马尾松优树种子表型变异规律研究
2017-12-01,,,
, , ,
(1.贵州大学林学院, 贵阳 550025; 2.贵州大学贵州省森林资源与环境研究中心, 贵阳 550025; 3.贵州都匀国家马尾松良种基地, 贵州 都匀 558000)
·问题探讨·
马尾松优树种子表型变异规律研究
王好运1,2,朱小坤1,2,谢维斌3,吴峰1,2
(1.贵州大学林学院, 贵阳 550025; 2.贵州大学贵州省森林资源与环境研究中心, 贵阳 550025; 3.贵州都匀国家马尾松良种基地, 贵州 都匀 558000)
以贵州都匀马尾松良种基地基因收集区内优树单株所采集的种子为材料,测定种子形态、品质和营养物质含量等性状,剖析马尾松种子在单株间及单株内的变异规律。结果表明:不同马尾松单株优树间种子形态特征、品质指标、可溶性糖、可溶性蛋白质及脂肪含量存在显著差异。其中籽粒较大的种子拥有较高的萌发能力,发芽率最高达96.17%,内含物质主要以脂肪为主,占种子总重量的55%左右。种子形态指标与千粒重、可溶性糖和可溶性蛋白质呈显著正相关;种子发芽指标与可溶性蛋白质、脂肪含量呈显著正相关;可溶性蛋白质与脂肪含量呈显著正相关。研究表明,马尾松种子形态特征、萌发能力和营养物质含量在单株水平上存在一定的选择潜力,进行种子园的母树选择时,可结合种子表型性状进行综合考虑。
马尾松; 优树; 种子性状; 萌发能力;
种子是有性繁殖植物生活史中重要的环节,关系到物种的延续、进化。种子的大小、外部形态特征、内含物种类和含量等特性,在不同的物种间和同一物种内部都存在着变异,这些变异都是植物适应周围环境而自然选择的结果。而种群内种子大小的变异主要来源于各个个体间,个体间种子大小变异既受遗传控制,又受环境的影响[1]。大量的研究发现,种子大小、千粒重及内含物含量对植物生长发育的早期阶段有较明显影响,主要体现在对发芽率、幼苗大小、幼苗竞争力及相对生长速率等方面,种子形态、千粒重及内含物含量的变异均对子代苗期生长有明显的影响[2-5]。
马尾松(Pinusmassoniana)系松科松属,是主要的产脂、纸浆用材树种,也是我国南方主要的造林树种之一[6]。已有研究表明,不同地理种源马尾松种子的发芽特性存在差异[7],种子内含物含量会影响到种子的萌发速率及生长特性[8]。但关于马尾松种子表型性状在单株间及单株内变异方面的研究较为缺乏。对马尾松种子形态、品质和营养物质含量等性状在单株间变异规律进行的研究,可为马尾松种子园母树选择及良种繁育提供理论依据。
1 材料与方法
1.1 试验地概况
试验地位于贵州省都匀市马鞍山林场国家马尾松良种基地的基因收集区,地处北纬26°16′,东经107°31′,海拔1 000 m左右,为中亚热带湿润气候,年均温15.8 ℃,年平均降水量约1 400 mm。年平均相对湿度79%,全年日照时数1 145.5 h,生长期达300 d以上。
1.2 试验材料
本研究所用马尾松种子来自于基因收集区内随机挑选的9个优树单株,全部为自由授粉,依次编号K 1、H 12、D 53、H 50、H 36、D 126、K 3、D 8、H 18。于2015年11月进行单株球果收集,球果经烘干脱粒后,收集种子置于低温密封条件备用。
1.3 研究方法
1.3.1 种子形态指标测定
