外源氯对花魔芋苗期生长发育及Cl-分配的影响
2017-12-01刘海利钟秀来张盛林
辜 涛,刘海利,牛 义,钟秀来,张盛林*
(1.西南大学 园艺园林学院,重庆 400716;2.教育部 南方山地园艺学重点实验室,重庆 400716;3.西南大学 魔芋研究中心,重庆 400716)
外源氯对花魔芋苗期生长发育及Cl-分配的影响
辜 涛1,刘海利2,牛 义3,钟秀来1,张盛林3*
(1.西南大学 园艺园林学院,重庆 400716;2.教育部 南方山地园艺学重点实验室,重庆 400716;3.西南大学 魔芋研究中心,重庆 400716)
【目的】为了研究源氯对花魔芋苗期生长发育及Cl-分配的影响。【方法】以花魔芋为实验材料,通过盆栽试验,研究了不同浓度的外源氯处理下花魔芋苗期病死率、光合速率、叶绿素含量、根系活力、壮苗指数等指标的变化,并探讨了外源施氯对苗期花魔芋生长发育及CI-分配的影响。【结果】随外源氯浓度的增加,花魔芋病死率呈先降后升的趋势,在16 mmol/L时最低且极显著低于对照,且各处的病死率均低于对照;花魔芋光合速率、生长速率、干物质积累速率、根系活力以及壮苗指数均随外源氯浓度的增加而呈先增后减的趋势,分别在32、4、4、8和8 mmol/L时最高,分别在64、32、32、256和128 mmol/L时显著或极显著低于对照;随外源氯浓度的增加,花魔芋叶绿素含量不断下降,在32 mmol/L时显著低于对照,在128 mmol/L时极显著低于对照;花魔芋各器官干物质分配量在外源氯浓度≤16 mmol/L时叶gt;茎gt;根,而在≥32 mmol/L时叶gt;根gt;茎;花魔芋植株和各器官氯离子含量均随外源氯浓度的增加而增加,各处理均显著或极显著高于对照,各器官氯离子含量均为根gt;叶gt;茎,各器官氯离子分配率均为叶gt;根gt;茎,高氯下分配到叶中的氯明显增加,分配到茎中的氯明显减少,而根则相对较为稳定。【结论】因此,适当施氯对花魔芋苗期生长发育无显著不良影响,甚至还有促进作用,而高氯对花魔芋具有毒害作用,显著抑制其生长发育。
花魔芋;外源氯;苗期;生长发育
【研究意义】魔芋(AmorphophallusrivieriDurieu)是天南星科(Araceae)魔芋属(AmorphophallusBlume)多年生草本植物,起源于热带雨林底层植被,主要分布在我国南部、中南半岛以及东南亚和南亚,是目前发现地唯一能大量合成葡甘露聚糖的植物[1]。花魔芋(A.konjac)以适应性广,产量高,品质好著称,为目前魔芋主栽种。氯是植物必需的微量元素,在植物体内,氯仅有少部分参与生理生化反应,大部分是以活性很强的Cl-形态存在,参与电荷补偿和渗透调节[2]。适量施氯可促进蔬菜作物生长发育,抑制病害发生,提高光合速率等,进而提高产量和改善品质,而过高的氯破坏细胞的超微结构,从而产生毒害作用,严重抑制作物的生长发育[3]。因此,开展花魔芋氯素营养研究和毒害机理研究,对了解外源施氯对苗期魔芋生长发育的影响具有重要作用和理论意义。【前人研究进展】迄今为止国内外研究者们已经对多种作物开展了有关氯素营养和毒害作用的研究,认为不同作物以及同种作物不同品种对氯敏感性不同,一般而言,作物仅在某一特定生育时期对氯比较敏感,敏感期过后即使氯浓度偏高植株生长也会基本恢复正常。对大多数作物而言,氯的敏感期主要在苗期如小麦、大麦等主要在2~5叶龄期,大白菜、小白菜等主要在4~6叶龄期,而柑橘、茶树等主要在1~4年的幼龄期等[2]。植物的生长特征是植物在外部形态上对氯素营养功能和毒害作用的响应,而植物生理生化反应则是植物内部活动对氯素营养功能和毒害作用的响应,众多研究表明,在一定范围内施氯对植物的生长发育具有促进作用,而高浓度的氯则相反,张兆辉[4]、刘善江[5]、戚冰洁[6]、曾睿[7]、耿贵[8]、汪吉东[9]等均研究表明,一定范围的外源氯处理有利于促进植株生物量、株高、叶面积、茎粗的增加,有利于提高植株的存活率、光合速率和根冠比,有利于促进植株干物质的积累等,而高浓度的氯处理则相反。