达,并且 TaEXPA7-B基因的相对表达量较高, TaEXPA7-D的相对表达量较低。由此可见,多倍体小麦不同染色体上的基因在应对不同环境胁迫时,表达以及应答模式存在差异,并且在不同的环境胁迫下,发挥主导作用的染色体也存在差异;此外,该基因在根中的表达可能与低温、干旱以及外源激素的胁迫有一定的关系,可能是促进根系生长的重要基因。
冬小麦; TaEXPA7;部分同源;表达特性
膨胀素(expansin)是植物生长发育过程中诱导细胞壁松驰和伸展的蛋白,于1989年首次被发现。1992年,McQueen-Mason等[1]从黄瓜中分离纯化出这种蛋白,并发现其能调节幼苗下胚轴的伸长,第一次将其命名为expansin。在植物中,expansin可以分为四个基因家族α-expansin(EXPA)、β-expansin(EXPB)、expansin-likeA(EXLA)以及expansin-likeB(EXLB),由信号肽、催化区和结合区三部分构成[2]。
Hu等[3]分别克隆了小麦 TaEXPA1-A/B/D三个基因,并将其转入拟南芥,发现其具有相似的功能。然而,小麦染色体组的复杂多倍体特性使得对其遗传、表达特性及其功能的分析极具挑战性。研究发现, OsEXPA1、 OsEXPA2、 OsEXPA3、 OsEXPA4、OsEXPB2和 OsEXPA17等基因均在水稻根尖特异性表达,参与根尖形态建成,促进根系生长发育[4-7]。近年来,国内外关于膨胀素基因在各种非生物胁迫下的表达特性被广泛研究,如干旱、低温和激素会影响膨胀素基因的表达,从而影响植物生长。Che等[8]研究发现,Al离子诱导后, OsEXPA10基因参与水稻的根细胞伸长。韩阳阳等[9]研究发现,GA3、ET和IAA显著抑制了 TaEXPB23的表达水平,而MeJA则上调了该基因的表达,IAA处理先抑制随后诱导了 TaEXPB23的表达,而ABA则在后期稍微抑制了 TaEXPB23的表达;此外,盐胁迫能够明显提高 TaEXPB23的mRNA表达水平,而高温则抑制该基因的表达。
目前,在小麦中关于膨胀素和根之间关系的研究鲜有报道,复杂的小麦各个基因组之间的关系以及基因的表达差异关系也尚未明确。因此,本研究以寒地高产冬小麦品种东农冬麦2号为材料,同源克隆了 TaEXPA7-A/B/D三个基因;通过生物信息学分析了解该基因基本的生物学功能;利用qRT-PCR技术,研究了这三个基因在小麦幼苗期根中不同部位及在低温、干旱和激素胁迫下的表达特性及表达差异,以期为进一步研究膨胀素在根系形态建成中的作用提供理论依据。
1 材料与方法
1.1 试验材料及处理
试验材料为高抗寒的冬小麦品种东农冬麦2号,由东北农业大学农学院小麦育种研究室提供。
挑选颗粒饱满的东农冬麦2号种子,用0.1% HgCl2溶液消毒处理30 s,蒸馏水冲洗6 h,室温浸种24 h,然后将种子平铺于含有双层滤纸的培养皿中培养,露白后移栽至苗钵,置于培养室中培养[9]。待根长至大约30 cm时对根尖、伸长区和成熟区进行取样,每个部位均单株取材,随机取三株。以水培为对照组,对4 ℃、聚乙二醇(PEG)、脱落酸(ABA)、茉莉酸甲脂(MeJA)及水杨酸(SA)分别处理2 h、6 h、12 h、24 h和48 h的根进行取材,均单株取材,随机取三株,取样后均迅速放入液氮中速冻,然后置于-80 ℃超低温冰箱保存备用。
RNA提取试剂盒TransZol、RT-PCR所用荧光染料SYBR Green、Top Taq酶、 Peasy-T3克隆载体、质粒提取试剂盒、DNA回收试剂盒、反转录试剂盒均购自全式金生物科技有限公司;试验所有引物合成和测序在哈尔滨博仕生物进行;试验所需大肠杆菌 DH5α、ABA、茉莉酸甲酯、水杨酸、聚乙二醇、LB肉汤、LB营养琼脂、氯仿、甘油等生化试剂均购自凯誉生物科技有限公司。使用Primer 5.0软件进行引物设计;使用Excel和GraphPad Prism软件完成作图和数据处理。
1.2 RNA的提取及cDNA的合成