采用WinSEEDLE种子和针叶图像分析系统对种子形态进行测定,每株优树随机选取100粒种子测定弧长、弧宽、体积等形态指标,重复4次。
1.3.2 种子品质指标的测定
种子千粒重、发芽率、发芽势以及发芽指数的测定参照GB 2772-1999《林木种子检验规程》[9]完成,每处理重复4次。
表1 马尾松优树种子外部形态统计分析结果

种子性状平均值变异幅度标准差F值单株间单株内变异系数(%)弧长(mm)5.353.71~6.930.53635.51∗∗0.839.91弧宽(mm)2.981.97~3.720.29284.41∗∗0.889.73体积(m3)21.577.69~38.335.65665.99∗∗1.3026.19长宽比1.791.23~2.400.20124.36∗∗1.1511.17
注:“*”p<0.05;“**”p<0.01。
表2 马尾松不同单株优树种子的形态特征

编号 弧长(mm) 弧宽(mm) 长宽比 体积(mm3) 均值±标准差CV(%)均值±标准差CV(%)均值±标准差CV(%)均值±标准差CV(%)K15.46±0.32c5.843.10±0.21c6.891.74±0.11e6.0723.81±4.01c16.84H125.35±0.31d5.682.80±0.20g7.271.89±0.11a6.0118.64±3.21g17.24D535.57±0.40b7.233.04±0.25d8.241.83±0.14cd7.6922.67±4.45d19.64H505.50±0.40c7.312.97±0.25e8.451.84±0.12b6.5821.86±4.69e21.44H364.63±0.26h5.652.68±0.18h6.731.71±0.10f5.9814.95±2.43i16.25D1265.28±0.35e6.552.91±0.21f7.151.81±0.12d6.7519.41±3.29f16.97K36.06±0.36a5.893.28±0.22a6.611.82±0.11cd6.1728.81±4.49a15.60D85.59±0.33b6.013.25±0.18b5.461.71±0.10f5.8727.18±3.89b14.31H184.80±0.34g7.062.82±0.22g7.741.68±0.11g6.4717.58±2.86h16.27
注:不同字母表示差异显著。
表3 马尾松不同单株优树种子的营养物质含量与千粒重比较
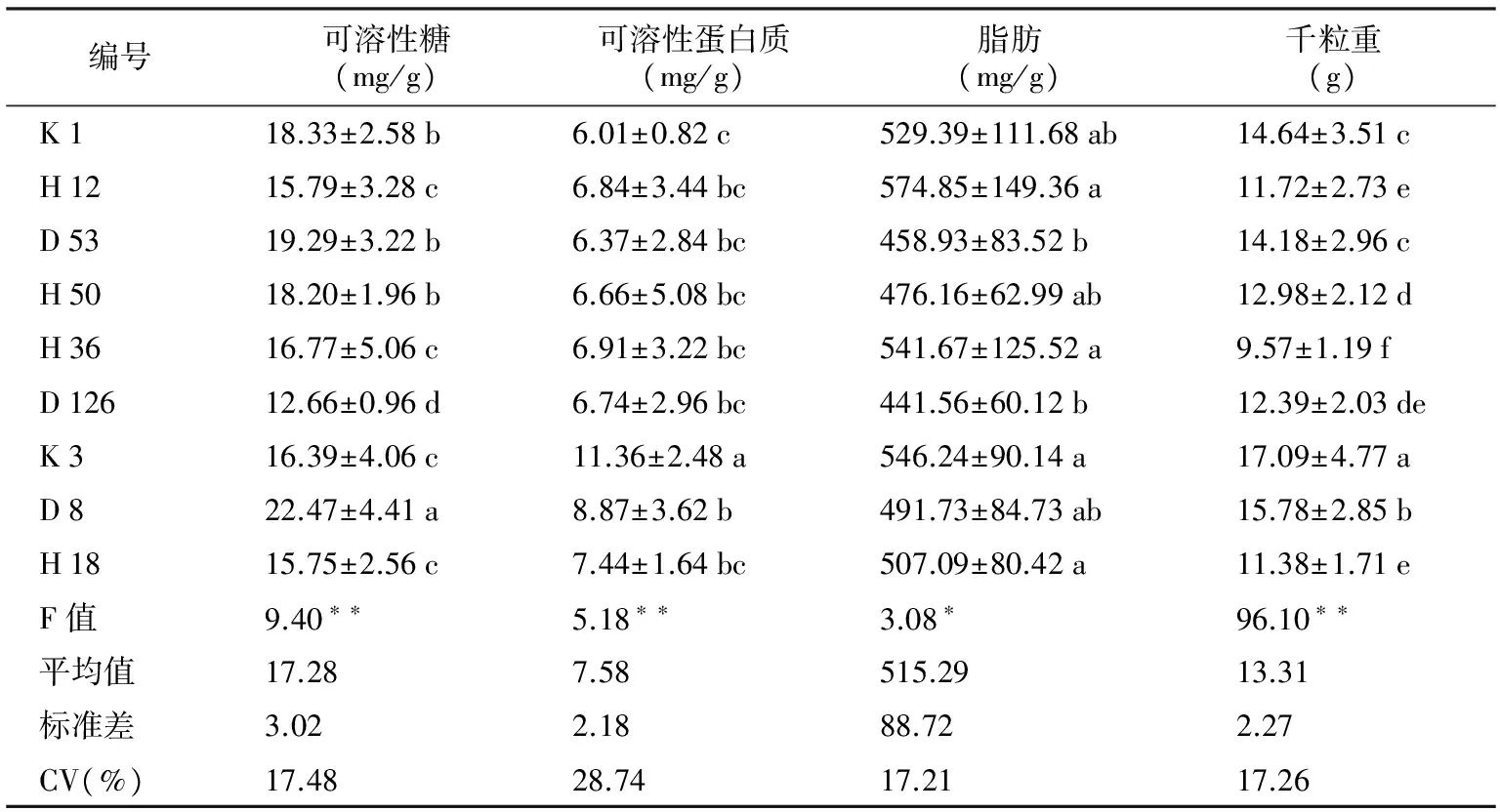
编号可溶性糖(mg/g)可溶性蛋白质(mg/g)脂肪(mg/g)千粒重(g)K118.33±2.58b6.01±0.82c529.39±111.68ab 14.64±3.51cH1215.79±3.28c6.84±3.44bc574.85±149.36a 11.72±2.73eD5319.29±3.22b6.37±2.84bc458.93±83.52b 14.18±2.96cH5018.20±1.96b6.66±5.08bc476.16±62.99ab 12.98±2.12dH3616.77±5.06c6.91±3.22bc541.67±125.52a 9.57±1.19fD12612.66±0.96d6.74±2.96bc441.56±60.12b 