此外张兆辉等[4,6,9]研究发现,植株体内氯离子含量均随外源氯浓度的增加而增加,但氯离子的分配、分布与植物种类、品种以及生长发育期和施用时间长短有关。但施氯对植物叶绿素含量的影响目前看法不一,有的研究表明施氯不利于叶绿素的合成和积累,而有的研究表明适量施氯有利于促进植物叶绿素的合成和积累[5-6]。【本研究切入点 】在不同外源氯浓度处理下,花魔芋的形态指标、动态生长指标、生理生化指标等都可能受到影响,因此在开展花魔芋苗期氯素营养和毒害机理研究时,仅仅通过其中任何单一指标来对其进行研究和评价,均不准确,不科学,应综合各方面原因进行考虑,综合分析。魔芋属薯芋类作物,而薯芋类作物又是传统上的忌氯作物,且含氯化肥具有肥效高、价格便宜等优点,因此研究魔芋氯素营养和毒害机理具有重要意义, 但有关魔芋苗期氯素营养和毒害机理的研究目前尚未见报道。【拟解决的关键问题】本试验采用大棚盆栽方式,研究了氯对魔芋苗期生长发育的影响,有利于了解氯对魔芋的营养生理功能和氯对魔芋的毒害作用,为魔芋科学施用含氯化肥提供理论基础。
1 材料与方法
1.1 材料
大小一致的花魔芋(约10 g)种球茎(西南大学魔芋研究中心提供)、珍珠岩(沂水沂蒙珍珠岩厂提供)和蛭石(石家庄永恒建材有限公司提供)。
1.2 实验设计
试验于2015年5月至7月在西南大学魔芋研究中心歇马实验基地进行。试验采用大棚盆栽方式,花盆直径50 cm,高40 cm。基质采用珍珠岩、蛙石2∶1混合,浇营养液,出叶后进行遮阴、常规管理。营养液采用Hongland配方,配方中氯化钴用等量的硫酸钴代替,不同浓度外源氯水平由氯化铵调节,营养液中的氮由硝酸铵调节,保持同一氮水平。试验设8个氯水平处理:CK(0 mmol/L)、处理1(4 mmol/L)、处理2(8 mmol/L)、处理3(16 mmol/L)、处理4(32 mmol/L)、处理5(64 mmol/L)、处理6(128 mmol/L)、处理7(256 mmol/L),每个处理3次重复,每个重复3盆,每盆10株。种芋于5月10日浸种、催芽,5月25日(芽长2 cm左右)移栽到花盆中,每5 d浇1次营养液(浇透:每盆约2 L)。
1.3 测定方法
于2015年6月15、20、25日分别记录各处理病死数;6月25日分别测定各处理的光合速率、叶绿素含量、株高、叶柄粗和根系活力,采用CB-1101光合仪测定各处理植株相同位置的第2片功能叶(从顶叶到小叶叶柄数)的光合速率,采用混合提取法[10]测定叶绿素含量,采用TTC还原法[11]测定根系活力,用直尺测定其株高(叶柄基部到小叶叶柄基部),用游标卡尺测定叶柄粗(主叶柄基部);6月25日采集各处理植株地上部叶以及地下部茎和根,分别在105 ℃杀青15 min,然后转移到75 ℃下烘干,烘干后分别测定其干重和氯离子含量,氯离子含量采用改进莫尔法[12]。生长速率=株高/(移栽后到6月25日测定时的天数);干物质积累速率=植株干重/(移栽后到6月25日测定时的天数);壮苗指数=(叶柄粗/株高+根干重/茎叶干重)×全株干重。

表1 不同浓度外源氯处理对花魔芋苗病死率的影响
注:同一行数据间不同小写字母/大写字母表示处理间差异显著(Plt;0.05)/极显著(Plt;0.01),下同。
Note: Different lowercase letters (Capital letters) in the same line mean significant difference at 5 % level (extremely significant difference at 1 % level). The same as below.
