中国沙棘克隆生长对造林密度的早期响应及其生物量分配调节机制*
2017-12-05刘丹一陈贝贝高海银刘春红张增悦李根前
姜 准 刘丹一 陈贝贝 高海银 刘春红 张增悦 邹 旭 李根前
(1.西南林业大学林学院 昆明 650224; 2.中国治沙暨沙业学会 北京 100714; 3.陕西省定边县林业工作站 榆林 718600)
中国沙棘克隆生长对造林密度的早期响应及其生物量分配调节机制*
姜 准1刘丹一2陈贝贝1高海银3刘春红3张增悦1邹 旭1李根前1
(1.西南林业大学林学院 昆明 650224; 2.中国治沙暨沙业学会 北京 100714; 3.陕西省定边县林业工作站 榆林 718600)
【目的】 探讨中国沙棘克隆生长对造林密度的早期响应及其生物量分配调节机制,确定利于种群稳定性长期维持的适宜造林密度。【方法】 设置4个造林密度,即2 500、4 444、10 000、40 000株·hm-2。调查测定时,分株生长能力采用每木检尺法,克隆繁殖能力采用子株个体计数法,克隆扩散能力采用跟踪挖掘法,地上生物量采用平均标准木法,垂直根生物量测定采用格子样方法和全挖法,水平根生物量测定采用跟踪挖掘法。【结果】 随着造林密度的增大,分株地径和冠幅生长量呈对数函数下降,而树高生长量差异不显著; 克隆繁殖、克隆扩散能力呈上凸抛物线变化即先升后降,且两者之间具有协同作用; 地上生物量分配下降、地下生物量分配上升,水平根生物量分配也呈上凸抛物线变化即先升后降; 分株生长能力与地上生物量分配呈正相关,克隆繁殖、克隆扩散能力与水平根生物量分配呈正相关。【结论】 种群通过地上与地下、水平根与垂直根生物量分配调节,在分株生长与克隆繁殖、克隆扩散之间做出权衡,从而形成与造林密度相适应的个体形态和克隆生长格局。随着造林密度的增大,分株形态呈粗-中-细的变化过程,克隆繁殖和克隆扩散能力呈低-高-低的变化过程,即克隆生长格局呈离散型-聚集型-离散型、种群稳定性呈低-高-低的变化过程。最有利于克隆生长和种群稳定性维持的适宜造林密度为15 000~20 000株·hm-2。相应的株行距为0.5 m×1.0 m~0.7 m×1.0 m。
克隆繁殖; 克隆扩散; 生物量分配; 造林密度; 中国沙棘; 毛乌素沙地
中国沙棘(Hippophaerhamnoidesssp.sinensis)是我国北方干旱、半干旱地区的优良多用途树种,果实、叶片富含多种活性物质和营养成分,可广泛用于医药、食品、饮料、化妆品及保健品制造(黄铨, 2001; 张建国等, 2005; 廉永善等, 1989; 段爱国等, 2013)。中国沙棘又是典型的克隆植物(clonal plant),水平根(侧根)的延伸和萌蘖能力极强,如能正确运用其克隆习性,人工造林便可获得“栽一株,成一片”的效果(Silvertown, 1993; Takahashietal., 2011; 高志义, 1989; 李根前等, 2001a; 2001b; 2004)。近年来沙棘人工林出现了大面积早衰现象(惠兴学等, 2009; 李秀寨等, 2005,这可能是造林(定居)后依靠克隆生长(分株生长、克隆繁殖和克隆扩散)实施种群扩散并维持其稳定性,克隆生长能力强则种群稳定性高、克隆生长能力弱则种群稳定性低(贺斌等, 2007a; 2007b; 2012; 曾诚等, 2016)。克隆生长能力的强弱则取决于环境资源水平的高低,只有环境资源水平适宜时才能使克隆生长能力得到充分发挥,环境资源水平过高或过低均可抑制克隆生长,据此规律即可确定利于克隆生长的适宜环境资源水平(李甜江等, 2010a; 2010b; Goughetal., 2012; Caoetal., 2016),而作为最易调控的造林密度研究还未见报道。李孙玲等(2011)、刘丹一等(2012)研究了沙棘造林密度对母株存活、分株生长的影响,但忽视了克隆繁殖、克隆扩散对造林密度的响应规律,因此无法确定克隆生长对造林密度的响应规律以及利于种群稳定性维持的适宜造林密度。目前确定造林密度的主要依据仍是立地条件、林分种类等(吴宗凯等, 2009; 李根前等, 2000; 韩江万, 2004)。为探讨中国沙棘克隆持久性的适宜造林密度和种群稳定性的维持,本文采用毛乌素沙地的试验数据,在分析克隆生长对造林密度响应规律的基础上,探讨密度效应规律的生物量分配调节机制,以期为中国沙棘造林密度设计及林分密度管理提供理论依据,并为克隆植物的造林密度估计提供新思路。