12.39±2.03deK316.39±4.06c11.36±2.48a546.24±90.14a 17.09±4.77aD822.47±4.41a8.87±3.62b491.73±84.73ab 15.78±2.85bH1815.75±2.56c7.44±1.64bc507.09±80.42a 11.38±1.71eF值9.40∗∗5.18∗∗3.08∗ 96.10∗∗平均值17.287.58515.29 13.31标准差3.022.1888.72 2.27CV(%)17.4828.7417.21 17.26
注:平均值±标准差,小写字母表示差异显著,“*”p<0.05;“**”p<0.01。下同。
1.3.3 种子营养成分的测定
种子可溶性糖含量采用蒽酮显色法测定[10];种子可溶性蛋白含量用考马斯亮蓝法进行测定[10];种子脂肪含量采用索氏提取法测定[11],以石油醚(30~60 ℃)为提取溶剂,重复4次。
1.4 数据处理
利用SPSS 20.0软件对试验数据进行GLM分析、Duncan法多重比较、相关性分析。
2 结果与分析
2.1 马尾松优树种子形态变异
从表1可看出,不同单株间,马尾松种子弧长、弧宽、体积及长宽比的差异达极显著水平,而单株内种子形态未呈显著差异。马尾松种子体积表型变异最大,变异系数达26.19%,种子的弧长、弧宽及长宽比变异系数均在10%左右。
不同单株优树种子的形态特征显示(表2),K 3的单株种子最大,其弧长、弧宽及体积分别为6.01,3.31 mm及28.81 mm3,均显著高于其他单株。长宽比可作为种子形态的指标,种子长宽比变化范围为1.68~1.89,最大的是H 12,最小的是H 18。H 36的单株种子最小,其体积仅为K 3的52%,弧长和弧宽是K 3的80%左右。
2.2 马尾松优树种子营养物质含量变异
由表3可知,马尾松种子内部脂肪含量较高,达515.29 mg/g,远大于可溶性糖及可溶性蛋白质含量。在不同单株之间,种子蛋白质含量变异系数最大,为28.74%,可溶性糖和可溶性蛋白质含量的变异系数均在17%左右。种子可溶性糖、可溶性蛋白质及脂肪含量在各单株间的差异达极显著(p<0.01)或显著(p<0.05)水平。种子可溶性糖含量最高的单株是D 8(22.47 mg/g),最小的是D 126(12.66 mg/g)。可溶性蛋白质含量最大的单株是K 3(11.36 mg/g),最低的是K 1(6.01 mg/g)。H 12的单株种子脂肪含量最高,为574.85 mg/g,D 126单株含量最少,为441.56 mg/g。不同马尾松单株优树种子千粒重平均值为13.31 g,其中K 3种子千粒重最大,H 36号种子最小,前者是后者的1.78倍,变异系数为17.26%。
2.3 马尾松优树种子萌发能力变异
结果(表4)表明,不同马尾松单株优树种子的发芽率、发芽指数、发芽势及千粒重均存在极显著差异(p<0.01)。各单株优树种子发芽率平均值为89.08%,其中发芽率最大的是K 3(96.17%),最小的是D 53(71.59%),前者是后者的1.34倍,变异系数为10.17%;发芽指数可以反映种子在萌发过程中速率的大小。各单株优树种子的发芽指数平均值为34.48,略高于对照组,发芽指数最高的是H 36(41.65),最低的是D 53(21.49),变异系数为17.99%。各单株优树种子发芽势平均变异系数高达29.41%,发芽势最高的是H 36(85.13%),最低的是D 53(37.49%),前者是后者的2.27倍。