采用SPSS22.0和Excel软件进行试验数据处理和分析。
2 结果与分析
2.1 不同浓度氯处理对花魔芋病死率的影响
由表1可知,花魔芋病死率随外源氯浓度的增加呈现为先降后升的趋势,其病死率在外源氯浓度为16 mmol/L时最低(5.0 %)且极显著低于对照;花魔芋在外源氯浓度为8~128 mmol/L时,施氯处理病死率均显著或极显著低于对照,在256 mmol/L时低于对照但不显著。
2.2 不同外源氯处理对花魔芋光合速率(Pn)和叶绿素(Chl)含量的影响
由表2可以看出,花魔芋叶绿素含量均随外源氯浓度的增加而降低。在外源氯浓度≤16 mmol/L时,花魔芋叶绿素含量各处理间无显著性差异;与对照相比,其叶绿素含量在32 mmol/L时显著低于对照,在128 mmol/L时极显著低于对照。
花魔芋光合速率随外源施氯浓度的增加而表现为先增后降的趋势,在32 mmol/L外源氯处理下达到最大值6.50,且极显著高于对照。当外源氯浓度≥64 mmol/L时,其光合速率极显著下降且极显著低于对照,且外源氯浓度越高下降越显著。
2.3 不同Cl-浓度处理对魔芋根系活力影响
由表3可知,随外源氯浓度的增加,花魔芋根系活力表现为先升后降的趋势,处理2(8 mmol/L)最高。外源氯浓度为4~128 mmol/L时,其根系活力均极显著高于对照,且各处理间差异极显著,而在256 mmol/L时极显著低于对照。
2.4 不同浓度氯处理对花魔芋生长状况的影响
随外源氯浓度的增加,花魔芋生长速率、干物质积累速率和壮苗指数均呈先升后降的趋势,分别在4、8和8 mmol/L 时达到最高值,且均显著或极显著高于对照(表4)。在外源氯浓度分别在128、16和16 mmol/L 时,其生长速率、壮苗指数和干物质积累速率均显著或极显著下降;与对照相比,其生长速率、干物质积累速率以及壮苗指数分别在外源氯浓度为8、32和32 mmol/L时开始低于对照,而分别为16、32和64 mmol/L时显著或极显著低于对照。

表2 不同浓度外源氯处理对花魔芋光合速率和叶绿素含量的影响

表3 不同Cl-浓度处理对花魔芋根系活力的影响

表4 不同浓度外源氯处理对花魔芋生长速率、干物质积累速率以及壮苗指数的影响
2.5 不同Cl-浓度处理对魔芋干物质分配的影响
由表5可知,花魔芋叶和根干物质分配率随外源氯浓度的增加而呈先降后升的趋势,其茎则表现为先升后降的趋势。在处理2(8 mmol/L)时,花魔芋叶和根干物质分配率达到最小值,其茎干物质分配率则达到最大值,且均显著差异于对照;其叶干物质分配率在16 mmol/L时极显著升高,在32 mmol/L时显著高于对照,在≥64 mmol/L时极显著高于对照;其根干物质分配率在0~16 mmol/L时各处理间无显著性差异,而在32~128 mmol/L时极显著高于对照,但处理间差异不显著;其茎干物质分配率在4~8 mmol/L时极显著高于对照,在16 mmol/L时显著下降,而与对照无显著性差异,在≥32 mmol/L时极显著低于对照。