1 研究区概况
研究区位于毛乌素沙地南缘的陕西省定边县,(107°15′—108°22′E、36°49′—37°53′N),属温带干旱、半干旱大陆性季风气候,其特点是四季分明、阳光充足、干旱缺水、春季和秋季沙尘暴肆虐; 年均气温7.9 ℃,最高37.7 ℃、最低-29.4 ℃; 年均降水量316 mm,年蒸发量2 490 mm,年均相对湿度53.1%。地貌特征是沙丘起伏、沙带绵延,海拔1 303~1 418 m,以风沙土和盐碱土为主。地带性植被为干草原和荒漠草原,区系成分以旱生、耐盐植物和草甸植物为主。试验地设在林业站试验基地内,使用经人工平整的风沙土。
2 研究方法
2.1试验设计
试验采用单因素设计,分4个造林密度水平,即2 500、4 444、10 000、40 000株·hm-2,株行距分别为2 m×2 m、1.5 m×1.5 m、1 m×1 m、0.5 m×0.5 m,田间排列采用随机区组法,每个小区面积40 m2,重复3次。其中,所用苗木为规格基本一致的实生苗,栽植后浇水使其成活,此后的管理措施相同。
2.2试验调查与测定
造林后,中国沙棘分株生长、克隆繁殖、克隆扩散能力决定种群的稳定性,因此对这些指标进行了逐一测定。其中,分株生长能力采用个体大小(树高、地径、冠幅生长量)表示,克隆繁殖能力采用克隆(萌蘖)子株数量表示,克隆扩散能力采用水平根的延伸距离和分枝强度表示。1)分株(包括母株和子株)生长能力采用每木检尺法,逐株测定树高、地径、冠幅生长量。2)克隆繁殖能力采用个体计数法,分别试验小区统计克隆(萌蘖)子株的个体数量。3)克隆扩散能力采用跟踪挖掘法,从某一个一级水平根(源自母株)开始,通过跟踪挖掘逐步将与之联结的二级水平根(源于一级水平根)、三级水平根(源于二级水平根)等等全部挖出,测定各级水平根粗度、长度、条数以及分枝级数,直到将小区平均标准木(母株)的地下联结构件全部挖出并完成相应的测定。4)地上生物量采用平均标准木法,根据每木检尺结果分别选择母株、子株平均标准木,并分别母株、子株以及树干、枝条、叶片称其重量; 垂直根生物量测定采用格子样方法和全挖法,先以母株为中心、株行距为依据确定格子样方面积,如株行距为2 m×2m则以母株为中心向四周各挖1 m,然后将样方内的垂直根全部挖出称质量; 水平根生物量测定采用跟踪挖掘法,将其全部挖出称质量。然后,取一定数量样品带回实验室烘至恒质量称其干质量并计算含水率。最后,根据生物量鲜质量、含水率、母株数量、子株数量等推算种群以及不同构件单位面积的生物量。
在上述观测中,生长量、子株数量于5—8月份连续观测3年; 生物量、克隆扩散能力等破坏性取样于造林第3年秋末进行,此时对应造林密度Ⅰ、Ⅱ、Ⅲ、Ⅳ的母株存活率分别为90.0%、100.0%、98.0%、97.5%,林分郁闭度分别为0.39、0.84、0.88、0.88。
3 结果与分析
3.1克隆生长对造林密度的响应
3.1.1 分株生长对造林密度的响应 由表1可知: 树高生长量在造林密度Ⅰ、Ⅱ、Ⅲ、Ⅳ之间差异不显著; 地径生长量在造林密度Ⅰ、Ⅱ、Ⅲ之间差异不显著,但造林密度Ⅳ的分株地径生长量显著小于造林密度Ⅰ、Ⅱ、Ⅲ; 冠幅生长量在造林密度Ⅰ、Ⅱ之间差异不显著,但造林密度Ⅲ的分株冠幅生长量显著小于造林密度Ⅰ和Ⅱ、造林密度Ⅳ的分株冠幅生长量又显著小于造林密度Ⅲ。由图1可知: 地径、冠幅生长量随着造林密度的增大呈对数函数下降(Plt;0.001)。由此表明: 造林密度对树高生长量影响不显著,但地径和冠幅生长量随着造林密度的增大而下降。