使用TransZol试剂盒(ER501)进行总RNA的提取,-80 ℃保存备用。使用UV-240紫外分光光度计检测RNA浓度,OD260/OD280=1.8~2.1,且OD260/OD230>2.0,浓度不小于500 ng·μL-1,使用琼脂糖凝胶电泳检测总RNA的完整性。以Oligo(DT)为引物,将所得RNA反转录为cDNA,-20 ℃保存,并用β-actin检测反转录效果。
1.3 TaEXPA7-A/B/D基因的克隆及序列分析
组织特异性表达分析表明, TaEXPA7基因在根中的相对表达量较高,且东农冬麦2号的RNA-seq结果显示,该基因的整体表达量较高,可能与根系生长有重要关系。在NCBI的GenBank中查询 TaEXPA7三个同源基因的CDS序列,对这三个基因的相似部分利用Primer 5.0设计特异性引物(表1)。以冬小麦幼苗期根的cDNA为模板进行PCR扩增,电泳回收后,将回收产物连接到Peasy-T3载体上,转入DH5α感受态细胞,用含Amp(100 μg·mL-1)的LB平板筛选目的菌落,随机挑取30个单菌落,采取菌液PCR的方法进行目的基因扩增,将出现目标基因的克隆送公司测序。利用NCBI ORF Find分析CDS序列的ORF,并用Primer 5.0软件预测编码的氨基酸序列,对获得的序列用BLAST程序搜索GenBank中与 TaEXPA7-A/B/D基因同源的氨基酸序列,采用DNAMAN 6.0和MEGA 6.0软件进行氨基酸序列比对和系统进化树的构建。
1.4 TaEXPA7-A/B/D基因的表达特性分析
在NCBI的GenBank中查询 TaEXPA7-A/B/D的CDS差异序列并参照此差异序列分别设计特异性qRT-PCR引物(表1)。对1.2中得到的各处理cDNA进行梯度稀释,根据Ct值确定最佳反应浓度。以确定好最佳浓度的cDNA为模板,以小麦肌动蛋白基因β-actin为内参,采用qRT-PCR进行扩增、分析,生物学重复和技术重复各3次。反应体系(20 μL)如下:2×Trans Star Top Green Qpcr Super Mix 10 μL,上下游引物各0.4 μL,cDNA 1.0 μL,ddH2O 8.2 μL。反应在实时荧光定量PCR仪(FQD-9620)上进行。采用2-ΔΔCt法计算相对表达量,采用DPS 7.05 数据处理系统软件进行方差及显著性分析。

表1 引物信息Table 1 Information of primers used in this study
2 结果与分析
2.1 RNA的检测结果
以电压150 V,电流120 mA,1%的琼脂糖凝胶电泳检测所提取的RNA(图1),可以看出,28S和18S两条谱带清晰且前者的亮度大于后者,说明提取的总RNA质量较好。
2.2 TaEXPA7-A/B/D基因CDS的克隆及序列分析
为了获得 TaEXPA7的部分同源基因,根据 TaEXPA7-A/B/D三个基因的ORF(Open Reading Frame,开放阅读框)序列(GenBank登录号:AA0307290.1、AA0383180.1、AA0559500.1)的相似部分,设计了两条特异引物( TaEXPA7-F,TaEXPA7-R),进行了PCR,经琼脂糖凝胶电泳后获得了一条800 bp左右的条带(图2)。将携带该目的条带的克隆挑取30个进行测序,获得了3条分别定位于2AL、2BL和2DL染色体上的CDS序列,其长度均为777 bp,依次命名为: TaEXPA7-A、TaEXPA7-B、TaEXPA7-D。生物信息学分析发现,3个基因均编码258个氨基酸,分子量为27 670.74 Da,等电点为8.09,均为疏水性蛋白,包括DPBB-1和Pollen-allerg-1两个保守结构域,其三级结构没有明显差异(图3)。利用DNAMAN进行氨基酸同源比对结果(图4)表明,3条序列仅在信号肽处有两个氨基酸的编码存在差异。而冬小麦与春小麦中 TaEXPA7-A/B/D仅在核苷酸序列中存在碱基差异,蛋白序列不存在差异。众所周知,一般的蛋白质都通过保守的结构域行使其功能,而本试验中的这3条氨基酸序列仅在信号肽处存在差异,其功能是否相似还有待研究。