花魔芋通过光合作用和从种球茎转化而来的营养物质在≤16 mmol/L时主要分配到地上部叶和地下部茎中,分别占50 %和20 %以上,根中分配较少且相对较为稳定;在外源氯浓度≥32 mmol/L时则其主要分配到叶和根中,分别占60 %和19 %以上,且在茎和叶的变化幅度较大,根则相对较为稳定。
2.6 不同Cl-浓度处理对魔芋不同器官中氯离子含量的影响
花魔芋植株及各器官氯离子含量均随外源氯浓度的增加而增加,根氯离子含量最高,其次是叶,茎最少(表6)。外源氯处理下,花魔芋植株和各器官氯离子含量均显著或极显著高于对照;花魔芋植株、叶和根在≥16 mmol/L时,各处理间差异极显著;其茎在≥64 mmol/L时,各处理间差异不显著,说明高氯下花魔芋茎氯离子含量相对较为稳定。
由表7可知,不同浓度外源氯处理下,花魔芋吸收的氯主要分布在叶中(40 %以上),其次为根(29 %以上),茎(23 %以下)中分配最少;花魔芋分配到叶中的氯随着外源氯浓度的增加而增加,分配到茎中的氯则表现为先增后减的趋势且浓度越高下降趋势越明显,而根中氯离子分配率则相对较为稳定,说明高氯下,花魔芋吸收的氯主要分配到了叶中,对叶的影响较大。
3 讨 论
植物的生长发育与激素和酶活性密切相关,植物激素调控细胞分裂和生长,调节和控制个体发育,而植物体内的各种酶则起着调控物质和能量的形成以及转化,为植株个体的生长发育提供能量、营养物质和结构物质等[13]。在植物体内,氯参与某些激素的组成(如4-氯-吲哚-3-乙酸)和合成(如ABA、IAA、GA3、ZR以及CH2等)[14-15];在植物体内,某些酶类必需有Cl-的存在和参与才可能具有活性,如淀粉酶、液泡膜上H+-ATP酶、硝酸还原酶、蔗糖酶以及SOD等[2]。但当外源氯离子过量时,就可能对作物产生氯毒害,破坏细胞的超微结构(如破坏线粒体基质和脊结构以及细胞膜系统等),导致植株生长缓慢、矮小甚至死亡[16-17]。郑青松等[18]研究表明,外施氯6.25~50 mmol/L 可显著促进番茄生长,显著促进其干物质积累,而在200和300 mmol/L时幼苗干物质积累速率极显著下降;曾睿等[7]报道,不同的供氯水平对烤烟的株高影响显著,供氯水平为90 kg/hm2时株高最高,与对照差异显著;郑青松等[19]用不同浓度外源氯处理向日葵幼苗8d,各处理幼苗生长速率均显著下降,其干物质积累速率在6.25~25 mmol/L时显著高于对照,而200~300 mmol/L时则极显著下降。在本试验中,与对照相比,0~16 mmol/L外源氯处理对花魔芋的生长速率、壮苗指数和干物质积累速率均无显著不利影响甚至还有一定的促进作用,高氯(≥64 mmol/L)严重抑制了花魔芋的生长发育,显著或极显著降低其生长速率、壮苗指数和干物质积累速率,这与前人研究结果相一致,说明适量施氯有助于壮苗,有利于物质的合成和转化。除处理1(4 mmol/L)外,苗期花魔芋生长速率均随外源氯浓度的增加而有不同程度的降低,这与郑青松等研究结果不一致,这可能由处理浓度和材料的不同引起的。