表1 不同造林密度之间分株生长量差异显著性检验①Tab.1 Difference significance analysis of the parameters characterizing the ramet growth with different planting densities
①字母不同表示在Plt;0.05的水平差异显著,字母相同表示差异不显著。下同。Different letters next to the number indicate the significant differences between the families atPlt;0.05,the same letters do the insignificant differences.The same below.

图1 地径与造林密度的关系Fig.1 Dependence of basal diameter on initial planting density
3.1.2 克隆繁殖与克隆扩散对造林密度的响应 由表2、图2可知: 随着造林密度的增大,克隆繁殖(子株数量)以及克隆扩散(水平根粗度、长度、分枝级数、分枝条数)能力呈上凸抛物线变化,方程顶点即为特定参数最大时的相应最佳造林密度。小于最佳造林密度,克隆繁殖、克隆扩散能力随着造林密度的增大而上升; 等于最佳造林密度,克隆繁殖、克隆扩散能力最大; 大于最佳造林密度,克隆繁殖、克隆扩散能力随着造林密度的增大而下降。由此表明: 克隆繁殖、克隆扩散能力随着造林密度的增大先升后降,最有利于克隆繁殖和克隆扩散能力发挥的适宜造林密度约为15 000~20 000株·hm-2。
由表3可知: 子株数量与一级水平根粗度、长度、条数以及水平根分枝级数之间呈正相关。由此表明: 克隆繁殖、克隆扩散能力对造林密度的响应具有协同性,即克隆繁殖能力随着克隆扩散(水平根的延伸和分枝)能力的增强而升高、随着克隆扩散能力的减弱而下降。
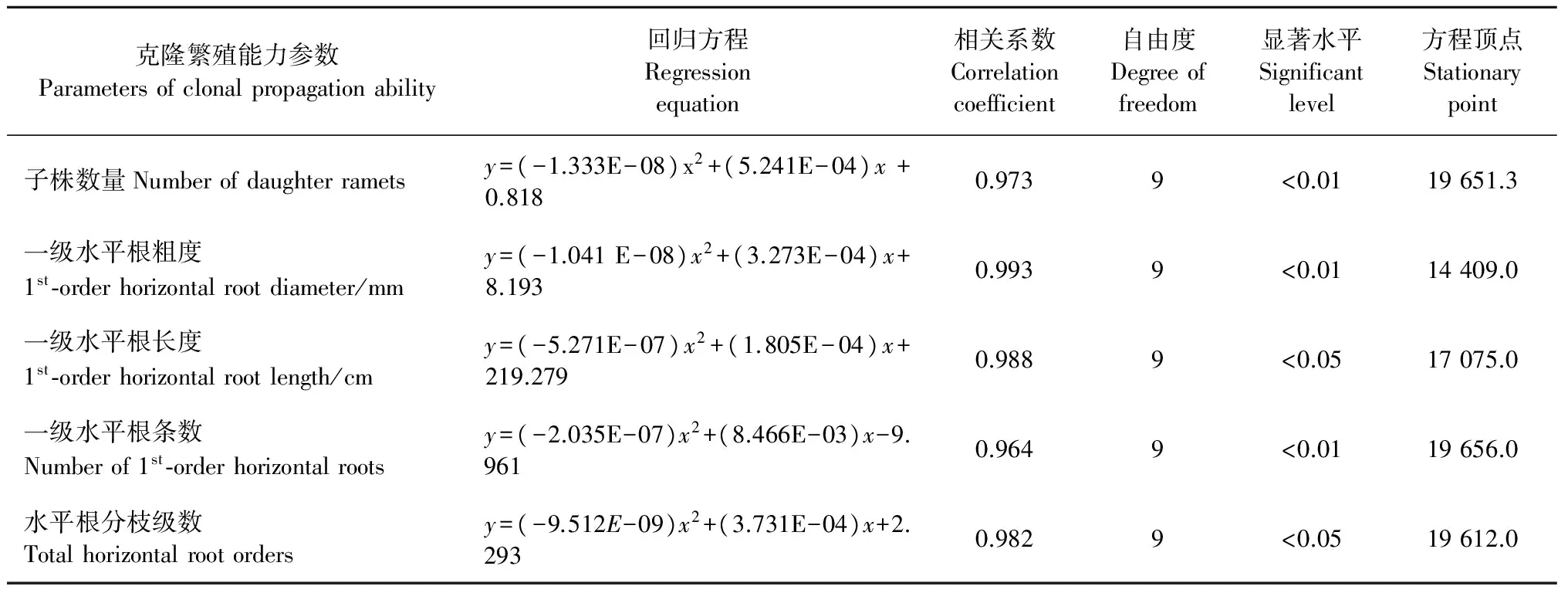
表2 克隆繁殖与克隆扩散能力对造林密度的响应Tab.2 Response of clonal propagation and clonal expansion to initial planting density
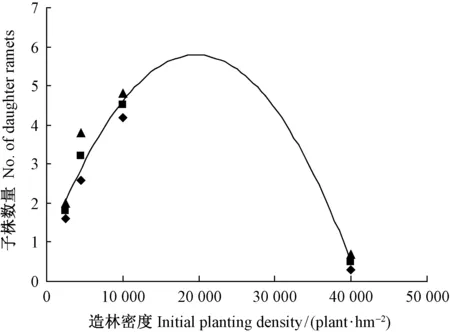
图2 克隆繁殖能力与造林密度的关系Fig.2 Dependence of clonal propagation on initial planting density
3.2生物量及分配对造林密度的响应
3.2.1 生物量对造林密度的响应 由表4、表5、图3可知: 种群、地上、地下生物量随造林密度的增大而上升,4个造林密度之间存在显著差异。地上构件中,树干、活枝生物量先上升然后趋于上限,在造林密度Ⅰ、Ⅱ与Ⅲ及Ⅳ之间存在显著差异; 叶片生物量随着造林密度的增大而上升,在4个造林密度之间存在显著差异; 枯枝生物量随着造林密度的增大而上升,在造林密度Ⅰ、Ⅱ与Ⅲ、Ⅳ之间存在显著差异。地下构件中,垂直根生物量随着造林密度的增大而上升,4个造林密度之间存在显著差异; 水平根生物量随着造林密度的增大先升后降,4个造林密度之间存在显著差异。由此表明: 种群及其存活构件(干、枝、叶、垂直根)随着造林密度的增大呈对数变化趋势,即先上升然后趋于上限; 死亡构件(枯枝)随着造林密度的增大呈直线上升; 水平根生物量随着造林密度的增大呈上凸抛物线变化,即先升后降。
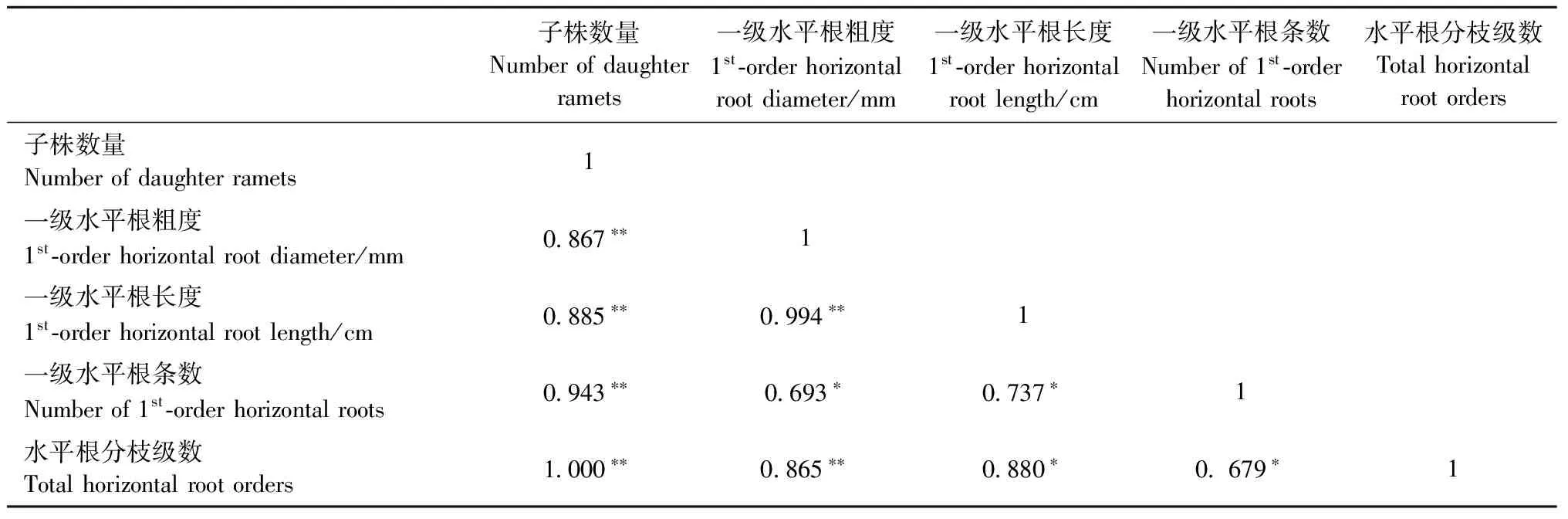
表3 克隆繁殖与克隆扩散能力的相关性①Tab.3 Correlation between clonal propagation and clonal expansion
①**Plt;0.01,*Plt;0.05.