M为DL2000;1和2为样品
M:DL2000;1 and 2:Sample
图1东农冬麦2号根中提取的总RNA
Fig.1TotalRNAextratedfromrootofDongnongdongmai2
M为DL2000,1为样品
M:DL2000, 1:Sample
图2TaEXPA7-A/B/DCDS的PCR扩增结果
Fig.2PCRamplificationofTaEXPA7-A/B/DCDS
图3 TaEXPA7-A/B/D编码蛋白质三级结构的预测
2.3 TaEXPA7-A/B/D基因的序列比对和系统进化树的构建
在NCBI数据库中进行BLAST检索,发现TaEXPA7-A/B/D基因序列与节节麦(XP_020162651.1)、二穗短柄草(XP_003580384.1)、水稻(XP_015636818.1)等大多数单子叶植物的同源性较高,表明 TaEXPA7-A/B/D基因普遍存在于单子叶植物中。用ClustalX软件对这些氨基酸序列进行多序列比对,用MEGA 6.0进行系统进化树的构建(图5),并用BOXSHADE对这些氨基酸序列进行着色,分析其结构(图6)。结果表明,这些基因序列均具有信号肽和两个保守结构域(DPBB-1和Pollen-allerg-1),小麦与节节麦的氨基酸序列相似性最高,亲缘关系最近。
2.4 TaEXPA7-A/B/D基因的表达特性分析
以获得的RNA为模板进行反转录,采用qRT-PCR引物进行荧光定量PCR,测定东农冬麦2号TaEXPA7-A/B/D在两叶一心期根中不同组织部位的表达特性以及三个基因之间的表达差异。结果(图7)显示,TaEXPA7-A/B/D基因在根的根尖、伸长区和成熟区均有表达,且在根尖中的相对表达量都显著高于伸长区和成熟区。其中,TaEXPA7-B在根尖中的相对表达量最高。
以获得的RNA为模板进行反转录,采用qRT-PCR引物进行荧光定量PCR,测定冬小麦两叶一心期根中TaEXPA7-A/B/D基因在不同环境胁迫下的表达特性以及表达差异。结果(图8至图10)显示,无论在低温、干旱还是激素处理下,该基因均下调表达。
低温4 ℃处理后,TaEXPA7-A/B/D基因的表达量均呈现下降的趋势,处理12 h时的表达量最低,在12 h之前,TaEXPA7-D的相对表达量最高,12 h以后TaEXPA7-B的相对表达量最高。
干旱处理后,TaEXPA7-A/B/D基因的表达量均呈现下降的趋势,相对表达量表现为 TaEXPA7-B>TaEXPA7-A>TaEXPA7-D。30%PEG处理后,TaEXPA7-A/B/D基因的表达量呈现急剧下降的趋势,而20%PEG处理后基因的表达量下降得较缓慢。20%PEG处理后,在12 h时,其相对表达量最低,此后又呈现上升的趋势。

图4 TaEXPA7-A/B/D的氨基酸同源比对
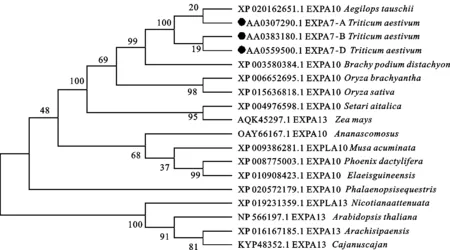
图5 TaEXPA7-A/B/D与其他不同物种同源基因编码蛋白的系统发生分析

图6 氨基酸序列相似性分析
激素处理后,TaEXPA7-A/B/D基因的表达量均呈下降的趋势。在MeJA处理后,在0~2 h之间,基因的表达量显著下降,在2~48 h之间其表达量变化较小;在ABA处理后,基因的表达量呈现先下降后上升再下降的趋势,TaEXPA7-D基因的表达量最低;在SA处理后,在0~2 h之间,基因的表达量显著下降,在2 h时其表达量最低,此后,各基因的表达量随时间的增加而缓慢上升。
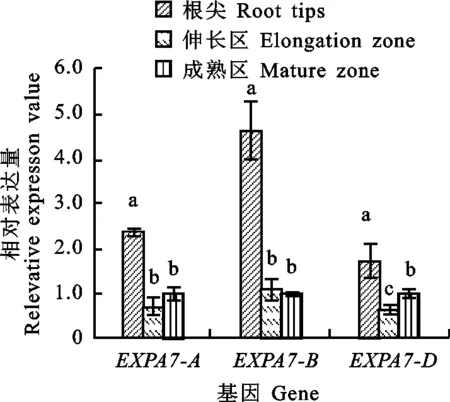
同一组图柱上不同字母表示根的不同部位之间差异显著(P<0.05)。
The different letter above columns indicate significant difference between different parts of the root(P<0.05).