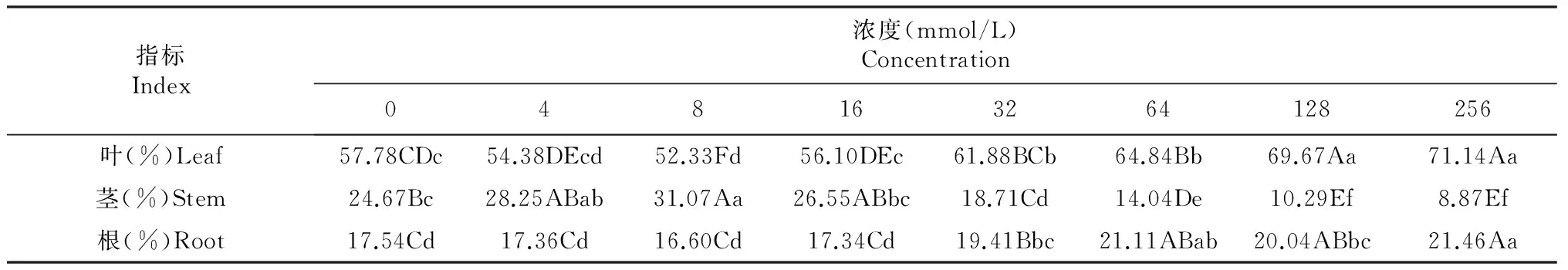
表5 不同浓度氯处理下,花魔芋根、茎和叶干物质分配情况

表6 不同浓度外源氯处理对花魔芋植株及各器官氯离子含量的影响
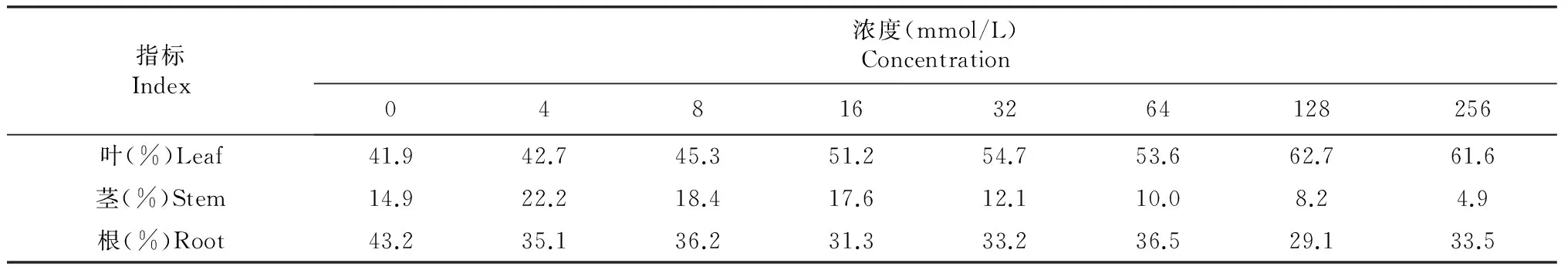
表7 不同浓度外源氯处理下,氯在植株不同器官的分配情况
氯对植物病害的发生具有明显的抑制作用,施氯可以减轻10多种作物的根叶病害[2],降低马铃薯空心病、褐心病的发生[17]。Sanogo和Yang[20]研究认为,钾素供应充足时,氯素较钾素更能抑制土传病害大豆碎死症的发生;Heckman等[21]发现在钾施用量相同的情况下,与施用硫酸钾相比,氯化钾对茎腐病的免疫效果达50 %以上。本研究中,花魔芋病死率随外源施浓度的增加而呈先降后升的趋势,在8~256 mmol/L 外源氯处理下,其病死率显著或极显著低于对照,这与前人研究结果相一致,说明施氯对花魔芋病害的发生有明显的抑制作用,而过高的氯可加剧病害的发生,这可能是由于过高的氯导致其生理紊乱,抗逆性下降造成的。
氯参与光合作用,促进光合磷酸化和ATP的合成,直接参与光系统Ⅱ氧化位上的希尔反应[2];氯参与气孔运动调节,气孔张开时,保卫细胞液胞中的Cl-浓度提高,反之Cl-浓度就会下降,这与植物表皮细胞中Cl-的移动有关[3,17]。李廷轩等[22]研究认为,氯离子能通过调节气孔开闭来间接影响光合作用;胡小婉等[23]研究表明,低氯短期处理对油菜幼苗光合速率、气孔导度和叶绿素含量均具有促进作用,高氯处理不管短期还是长期处理均显著降低其光合速率、叶绿素含量以及气孔开度;俏丽等[24]研究表明,低氯对白菜叶绿素含量无显著影响,中氯和高氯显著降低叶绿素含量。本试验中,花魔芋叶绿素含量随外源氯浓度的增加而降低,低氯对其叶绿素含量无显著影响,高氯显著降低其叶绿素含量;低氯对花魔芋光合速率具有促进作用,而高氯极显著降低其光合速率,这与前人研究结果相一致;高氯下,花魔芋光合速率下降幅度明显大于叶绿素含量,说明高氯影响光合速率下降的主要因素不是叶绿素含量的下降,这可能是由于高氯导致气孔减小或关闭引起的[23]。
俏丽等[24]研究表明,低浓度氯处理对白菜幼苗根系活力无显著性影响,中氯和高氯均显著降低其根系活力;周益等[25]研究认为,施适量的氯对烤烟根系活力有促进作用,施氯过多烤烟根系活力有明显的抑制作用。在本研究中,与对照相比,低氯和较高浓度的氯均极显著促进花魔芋的根系活力,而高氯(256 mmol/L)对其根系活力起极显著的抑制作用,这与周益的研究结果相一致,而与俏丽的研究结果不完全一致,这可能是材料的基因型差异引起的。说明根系活力的维持需要较高浓度的氯,根的耐氯能力较高。