表4 种群及其构件生物量差异显著性检验Tab.4 Difference significance test of biomass accumulation of population and its modules
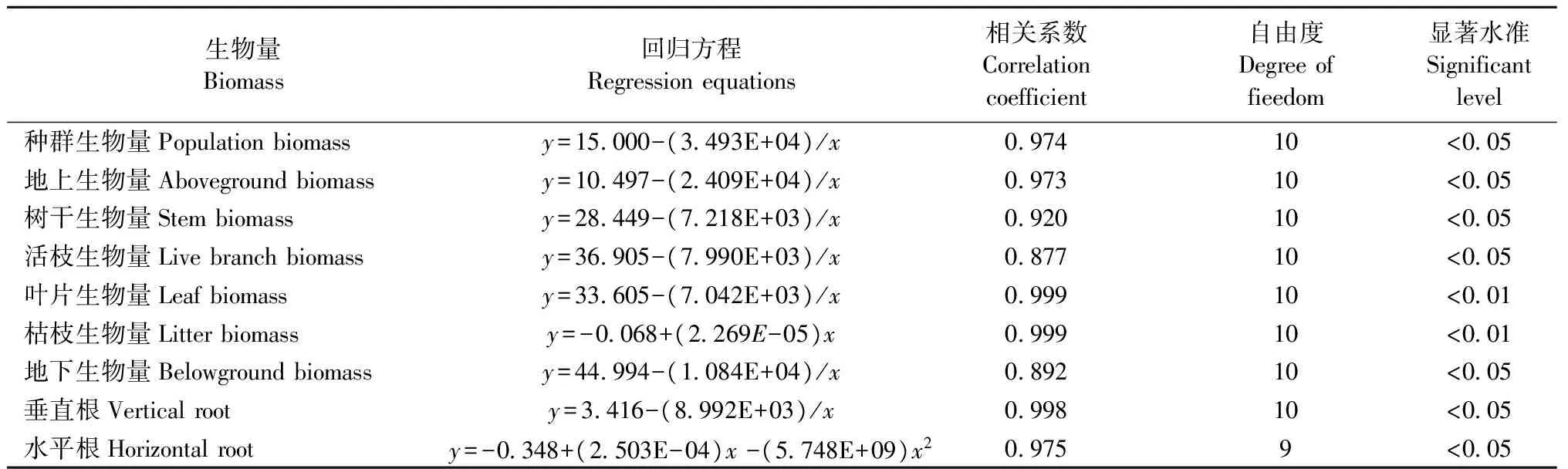
表5 种群及其构件生物量与造林密度的关系Tab.5 Relationship between population biomass accumulation as well as its modules and initial planting density

图3 生物量与造林密度的关系Fig.3 Relationship between biomass accumulation and initial planting density
3.2.2 生物量分配对造林密度的响应 由表5可知: 地上生物量分配随造林密度的增大而下降,地下生物量分配随造林密度的增大而上升。在地上生物量的再分配中,树干生物量分配随造林密度的增大而上升,活枝、叶片的生物量分配随造林密度的增大而下降; 在地下生物量的再分配中,垂直根生物量分配随造林密度的增大而上升,水平根的生物量分配随造林密度的增大先升后降。由表7可知: 地上与地下生物量分配呈负相关(r=-1.000),树干与枝条、叶片生物量分配呈负相关,垂直根与水平根的生物量分配呈负相关。由此表明: 地上与地下、树干与枝叶、垂直根与水平根之间的生物量分配存在权衡关系,即此长彼消。
3.3克隆生长对生物量及分配的响应
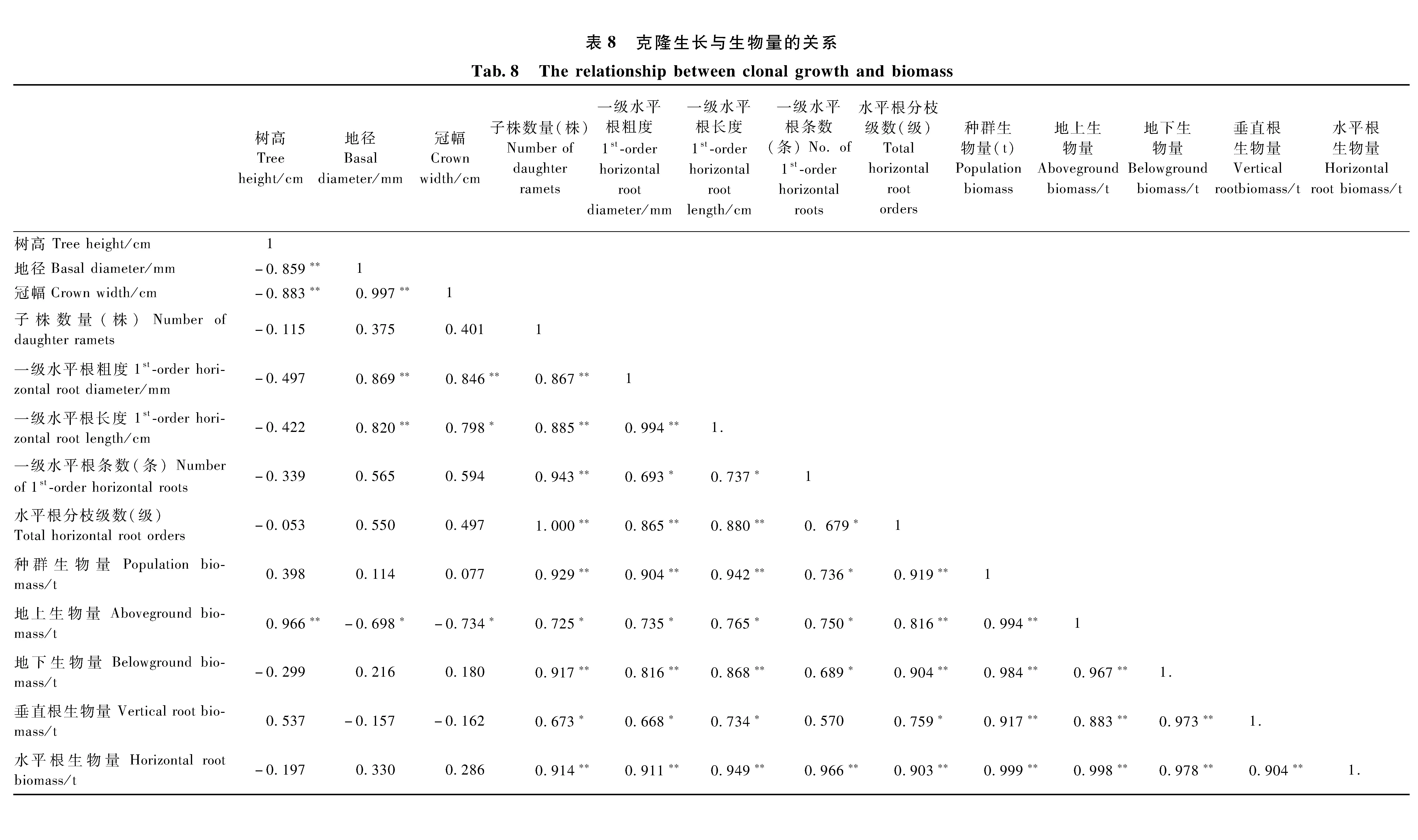
3.3.1 克隆生长对生物量投资的响应 由表7可知: 树高生长量与地上生物量投资呈正相关,而地径、冠幅生长量与地上生物量投资呈负相关; 子株数量以及水平根粗度、长度、分枝级数、分枝条数与种群、地上、地下、垂直根、水平根生物量投资呈正相关。由此表明: 随着生物量投资的增大,克隆繁殖、克隆扩散能力随之提高,而分株形态逐步由粗壮向纤细转变。
种群、地上、地下、垂直根和水平根的生物量投资之间呈正相关,说明它们之间具有协同作用,即此涨彼涨、此消彼消。
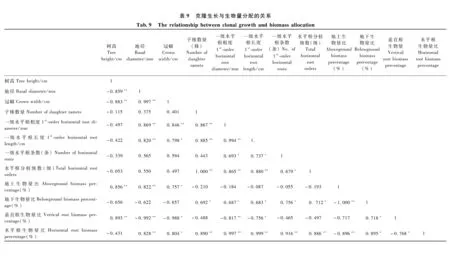
3.3.2 克隆生长对生物量分配的响应 由表8可知: 分株树高、地径、冠幅生长量与地上生物量分配呈正相关,与地下生物量分配具有负相关趋势; 子株数量以及水平根粗度、长度、分枝级数、分枝条数与地下生物量分配呈正相关,与地上生物量分配具有负相关趋势。 子株数量以及水平根粗度、长度、分枝级数、分枝条数与水平根生物量分配呈极显著正相关,而与垂直根生物量分配呈负相关或具有负相关趋势。由此表明: 随着地上生物量分配的增大,分株生长量随之提高; 随着水平根生物量分配的增大,克隆繁殖、克隆扩散能力随之提高。
由表9还可知: 分株地径、冠幅生长量与水平根粗度、长度呈正相关,说明水平根延伸越远则分株地径、冠幅越大; 分株树高与地径、冠幅呈负相关,说明随着造林密度的增大分株形态逐渐由粗壮向纤细转变。
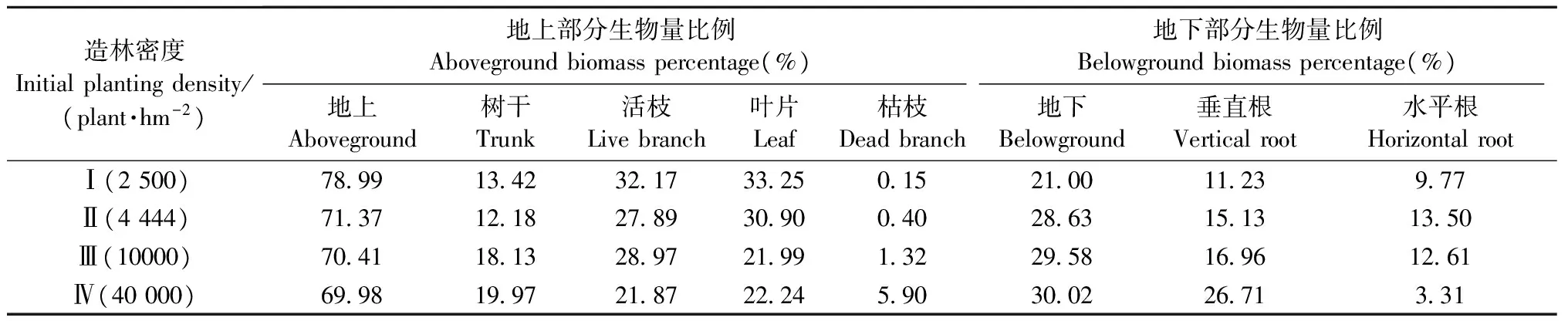
表6 生物量分配对造林密度的响应Tab.6 Response of biomass allocation to initial planting density

表7 生物量各分配组分相关分析Tab.7 The biomass allocation of each component related analysis
4 讨论
随着造林密度的增大,地上生物量分配减小而水平根生物量分配先升后降; 分株(地径和冠幅)生长能力与地上生物量分配呈正相关,克隆繁殖、克隆扩散能力与水平根生物量分配呈正相关。同时,地上与地下、水平根与垂直根生物量分配存在权衡关系,克隆繁殖与克隆扩散能力存在协同作用。在不同的造林密度下,种群通过地上与地下、水平根与垂直根生物量分配调节,形成与造林密度相适应的分株形态和克隆生长格局。造林密度过小,地上生物量分配高而水平根生物量分配低,种群以分株粗壮、冠幅较大、子株稀少为特征,克隆繁殖和克隆扩散能力弱; 造林密度适宜,地上生物量分配呈中等水平而水平根生物量分配高,种群以分株中等、树冠适中、子株密集为特征,克隆繁殖和克隆扩散能力强; 造林密度过大,地上生物量和水平根生物量分配均低而垂直根生物量分配高,种群以分株纤细、树冠较小、子株稀少为特征,克隆繁殖和克隆扩散能力弱。由此可见: 随着造林密度的增大,分株形态呈粗壮-中等-纤细的变化过程,克隆繁殖和克隆扩散能力呈低-高-低的变化过程。因此,造林密度过小或过大都会增大种群的早衰概率。