表4 马尾松不同单株优树种子萌发能力

编号发芽率(%)发芽指数发芽势(%)K196.07±4.04a38.29±4.16cd75.52±20.14abH1293.24±6.12ab36.61±8.34bcd74.35±28.02abD5371.59±11.55d21.49±8.04a37.49±18.58cH5082.31±12.86c33.24±8.18bc57.82±23.69bcH3696.03±8.02a41.65±7.26d85.13±14.04aD12685.36±15.01bc33.28±4.32bc59.72±8.86bcK396.17±9.24a37.47±4.96cd71.13±11.08aD892.43±3.82abc36.62±3.36bcd55.61±15.61bcH1888.19±14.04abc30.94±12.24b41.43±12.70cF值5.94∗∗8.33∗∗4.94∗∗平均值89.0834.4862.22标准差9.066.218.29CV(%)10.1717.9929.41
2.4 马尾松种子表型性状相关性分析
马尾松单株优树种子各性状的相关性分析结果见表5,从表5可知,马尾松种子形态特征、品质指标和营养物质含量指标间存在一定的相关性。种子平均体积与种子弧长、弧宽均呈极显著正相关,相关系数为0.991和0.922,说明马尾松种子形态分布均匀;种子平均体积与可溶性蛋白质、可溶性糖含量呈显著正相关,与千粒重呈极显著正相关;种子弧宽与可溶性蛋白质、可溶性糖含量呈显著正相关,相关系数为0.384和0.31;种子各发芽指标间呈极显著正相关,与可溶性蛋白质、脂肪含量呈显著正相关;种子可溶性蛋白质与脂肪含量呈显著正相关,相关系数为0.318;种子千粒重与种子弧长、弧宽呈极显著正相关,相关系数分别为0.922和0.991,与可溶性蛋白质、可溶性糖含量呈显著正相关;但脂肪含量与形态指标、千粒重及可溶性糖含量之间呈不显著的负相关。
3 结论与讨论
种子是植物的繁殖器官。植物在漫长的进化过程中为了适应复杂的环境会产生不同程度的遗传变异,并稳定地将遗传变异性状反映在种子的品质中[1]。大量的研究表明,林木种内存在不同层次的变异规律,主要表现在种源间、林分间和单株间,并且有些性状上的变异在种源间和家系间也不尽相同[12-15]。种子萌发能力的大小能够反映种子播种品质的优劣,还关系到苗木早期的生长状况[16],有研究者认为,可以通过将种子发芽能力作为参考来选择优良的种源[7,17]。本研究发现,不同马尾松优树单株在种子形态特征、萌发能力和营养物质含量上存在显著差异,种子的大小还与种子内营养物质含量存在显著相关性,不同优树单株种子相比,粒大的种子,可溶性蛋白质含量与脂肪含量相对较高;但总体上脂肪含量占马尾松种子内营养物质的主要部分。马尾松种子内营养物质含量的研究结果表明,脂肪含量高的种子,可溶性蛋白质含量相对也高,并且显著影响种子的发芽率、发芽势和发芽指数,说明马尾松种子萌发时主要的能量来源是脂肪和可溶性蛋白质,这与王培蒂等[18]研究结果一致。