图7TaEXPA7-A/B/D基因在根中不同组织部位的表达差异
Fig.7DifferentialexpressionofTaEXPA7-A/B/Dindifferentregionsofroots
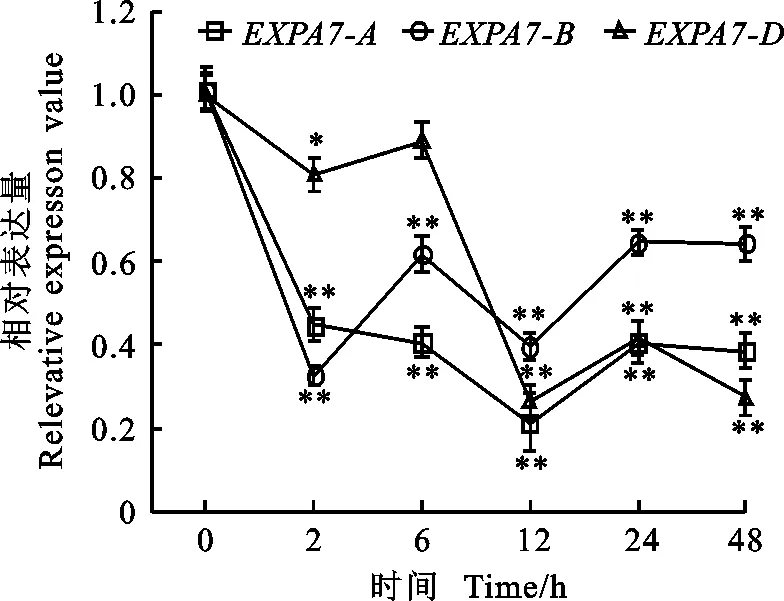
*和**分别表示与处理前差异显著(P<0.05)和极显著(P<0.01)。下同。
* and ** indicate difference between before and after treatment at 0.05 and 0.01 level , respectively. The same at following figures.
图84℃胁迫下TaEXPA7-A/B/D基因的表达差异
Fig.8DifferentialexpressionofTaEXPA7-A/B/Dunder4℃stress
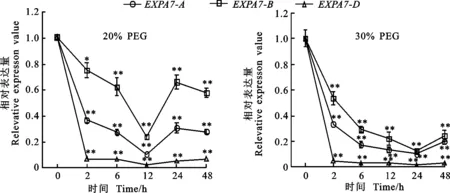
图9 干旱胁迫下TaEXPA7-A/B/D基因的表达差异
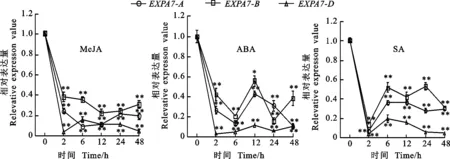
图10 不同激素胁迫下TaEXPA7-A/B/D基因的表达差异
3 讨 论
多倍化在生物学中是一种普遍存在的现象,是物种形成和进化的重要途径之一。多倍体基因组中的重复基因可能有以下三种不同的命运:保持原有的功能、基因沉默和分化并执行新的功能[10-11]。近年来,对于多倍化的研究越来越多。2007年,Shitsukawa等[12]分析了普通小麦中一个MADS 家族基因WLHS1 的3 个同源基因的结构和功能,发现WLHS1-A 基因存在一个新序列的插入,WLHS1-B的DNA高度甲基化,只有WLHS1-D 是有功能的。2010年,陈 琰等[13]采用RACE方法克隆了TaEXPB8的四个同源基因,在染色体定位时发现了多拷贝现象,其表达分析结果表明,该基因在小麦初生根的伸长区表达量最高,并且受外源高浓度生长素的抑制,由此推测出该基因可能在小麦根中起着重要作用。2012年孙其信等[14]对TaEXPA1-A/B/D基因研究发现,三个基因在幼根中的表达是沉默的,TaEXPA1-A/D在幼叶中表达,而TaEXPA1-B在幼叶中沉默,推测该基因的沉默可能与启动子中某一区域的甲基化和乙酰化有关,并对TaEXA1-A/B/D基因进行拟南芥的遗传转化,结果发现他们具有相似的功能。本试验从东农冬麦2号中克隆了TaEXPA7-A/B/D三个基因,发现其分别定位于2AL、2BL和2DL染色体上,对其核酸序列以及氨基酸序列进行分析发现,虽然核酸序列有几处的差异,但是氨基酸序列仅在信号肽位置有两处差异,三个基因的表达量表现为 TaEXPA7-B>TaEXPA7-A>TaEXPA7-D。这三个基因是否行使相似的功能以及在多倍体基因组上的作用机制还有待研究。