在不同的环境资源水平下,中国沙棘通过生物量分配调节使种群形成与之相适应的克隆生长格局,以便充分利用环境资源、削弱克隆内分株间的竞争,从而以更高的概率维持克隆持久性与种群稳定性(李根前等, 2001c; 肖智勇等, 2011; 刘明等, 2014; 贺斌等, 2006; Dongetal., 2012)。同样,在不同的造林密度下中国沙棘种群同样通过地上与地下、水平根与垂直根生物量分配调节,在分株生长与克隆繁殖、克隆扩散之间做出权衡,从而形成与造林密度相适应的个体形态与克隆生长格局。造林密度过小时,空间和环境资源相对充裕,种群首先对地上空间、资源展开竞争,因此将更多的生物量分配于地上构件从而促进分株的生长,但也因此降低了水平根的生物量分配从而削弱克隆繁殖和克隆扩散能力,导致分株相对粗壮但子株稀少,克隆生长格局倾向于“离散型”; 造林密度适宜时,种群对地上、地下空间和环境资源同时展开竞争,地上、地下生物量分配比较均衡,但在地下生物量的再分配中种群将更多的生物量分配于水平根,克隆繁殖和克隆扩散能力得到充分地展示,子株密集但分株大小呈现中等水平,克隆生长格局倾向于“聚集型”; 造林密度过大时,地上空间和环境资源短缺,种群首先对地下空间和环境资源展开竞争和觅食,因此将更多的生物量分配于地下构件尤其是垂直根,这样利用种群对地下空间、环境资源的竞争和觅食,导致分株生长、克隆繁殖和克隆扩散受到抑制,子株稀少且分株比较纤细,克隆生长格局倾向于“离散型”。研究表明: “聚集型”格局利于种群对生境资源的占据和利用,并可增强种群对其他物种入侵的排斥能力; “离散型”格局可使克隆在更大的空间内占据和获取必要的生境资源,尽可能地避免克隆内部分株之间的竞争,并提高克隆将子株放置在有利生境斑块的概率,但以降低分株生长、克隆繁殖能力为代价(董鸣, 1996; Ningetal., 2014; Wangetal., 2011; Hutchings, 1994; Salzman, 1985; Yanetal., 2013; Ronaldetal., 1996; Dongetal., 2013; Qiangetal., 2014)。“聚集型”格局有利于克隆持久性和种群稳定性的维持。因此,随着造林密度的增大,克隆生长格局呈现离散型-聚集型-离散型的变化,种群稳定性呈现低-高-低的变化。
调节过程中,克隆可塑性与其竞争的主要资源一致 (Slade, 1987a; 1987b; Alpert, 1991; Evans, 1992; Luoetal., 2013)。由于中国沙棘集中分布在干旱、半干旱地区,对土壤水分的竞争和觅食起着决定性作用。 随着土壤水资源水平的提高,克隆繁殖和克隆扩散能力呈上凸抛物线变化即先升后降,因此土壤水资源水平过高或过低均会抑制克隆生长从而增大种群早衰概率(李甜江等, 2010a; Luo, 2002; Caoetal., 2016; 贺斌等, 2007b)。根据回归方程估计,利于克隆繁殖和克隆扩散的土壤含水率小于16.0%而利于分株生长的土壤含水率大于16.0%(唐翠平, 2015)。 造林密度Ⅰ、Ⅱ、Ⅲ、Ⅳ的土壤含水率分别为16.20%、14.11%、9.60%、6.62%,4个处理之间存在极显著差异。因此,过小的造林密度有利于土壤水分维持,促进了种群对地上空间和资源的竞争,地上生物量分配增大而地下(尤其是水平根)生物量分配减小,分株生长潜力大而克隆繁殖和克隆扩散能力受到削弱; 过大的造林密度导致土壤水分过度消耗,促进了种群对土壤水资源的竞争和觅食,因此垂直根生物量分配增大而地上及水平根生物量分配减小,分株生长、克隆繁殖、克隆扩散能力均受到抑制。相关研究也表明,克隆可塑性具有明显的密度制约作用,包括分株大小、子株多少以及水平根的延伸、分枝能力和垂直根发育,且地上构件与地下构件的可塑性存在密切的内在联系(Charpentieretal., 2012; 李孙玲等, 2011; 刘丹一等, 2012)。 中国沙棘能够通过克隆生长调节对造林密度做出整合地响应,从而适应相应的环境资源水平。
5 结论
中国沙棘种群生物量分配格局与造林密度具有极其密切的内在联系,生物量分配格局影响克隆生长能力(格局),而克隆生长能力(格局)决定种群稳定性。在不同的造林密度下,种群通过生物量分配调节对分株生长与克隆繁殖、克隆扩散做出权衡,从而形成与造林密度相适应的分株形态与克隆生长格局,这不仅是一种生态适应对策,也是维持克隆持久性和种群稳定性的生物学途径。根据克隆生长能力与造林密度的回归方程估计,最有利于种群稳定性(克隆生长能力)维持的适宜造林密度为15 000~20 000株·hm-2,相应的株行距为0.5 m×1.0 m~0.7 m×1.0 m,这一研究结果可作为毛乌素沙地流动沙区沙棘防护林营造的参考,在其他立地类型或林种应用时可做适当调整。此外,本试验阐述了中国沙棘克隆生长的早期密度效应,随着分株个体增大和分株数量增多,回归方程顶点必然左移,即适宜造林密度减小因此,生产中可适度降低造林密度并与异质性造林设计相结合解决这一问题。
董 鸣. 1996. 资源异质性环境中的植物克隆生长: 觅食行为. 植物学报, 38 (10): 828-835.
(Dong M. 1996. Clonal growth in plants in relation to resource heterogeneity foraging behavior. Act a Botanica Sinica, 38 (10): 828-835. [in Chinese])
段爱国, 张建国, 何彩云, 等. 2013. 沙棘良种 ‘白丘杂’. 林业科学, 49 (11): 196.
(Duan A G, Zhang J G, He C Y,etal. 2013. A newHippophaerhamniodeshybrid‘Baiqiuza’. Forest Research, 49 (11): 196. [in Chinese])