氯是必需元素,不足或过多均影响作物的生长发育,影响光合效率和光合产物的运输,当氯浓度高于300 mg/kg 时,马铃薯CO2同化量降低,光合产物向其块茎运输速率下降20 %~40 %,导致块茎小而少[2]。马国瑞[26]等研究认为,随着外源氯浓度增加,块茎、茎和叶片中的光合产物减少,尤其是块茎和茎,而当≥300 mg/L时,其光合产物大幅度的减少,而叶片中光合产物的放射性活下降的幅度则相对较低。在本试验中,花魔芋光合产物以及从种芋中吸收转化而来的营养物质不管是低氯还是高氯均主要分配到叶中,其余部分的营养物质在低氯下主要分配到茎中,而在高氯下则主要分配到根中,这与前人研究结果相一致,说明低氯有利于营养物质的合理分配,高氯严重抑制营养物质向茎中分配,这可能是因为高浓度的氯严重抑制了花魔芋光合产物的形成以及其他营养物质的转化,导致总营养物质严重不足,营养物质主要供应叶、根等营养器官。
氯素在植株体内的分布, 不论种类和种植茬次,均主要分布在叶和茎等营养器官, 其含量占80 %以上[27];在王芳等[28]对西瓜的研究中,氯在地上部的分布为根部叶gt;茎干gt;顶部叶gt;果实;唐年鑫等[27]研究认为,氯在棉株各部位的分布为:叶gt;铃gt;壳gt;茎gt;根gt;棉籽gt;纤维;在油菜幼苗的研究中,不同浓度处理下,其体内不同器官氯含量分布不同,其中叶柄和叶中含量较高,且逐渐上升,根中最低[23]。本试验研究表明,花魔芋植株氯离子含量均随外源氯浓度的增加而显著或极显著升高;花魔芋植株及各器官氯离子含量均随外源氯浓度的增加而增加,根中氯离子含量最高,其次是叶,茎最少且高氯下较为稳定;不同浓度外源氯处理下,花魔芋吸收的氯主要分配到叶中(40 %以上),其次为根(29 %以上),茎(23 %以下)中分配最少;外源施氯对花魔芋茎叶中氯离子的分配有较大的影响,对根的影响较小,尤其是高氯下,花魔芋叶中氯离子分配率显著增加,而茎则显著或极显著降低,这与前人研究结果相一致,这说明外源氯浓度是花魔芋吸收氯的主要因素,根吸收的氯主要分配到叶中,茎中最少;对照组茎和根与处理1有明显的差异,对照的茎氯离子分配量明显降低,而根则明显升高,叶的差异不明显,这可能是因为对照组花魔芋植株从种球茎中获得的氯不足以满足植株的生长,把吸收的氯主要分配到叶和根,促进营养器官的生长。
4 结 论
适量施氯有利于促进苗期花魔芋的生长发育,有利于提高其生长速率、壮苗指数、干物质积累速率、光合速率和根系活力,而高浓度的氯则相反。外源施氯不利于苗期花魔芋叶绿素的合成和积累,高浓度的氯显著降低其叶绿素含量。在花魔芋苗期,低氯有利于促进营养物质运输和合理分配,高氯严重抑制营养物质向茎中分配和运输。在花魔芋苗期,适量施氯对花魔芋病害的发生有均明显的抑制作用,而过高的氯可加剧病害的发生。苗期花魔芋对外源氯的吸收、运输和积累均与外源氯浓度呈显著正相关,且吸收的氯主要分配到地上部叶中,其次是根,地下茎中最少。
[1]刘佩瑛. 魔芋学[J]. 中国. 北京: 农业出版社, 2004: 1-30.