当受到非生物胁迫时,植物可通过调控胁迫相关基因的表达,激活植物对胁迫的响应机制,这些基因可以编码控制抗逆基因表达的调节因子,也可以直接编码与抗胁迫有关的酶和蛋白质。因此,研究基因的表达特性对分析植物抵御非生物胁迫的作用机制是十分重要的。2016年,李 飞等[15]对低温处理下东农冬麦2号小麦三叶期根组织TaEXPA基因的表达分析表明,TaEXPA5、TaEXPA6和 TaEXPA7三个基因的表达量可能与其抗御低温胁迫的能力正相关。本研究中,幼苗期根中的 TaEXAP7基因随着4 ℃处理时间的延长呈现先下降后上升的趋势,且其表达量均小于未处理时基因的表达量,由此可见,该基因在冬小麦生长的不同时期的表达量以及表达模式是不相同的,其作用机制也可能存在差异。赵美容等[16]于2016年在抗旱性不同的小麦扩展蛋白活性及基因表达分析中发现,TaEXPA3基因的表达在干旱胁迫下上调,且与品种抗旱性相关,表明干旱胁迫诱导的扩展蛋白活性的提高与扩展蛋白表达上调有关。而本研究中, TaEXPA7-A/B/D基因的表达在干旱胁迫下下调,且PEG浓度越高,其表达量随着时间的延长下降越明显。在膨胀素庞大的基因家族中,不同的基因行使着不同的功能,所以该基因也有可能参与和抗旱有关的作用机制。ABA、SA和JA是植物生长必要的激素,可通过复杂而精细的网络来调控植物不同阶段的生长发育,还可以响应各种生物与非生物胁迫,从而提高植物对环境的适应性。近年来,有人在外源激素影响基因表达特性的研究中发现,茉莉酸主要通过诱导扩展蛋白基因的上调表达来提高扩展蛋白的活性[17-19]。 TaEXPAB23在受到MeJA的处理后其mRNA水平表达量升高。本研究中, TaEXPA7-A/B/D三个基因均属于下调基因。该基因能否降低膨胀素活性,是否参与了植物生长的主要机制还有待研究。
最近,关于膨胀素基因与根系发育的研究已经有许多报道。AtEXPA7、OsEXPA10、OsEXPA7、 HvEXPB7等均是与植物根系生长相关的重要基因[20-22]。
TaEXPA7-A/B/D与 OsEXPA10基因有较高的相似性,结合本试验研究结果,推测 TaEXPA7基因也是一组和植物根系生长相关的重要基因,而根系能否正常生长发育又是冬小麦成功越冬的关键,因此推测该基因可能是影响冬小麦生长发育的重要基因。
[1]COSGROVE D J.Loosening of plant cell walls by expansins[J].Nature,2000,407(6802):321.