高志义, 张玉胜. 1989. 沙棘根系特性的观察与研究. 北京林业大学学报, 11 (4): 53-59.
(Gao Z Y, Zhang Y S. 1989. The observation and investigation on the feature of root system of seabuckthorn. Journal of Beijing Forestry University, 11 (4): 53-59. [in Chinese])
韩江万. 2004. 甘南白龙江流域沙棘育苗及造林技术. 甘肃农业科技, (5): 36-37.
(Han J W. 2004. Seedling raising and afforestation techniques ofHippophaerhamniodessubsp.sinensisin Longjiang white Gannan River Basin.Agricultural Science and Technology, (5): 36-37. [in Chinese])
贺 斌, 赵粉侠, 李根前, 等. 2012. 毛乌素沙地中国沙棘克隆生长对土壤水分含量的响应. 南京林业大学学报:自然科学版, 36(4): 46-50.
(He B, Zhao F X, Li G Q,etal. 2012. The response of clonal growth ofHippophaerhamniodesL. subspsinensisto the availability of soil moisture in Mu Us sand land. Journal of Nanjing Forestry University: Natural Science Edition, 36 (4): 46-50. [in Chinese])
贺 斌, 李根前, 高海银, 等. 2007a. 不同土壤水分条件下中国沙棘克隆生长的对比研究. 云南大学学报:自然科学版, 29(1): 101-107.
(He B, Li G Q, Gao H Y,etal. 2007a. A comparison study on theclonal growth ofHippophaerhamniodesL.subsp.sinensisat different soil moisture condition. Journal of Yunnan University:Natural Sciences Edition, 29 (1): 101-107. [in Chinese])
贺 斌, 李根前. 2007b. 木本克隆植物中国沙棘种群数量与结构对土壤水分的响应. 西北农林科技大学学报:自然科学版, 35(3): 183-187.
(He B, Li G Q. 2007b. Population quantity and population structure in response to soil moisture in the clonal tree speciesHippophaerhamniodesL.subsp.sinensis. Journal of Journal of Northwest Agriculture and Forest University:Natural Sciences Edition, 35 (3): 183-187. [in Chinese])
贺 斌, 李根前, 徐德兵, 等. 2006. 沙棘克隆生长及其生态学意义. 西北林学院学报, 21 (3): 54-59.
(He B, Li G Q, Xu D B,etal. 2006. The clonal growth and its ecological significance ofHippophae. Journal of Northwest Forestry University, 21 (3): 54-59. [in Chinese])
黄 铨. 2001. 种植沙棘加速“三北”生态环境建设. 林业科学, 37(2): 20.
(Huang Q. 2001. Accelerate theHippophaecultivation of ecological environment construction of ‘Three North’. Scientia Silvae Sinicae, 37 (2): 20. [in Chinese])
惠兴学, 洪 新, 于 欣, 等. 2009. 辽西地区沙棘退化林分的修复与前景. 国际沙棘研究与开发, 6 (4): 24-27.
(Hui X X, Hong X, Yu X,etal. 2009. Restoration of degraded seabuckthorn stands and prospects in the western Liaoning. The Global Seabuckthorn Research and Development, 6 (4): 24-27. [in Chinese])
李根前, 黄宝龙, 唐德瑞, 等. 2001a. 毛乌素沙地中国沙棘无性系种群年龄结构动态与遗传后果研究. 应用生态学报, 12 (3): 347~350.