[2]黄建国. 植物营养学[J]. 北京: 中国林业出版社, 2004.
[3]张兆辉, 汪李平, 陈龙英. 氯在蔬菜作物上的研究进展[J]. 北方园艺, 2010(8): 225-229.
[4]张兆辉, 汪李平, 王 林,等. 外源氯对西瓜幼苗生长及离子分配的影响[J]. 华中农业大学学报, 2009, 28(3):316-319.
[5]刘善江, 马笑蕊. 不同氯离子浓度对黄瓜生长的影响[J]. 农业新技术, 2000, 18(6):31-32.
[6]戚冰洁, 曹月阳, 张珮琪,等. 外源氯胁迫对甘薯幼苗光合特性的影响[J]. 西北植物学报, 2013, 33(5):984-991.
[7]曾 睿, 董文汉, 何忠俊,等. 不同施氯水平对云南烤烟生长、产量和品质的影响[J]. 中国农学通报, 2009, 25(21):212-216.
[8]耿 贵, 周建朝, 陈 丽,等. 氯化钠胁迫下甜菜不同品种(系)种子发芽率和幼苗生长的差异[J]. 中国糖料, 2004(2):14-18.
[9]汪吉东, 郑青松, 刘兆普,等. 外源氯对库拉索芦荟生长及品质的影响[J]. 西北植物学报, 2006, 26(6):1201-1205.
[10]李合生. 植物生理生化实验原理和技术[M]. 高等教育出版社, 2000.
[11]宗学凤. 植物生理研究技术[M]. 重庆:西南师范大学出版社, 2011.
[12]高 岩, 高玉珍. 莫尔法测定烟叶氯含量的前处理方法改进试验[J]. 商场现代化, 2005(3):158-159.
[13]王 忠.植物生理学[J].中国.北京:中国农业出版社,2008,9:303-366
[14]李海云, 王秀峰, 魏 珉,等. 含氯化肥对黄瓜叶片激素含量及叶绿体超微结构的影响[J]. 园艺学报, 2003, 30(5):598-600.
[15]马国瑞,含氯化肥使用新技术[M].杭州:浙江科学技术出版社,1994:2-59.
[16]卢红霞. 氯(Cl-)对马铃薯的某些生理效应及对土壤中氮肥行为的影响[D]. 浙江大学, 2001.
[17]毛知耘.中国含氯化肥[M].中国.北京:中国农业出版社,2001.
[18]郑青松, 杜 爽, 刘兆普,等. 外源氯对番茄幼苗生长及养分吸收、利用的影响[J]. 园艺学报, 2006, 33(4):849-852.
[19]郑青松, 杨文杰, 刘兆普,等. 外源氯处理对向日葵幼苗生长、养分吸收及植株硝态氮含量的影响[J]. 植物营养与肥料学报, 2007, 13(6):1161-1165.
[20]Sanogo S, Yang X B. Relation of sand content, pH, and potassium and phosphorus nutrition to the development of sudden death syndrome in soybean[J]. Canadian Journal of Plant Pathology, 2001, 23(2): 174-180.
[21]Heckman J R. Corn stalk rot suppression and grain yield response to chloride[J]. Journal of Plant Nutrition, 1998, 21(1): 149-155.
[22]李廷轩, 马国瑞. 含氯化肥的研究进展[J]. 西南农业学报, 2002, 15(2):86-91.
[23]胡小婉. 氯对油菜生长与营养吸收利用的效应及其机制[D]. 南京农业大学, 2013.
[24]肖 丽, 孙宁波, 隋方功. 氯对白菜幼苗的生长及养分吸收的影响[J]. 植物营养与肥料学报, 2008, 14(3):608-612.