[2]SAMPEDRO J,COSGROVE D J.The expansinsuperfamily[J].GenomeBiology,2005,6(12):242.
[3] HU Z,HAN Z,SONG N,etal.Epigenetic modification contributes to the expression divergence of three TaEXPA1 homoeologs in hexaploid wheat(Triticumaestivum)[J].NewPhytologist,2013,197(4):1344.
[4]CHO H T,KENDE H.Expression of expansin genes is correlated with growth in deepwaterrice[J].PlantCell,1997,9(9):1661.
[5] 俞志明.EXPA7亚家族调控水稻根毛伸长机制研究[D].杭州:浙江大学,2010:9
YU Z M.Regulated mechanism of EXPA7 subclade members on rice root hair elongation [D].Hangzhou:Zhejiang University,2010:9.
[6] 文乙豪.水稻 OsEXPB2基因的功能研究[D].重庆:重庆大学,2013:77.
WENWEN Y H.Study on the function of rice OsEXPB2 gene [D].Chongqing:Chongqing University,2013:77.
[7] 何晓薇.水稻根毛发育相关基因 OsEXP17 的克隆和功能研究[D].杭州:浙江大学,2008:58
HE X W.Expression and function of OsEXP17 gene in rice root hair development [D].Hangzhou:Zhejiang University,2008:58.
[8]CHE J,YAMAJI N,SHEN R F,etal.An Al-inducible expansin gene, OsEXPA10 is involved in root cell elongation of rice.[J].PlantJournal,2016,88(1):132.
[9]HAN Y,LI A,LI F,etal.Characterization of a wheat(TriticumaestivumL.) expansin gene, TaEXPB23,involved in the abiotic stress response and phytohormone regulation[C]//7thCongress of Shandong Plant Physiology Society and Symposium on Plant Biology and Modern Agriculture,2012:49-58.
[10] 杨 继.植物多倍体基因组的形成与进化[J].植物分类学报,2001,39(4):357.
YANG J.Preparation and evolution of plant polyploid genomes [J].ChineseJournalofPlantTaxonomy,2001,39(4):357.
[11] 张连全.小麦异源六倍化过程及其在遗传育种中的应用[D].雅安:四川农业大学,2007:1.
ZHANG LIAN-QUAN.Application of heterotopic six-fold process of wheat and its application in genetic breeding [D].Ya’an:Sichuan Agricultural University,2007:1.
[12]SHITSUKAWA N,TAHIRA C,KASSAI K,etal.Genetic and epigenetic alteration among three homoeologous genes of a class E MADS box gene in hexaploid wheat.[J].PlantCell,2007,19(6):1723.
[13] 陈 琰,钟伶桃,姚颖垠,等.小麦膨胀素基因 TaEXPB8部分同源cDNA序列的克隆、定位及表达分析[J].农业生物技术学报,2010,18(4):645.
CHEN Y,ZHONG L T,YAO Y Y,etal.Cloning,mapping and expression analysis of partial homologous cDNA sequences of TaEXPB8 in wheat swelling protein [J].JournalofAgriculturalBiotechnology,2010,18(4):645.
[14]HU Z,SONG N,XING J,etal.Overexpression of three TaEXPA1 homoeologous genes with distinct expression divergence in hexaploid wheat exhibit functional retention inArabidopsis[J].PlosOne,2013,8(5):e63667.
[15] 李 飞,王晓磊,徐永清,等.低温处理下东农冬麦1号小麦根组织EXPA基因的表达分析[J].麦类作物学报,2016,36(9):1159.
LI F,WANG X L,XU Y Q,etal.Expression ofEXPAgene in wheat root tissue of Dongnongdongmai No.1 wheat under low temperature treatment [J].ActaMetallurgicaSinica,2016,36(9):1159.
[16] 赵美荣,李永春,刘 辉,等.抗旱性不同的小麦扩展蛋白活性及基因表达分析[J].江苏农业科学,2016,44(1):108.
ZHAO M R,LI Y CH,LIU H,etal.Expression of wheat protein and gene expression in wheat with different drought resistance [J] .JiangsuAgriculturalSciences,2016,44(1):108.
[17] 韩翠英.干旱胁迫下四个功能基因的表达模式与不同品种小麦的抗旱相关性分析[D].杨凌:西北农林科技大学,2015:31.