(Li G Q, Huang B L, Tang D R,etal. 2001a. Age structure dynamics and genetic consequences ofHippophearhamnoidesL. subsp.sinensisclone population in Mu Us Sand. Chinese Journal of Applied Ecology, 12 (3): 347-350. [in Chinese])
李根前, 黄宝龙, 唐德瑞, 等. 2001b. 毛乌素沙地中国沙棘克隆生长调节. 应用生态学报, 12 (5): 682-686.
(Li G Q, Huang B L, Tang D R,etal. 2001b. Regulation of clonal growth ofHippophaerhamnoidesL. subspsinensispopulation in Mu Us Sandland. Chinese Journal of Applied Ecology, 12 (5): 682-686. [in Chinese])
李根前, 黄宝龙, 唐德瑞, 等. 2001c. 毛乌素沙地中国沙棘无性系生长格局与生物量分配. 西北农林科技大学学报:自然科学版, 29 (2): 51-55.
(Li G Q, Huang B L, Tang D R,etal. 2001c. Studies on the clonal growth pattern and biomass allocation ofHippophaerhamnoidesL. subspsinensispopulation in Mu Us Sandland. Journal of Journal of Northwest Agriculture and Forest University:Natural Sciences Edition, 29 (2): 51-55. [in Chinese])
李根前, 赵粉侠, 李秀寨, 等. 2004. 毛乌素沙地中国沙棘种群数量动态研究. 林业科学, 40 (1): 180-185.
(Li G Q, Zhao F X, Li X Z,etal. 2004. Density and biomass dynamics ofHippophaerhamnoidesL.subsp.sinensispopulation in Mu Us Sand. Scientia Silvae Sinicae, 40 (1): 180-185. [in Chinese])
李根前, 唐德瑞, 赵一庆, 等. 2000. 沙棘林培育技术研究. 沙棘, 13 (3): 12-17.
(Li G Q, Tang D R, Zhao Y Q,etal. 2000. Studying on Cultivation Technology ofHippophaerhamniodessubsp.sinensis. Hippophae, 13 (3): 12-17. [in Chinese])
李孙玲, 李甜江, 李根前, 等. 2011. 毛乌素沙地中国沙棘存活及生长对灌水和密度的响应. 西北林学院学报, 26 (3): 107-111.
(Li S L, Li T J, Li G Q,etal. 2011. The growth and survival ofHippophaerhamniodesL. ssp.sinensisin response to irrigation intensity and initial density in Mu Us Sand. Journal of Northwest Forestry University, 26 (3): 107-111. [in Chinese])
李甜江, 李根前, 徐德兵, 等. 2010a. 中国沙棘克隆生长对灌水强度的响应. 生态学报, 30 (24): 6952-6960.
(Li T J, Li G Q, Xu D B,etal. 2010a. The clonal ofHippophaerhamniodesL.ssp.sinensisin response to irrigation intensity. Acta Ecologica Sinica, 30 (24): 6952-6960. [in Chinese])
李甜江, 李根前, 徐德兵, 等. 2010b. 克隆植物中国沙棘生长对外源植物激素的响应. 生态学报, 30 (3): 0659-0667.
(Li T J, Li G Q, Xu D B,etal. 2010b. The growth of clonal plantHippophaerhamnoidesL. ssp.sinensisin response to exogenous phytohorm one. Acta Ecologica Sinica, 30 (3): 0659-0667. [in Chinese])
李秀寨, 李根前, 韦 宇, 等. 2005. 中国沙棘大面积死亡原因的探讨. 沙棘, 18 (1): 24-28.
(Li X Z, Li G Q, Wei Y,etal. 2005. Causes of seabuckthorn death in large acreage in China. Hippophae, 18 (1): 24-28. [in Chinese])
廉永善, 陈学林, 郭建魁, 等. 1989. 甘肃省的沙棘资源及开发设想. 林业科学, 25 (1): 82-85.
(Lian Y S, Chen X L, Guo J K,etal. 1989. Resources and exploitative assumption forHippophae.L.in Gansu Province. Forest Research, 25 (1): 82-85. [in Chinese])
刘丹一, 李甜江, 李根前, 等. 2012. 初植密度对中国沙棘林木生长特征的影响. 西部林业科学, 41 (6): 78-83.
(Liu D Y, Li T J, Li G Q,etal. 2012. Effect of planting density on growing characteristics ofHippophaerhamniodessubsp. sinensis. Journal of West China Forestry Science, 41 (6): 78-83. [in Chinese])
刘 明, 唐翠平, 郭 峰, 等. 2014. 克隆植物中国沙棘种群动态及其生物量调节机制. 南京林业大学学报:自然科学版, 38 (4): 57-63.
(Liu M, Tang C P, Guo F,etal. 2014. A study on the population dynamics andits biomass adjustment mechanisms of clonal plantHippophaerhamhoidesL. ssp.sinensis. Journal of Nanjing Forestry University:Natural Science Edition, 38 (4): 57-63. [in Chinese])
唐翠平. 2015. 中国沙棘克隆生长对组织含水率的响应.昆明:西南林业大学硕士学位论文.
(Tang C P. 2015. The Clonal growth ofHippophaerhamnoidesL. ssp.sinensisin response to tissue water content. Kunming: MS thesis of Southwest Forestry University. [in Chinese])
吴宗凯, 刘广全, 匡尚富, 等. 2009. 黄土高原半干旱区退耕地沙棘林密度调控. 国际沙棘研究与开发, 7 (3): 5-10.
(Wu Z K, Liu G Q, Kuang S F,etal. 2009. Controlling seabuckthorn densities for returning into woodland in semi-arid region of The Loess Plateau. The Global Seabuckthorn Research and Development, 7(3): 5-10. [in Chinese])
肖智勇, 李根前, 代光辉, 等. 2011. 黄土高原不同坡向中国沙棘种群生物量投资与分配. 东北林业大学学报, 39 (5): 44-46.
(Xi Z Y, Li G Q, Dai G H,etal. 2011. Biomass allocation and investment ofHippophaerhamnoidesssp.sinensispopulations under different aspects of slope in the Loess Platea. Journal of Northeast Forestry University, 39 (5): 44-46. [in Chinese])
曾 诚, 陈贝贝, 李根前, 等. 2016. 毛乌素沙地土壤水分对中国沙棘人工林稳定性及生产力的影响. 林业资源管理, (1): 99-104.
(Zeng C, Chen B B, Li G Q,etal. 2016. Influence of soil physico-chemical properties in Maowusu Sandland on the stability and productivity ofHippophaerhamnoidessubsp.sinensisPlantation. Forest Resources Management, (1): 99-104. [in Chinese])
张建国, 罗红梅, 黄 铨, 等. 2005. 大果沙棘不同品种果实特性比较研究. 林业科学研究, 18 (6): 643-650.