[25]周 益. 闽西贫氯烟区施氯对烤烟生理特性和产质量影响[D]. 湖南农业大学, 2009.
[27]唐年鑫, 沈金雄, 陈少三. 作物对氯素吸收利用与分布特点的研究[J]. 核农学通报, 1996(6):273-277.
[28]王 芳. 氯化钾对西瓜产量品质和土壤中氯离子残留的影响[D]. 中国农业科学院, 2006.
(责任编辑 李山云)
EffectofDifferentConcentrationsofExogenousChlorineonGrowthandChloride’sDistributionofAmorphophalluskonjacinSeedlingStage
GU Tao1, LIU Hai-li2, NIU Yi3, ZHONG Xiu-lai1, ZHANG Sheng-lin3*
(1.College of Horticulture and Landscape Architecture, Southwestern University, Chongqing 400716,China ;2. Key Laboratory of Horticulture Science for Southern Mountainous Regions, Ministry of Education, Chongqing 400716,China;3.Key Laboratory of Olericulture Konjac Research Center, Southwestern University ,Chongqing 400716,China)
【Objective】The paper aims to study the effect of different concentrations of exogenous chlorine on growth and chloride’s distribution of theAmorphophalluskonjacin seedling stage. 【Method】Taken theAmorphophalluskonjac-konjacas tested materials, under different concentrations of exogenous chlorine ,the case-fatality rate ,the photosynthetic rate ,the chlorophyll content ,the root activity and the sound seedling index were studied by pot experiments, and the effects of the different concentrations of exogenous chlorine on growth and distribution of chloride ion of theAmorphophalluskonjacin seedling stage wereinvestigate. 【Result】The results showed that: Along with the increase of chlorine concentration, the case-fatality rate of theA.konjacshowed down first and then up, and when the concentration of chloride ion is at 16 mmol/L, the case-fatality rate is the lowest and it is most significantly lower than the control group. In various treatment, the case-fatality rate of theA.konjacwere lower than control group. Along with the concentrations of exogenous chlorine increasing, the photosynthetic rate, growth rate, dry matter accumulation rate, root activity and sound seedling index of theA.konjacincreased firstly and then decrease. When the concentrations of exogenous chlorine respectively were at 32, 4, 4, 8, and 8mmol/L, they reached a maximu value. When the concentrations of exogenous chlorine respectively were at 64, 32, 32, 256 and 128 mmol/L, they were significantly or very significantly lower than the control group. The chlorophyll content decreased continuously along with the increase of chlorine concentration. It's was significantly lower than the control group at 32 mmol/L and was very significantly lower than the control group at 128 mmol/L. When the chlorine concentration is less than or equal to 16 mmol/L , the dry matter’s distribution rate of various organs were: the leafgt;the stemgt;the root. However, when it is greater than or equal to 32 mmol/L, they were: the leafgt;the rootgt;the stem. The chloride ion content of individual and various organs increase with increasing chloride ion concentration, and the all treatments were significantly or very significantly higher than the control, and the chloride ion content of various organs was: the root gt;the leafgt;the stem. In the high concentration chlorine element treatments ,the Chlorine amount was increased significantly in the leaf , was dropped markedly in the stem, and was stabilized in the root relatively. 【Conclusion】Therefore, the proper chlorine fertilizer application had no effect on growth' of theA.konjacin seedling stage, and even to promote it. But high levels of exogenous chlorine was toxic to theA.konjacand restrained significantly its’ growth.
A.konjac; Exogenous chlorine; Seedling stage; Growth
S53
A
1001-4829(2017)11-2562-06
10.16213/j.cnki.scjas.2017.11.030
2016-10-27
重庆市“121”科技支撑示范工程(104190 /40605014)
辜 涛(1989-),男,四川眉山人,在读研究生,研究方向:蔬菜栽培生理及设施园艺, E-mail:532964250@qq.com,*为通讯作者:张盛林(1963-),男,四川成都人,研究员,硕士,主要从事魔芋种质资源研究与利用工作,E-mial:konjac@163.com。