HAN C Y.Study on the relationship between the expression patterns of four functional genes and the drought resistance of different varieties of wheat under drought stress [D].Yangling:Northwest A&F University,2015:31.
[18] 段龙飞.茉莉酸信号途径上关键基因家族COI/JAZ/MYC分子进化分析[D].杨凌:西北农林科技大学,2013:7.
DUAN L F.Study on evolution of COI/JAZ/MYC molecules in key gene family of jasmonic acid signal pathway [D].Yangling:Northwest A&F University,2013:7.
[19] 王文艳.葡萄水杨酸和茉莉酸信号转导途径中4个重要基因的克隆及表达分析[D].南京农业大学,2011:6.
WANG W Y.Cloning and expression analysis of four important genes in salicylic acid and jasmonic acid signal transduction pathway [D].Nanjing:Nanjing Agricultural University,2011:6.
[20]HE X,ZENG J,CAO F,etal. HvEXPB7,a novel β-expansin gene revealed by the root hair transcriptome of Tibetan wild barley,improves root hair growth under drought stress [J].JournalofExperimentalBotany,2015,66(22):7405.
[21]XU Q,SANALKUMAR K,EMILY M,etal.Gibberellin regulation and genetic variations in leaf elongation for tall fescue in association with differential gene expression controlling cell expansion[J].ScientificReports,2016,6:30258.
[22]LIN C,CHOI H S,CHO H T.Root hair-specific EXPANSIN A7 is required for root hair elongation inArabidopsis[J].MoleculesandCells,2011,31(4):393.
CloningandCharacterizationofTaEXPA7PartialHomologousGenesinRootsofWinterWheatDongnongdongmai2
DONGJiamin1,XUYongqing1,PengLina1,FENGXu1,YAOShukuan1,ZHAOQiaoqin1,LIFenglan1,HUBaozhong1,2
(1.College of Life Science,Northeast Agriculture University,Harbin,Heilongjiang 150030,China;2.Harbin University,Harbin,Heilongjiang 150086,China)
Expansin is a protein that induces cell wall relaxation and elongation during plant growth and development, which containsEXPA,EXPB,EXLAandEXLBfour gene families,playing an important role in the formation and rapid development of plant roots. In this experiment, the root of cold winter wheat Dongnongdongmai 2 at two leaf stage was used as the material to clone the three CDS full-length of the TaEXPA7 gene.Their amino acid sequences were highly homologous with only two different amino acids at signal peptide.The nucleic acid sequence length was 777 bp, encoding 258 amino acids.These three genes were named as TaEXPA7-A, TaEXPA7-B, TaEXPA7-D, located at 2AL, 2BL and 2DL chromosomes, respectively. The protein contained two conserved domains, DPBB-1 and Pollen-allerg-1, with molecular weight of 27 670.74 Da and isoelectric point of 8.09 and they were hydrophobic proteins. The similarity between them andAegilopstauschiiandBrachypodiumdistachyonwas 99% and 92%, respectively. qRT-PCR analysis of roots of winter wheat at two leaves stage showed that TaEXPA7-A/B/D expressed higher in root tips, followed by elongation zone and mature zone. The winter wheat roots were treated with low temperature, drought and hormone and the three genes were all down regulated,the relative expression of TaEXPA7-B gene was higher and that of TaEXPA7-D was lower,which indicated that genes in different chromosomes of polyploid wheat responded differently to environmental stresses,their gene expression and response patterns were different, and the dominant chromosomes were also different under different environmental stresses. Moreover, the expression of these genes in roots may be related to stress such as low temperature, drought and exogenous hormones.They may be important gene for root growth.
Winter wheat; TaEXPA7; Partial homology; Expression characteristics
时间:2017-11-14
网络出版地址:http://kns.cnki.net/kcms/detail/61.1359.S.20171114.1027.018.html
2017-05-25
2017-07-08
国家基础科学人才培养基金项目(J1210069);中俄国际合作项目(2013DFR30270)
E-mail:1970113717@qq.com
李凤兰(E-mail:lifenglan@neau.edu.cn);胡宝忠(E-mail:bzhu@neau.edu.cn)
S512.1;S330
A
1009-1041(2017)11-1419-09