(Zhang J G, Luo H M, Huang Q,etal. 2005. A comparative study on berry characteristics of large berry cultivars of sea buckthorn. Forest Research, 18 (6): 643-650. [in Chinese])
Alpert P. 1991. Nitrogen sharing among ramets increases clonal growth inFragariachiloensis. Ecol, 72 (1): 69- 80.
Cao Z L, Li T J, Li G Q. 2016. Modular growth and clonal propagation ofHippophaerhamnoidessubsp.sinensisin response to irrigation intensity. Journal of Forestry Research, 27 (5): 1019-1028.
Charpentier A, Anand M, Bauch C T. 2012. Variable offspring size as an adaptation to environmental heterogeneity in a clonal plant species: integrating experimental and modeling approaches. Journal of Ecology, 100 (1): 184-195.
Dong B C, Wang J Z, Liu R H,etal. 2013. Effects of heterogeneous competitor distribution and ramet aggregation on the growth and size structure of a clonal plant. Plos One, 8 (7): 1-12.
Dong B C,Alpert P, Guo W,etal. 2012. Effects of fragmentation on the survival and growth of the invasive, clonal plantAlternantheraphiloxeroides. Original Paper, 14 (6): 1101-1110.
Evans J P. 1992. The effect of local resource availability and clonal integration on ramet functional morphology inHydrocotylehonariensis. Oecologia, 89: 265- 276.
Gough L, Cross K L, Cleland E E,etal. 2012. Incorporating clonal growth form clarifies the role of plant height in response to nitrogen addition. Oecologia, 169 (4):1053-1062.
Hutchings M J, de Kroon H. 1994. Foraging in plants: the role of morphological plasticity in resource acquisition. Advances in Ecological Research, 25: 159-238.
Luo D, Qian Y Q, Han L,etal. 2013. Phenotypic responses of a stoloniferous clonal plantBuchloedactyloidesto scale-dependent nutrient heterogeneity. Plos One, 8 (6):1591-1594.
Luo X G, Dong M.2002. Architectural plasticity in response to soil moisture in the stoloniferous Herb,Duchesneaindica. Acta Botanica Sinica, 44 (1): 97-100.
Ning Y, Zhang Z X, Cui L J,etal. 2014. Adaptive significance of and factors affecting plasticity of biomass allocation and rhizome morphology: a case study of the clonal plantScirpusplaniculmis. Polish Journal of Ecology, 62 (1): 77-88.
Qiang Y Q, Luo D, Gong G,etal. 2014. Effects of spatial scale of soil heterogeneity on the growth of a clonal plant producing both spreading and clumping ramets. J Plant Growth Regul, 33 (2): 214-221.
Ronald H K, Terence P H, Susan R K. 1996. Density-dependent dynamics of soft coral aggregations: the significance of clonal growth and form. Ecology, 77 (5): 1592-1599.
Salzman A G. 1985. Habitat selection in a clonal plant. Science, 228 (4699): 603-604.
Slade A J, Hutchings M J.1987a. The effects of nutrient availability on foraging in the clonal herbGlechomahederacea. J of Ecol, 75(1): 95- 112.
Slade A J, Hutchings M J. 1987b. The effects of light intensity on foraging in the clonal herbGlechomahederacea. J of Ecol, 75 (3): 639-650.
Silvertown J W. 1993. Introduction to plant population.3rdedition. New York.
Takahashi M K, Horner L M, Kubota T,etal. 2011. Extensive clonal spread and extreme longevity in saw palmetto, a foundation clonal plant. Molecular Ecology, 20 (18): 3730-3742.
Wang J C, Shi X, Zhang D Y,etal. 2011. Phenotypic plasticity in response to soil moisture availability in the clonal plantEremospartonsongoricum(Litv.) Vass. Journal of Arid Land, 3 (1): 34-39.
Yan X, Wang H W, Wang Q F. 2013. Risk spreading, habitat selection and division of biomass in a submerged clonal plant: responses to heterogeneous copper pollution. Environmental pollution, 174(5): 114-120.
(责任编辑 王艳娜)
ClonalgrowthofHippophaeRhamniodesssp.sinensisattheEarlyStageinResponsetoInitialPlantingDensityandItsRegulationMechanismofBiomassAllocation
Jiang Zhun1Liu Danyi2Chen Beibei1Gao Haiyin3Liu Chunhong3Zhang Zengyue1Zou Xu1Li Genqian1
(1.ForestryCollegeofSouthwestForestryUniversityKunming650224; 2.ChinaNationalSandControlandDesertIndustryBeijing100714; 3.DingbianCountyForestryworkstation,ShannxiProvinceYulin718600)
【Objective】 The population stability ofHippophaerhamniodesssp.sinensisdepends on clonal growth ability and also on levels of environmental resources. However, the effects of the planting density are not well illustrated. This study aims to determine the optimal planting density according to the clonal growth response to the planting density and the regulation mechanism of the biomass allocation. 【Method】 Four planting densities, namely 2 500, 4 444, 10 000, and 40 000 plants·hm-2were applied. The ramet growth ability was measured for each tally tree. The clonal propagation ability was obtained by counting the daughter ramets. The clonal expansion capability was quantified by tracking the clones. Aboveground biomass was estimated from the average standard trees. The vertical and horizontal root biomasses were determined by weighting all the roots in the grid quadrats and tracked, respectively. 【Result】 The basal stem diameter and crown growth of the ramets showed logarithmic decreases with increasing planting density, while the height growth was independent of the density. The clonal propagation and expansion abilities showed parabola profiles, with synergistic effect between them. The biomass was allocated more to the belowground and less to the aboveground. The biomass allocation of horizontal roots showed a parabola. The growth ability of the ramets was positively correlated with the aboveground biomass allocation, and the clonal propagation and expansion abilities were positively correlated with the biomass allocation of the horizontal roots. 【Conclusion】 The population is traded-off in the biomass allocation between the above-and the below-ground and between the vertical and horizontal roots, and between ramet growth and the clonal propagation and expansion, by which forms individual shape and clonal growth configuration to adapt planting dnesity. With the increase of initial planting density, the ramet morphology changes from thick to medium and to slender, and the clonal propagation and expansion abilities did as a low-high-low pattern. That is to say, the clonal growth configuration went as a guerrilla-aggregation-guerrilla pattern, and the population stability exhibit a low-high-low changing process. According to this study, the suitable initial planting density is about 15 000 to 20 000 plants·hm-2, which is most conducive to the clonal growth and the population stability maintenance.
clonal propagation; clonal expansion; biomass allocation; initial planting density;Hippophaerhamniodesssp.sinensis; Mu Us sandy land
10.11707/j.1001-7488.20171004
2016-07-05;
2016-12-20。
国家自然科学基金项目(30371193、31070551、31570609);云南省林学一流学科及林学一级学科博士点建设项目(51600625)。
*李根前为通讯作者。
S718.43
A
1001-7488(2017)10-0029-11