本研究结果表明,在单株水平上,马尾松种子形态特征、萌发能力和营养物质含量存在一定的选择潜力。因此,在种子园的母树选择时,可结合种子表型性状进行综合考虑。而单株母树种子在苗期生长是否依然保持优良的遗传特性,还需要进一步的研究。
表5 马尾松单株优树种子形态特征、品质指标和营养物质的相关性分析

体积弧宽弧长发芽率发芽指数发芽势千粒重可溶性蛋白质可溶性糖弧宽0.99∗∗弧长0.92∗∗0.89∗∗发芽率0.040.04-0.08发芽指数-0.04-0.06-0.140.86∗∗发芽势-0.04-0.04-0.010.62∗∗0.79∗∗千粒重0.98∗∗0.99∗∗0.92∗∗0.04-0.040.04可溶性蛋白质0.37∗0.38∗0.250.43∗0.32∗0.46∗0.37∗可溶性糖0.39∗0.42∗0.16-0.01-0.07-0.250.43∗0.05脂肪-0.08-0.09-0.070.38∗0.41∗0.40∗-0.080.42∗-0.05
注:“*”在0.05水平(双侧)上显著相关;“**”在0.01水平(双侧)上显著相关。
[1]张世挺,杜国祯,陈家宽.种子大小变异的进化生态学研究现状与展望[J].生态学报,2003,23(2):353-364.
[2]王慧慧,王普昶,赵钢,等.干旱胁迫下白刺花种子大小与萌发对策[J].生态学报,2016,36(2):335-341.
[3]唐庆兰,黎海利,黄寿先,等.大叶栎优树种子性状变异研究[J].广西林业科学,2006,35(1):12-13.
[4]武冲,仲崇禄,张勇,等.麻棣种子形态和营养特征遗传变异分析[J].植物遗传资源学报,2014,15(2):429-435.
[5]刁松锋,邵文豪,姜景民,等.无患子实生群体种实表型性状变异研究[J].西北农林科技大学学报:自然科学版,2014(5):75-83.
[6]丁贵杰,周志春,王章荣,等.马尾松纸浆用材林培育与利用[M].北京:中国林业出版社,2006:35.
[7]秦晓佳,丁贵杰.不同地理种源马尾松种子性状及芽苗生长分析[J].种子,2012,31(1):14-17.
[8]李莹,刘淑欣,彭鸽,等.马尾松种子萌发与幼苗生长异养转自养生理过程研究[J].北京林业大学学报,2014,36(6):9-16.
[9]GB 2772—1999.林木种子检验规程[S].国家质量技术监督局,2000.
[10]李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
[11]尹燕枰,董学会.种子学实验技术[M].北京:中国农业出版社,2008:23-24.
[12]刘宇,徐焕文,尚福强,等.16年生白桦种源变异及区划[J].林业科学,2016,52(9):48-56.
[13]徐立安,陈天华.马尾松种源子代材性变异与制浆造纸材优良种源选择[J].南京林业大学学报(自然科学版),1997,21(2):1-6.
[14]胡勐鸿,贾子瑞,李青粉,等.云杉属7个种针叶性状种间和种内变异分析[J].西北植物学报,2013,33(12):2 529-2 536.
[15]郑书星,张建国,段爱国,等.新疆阿尔泰地区白杨派3个树种半同胞家系子代遗传多样性分析[J].林业科学研究,2013,26(3):366-372.
[16]李莹,刘淑欣,彭鸽,等.马尾松种子萌发与幼苗生长异养转自养生理过程研究[J].北京林业大学学报,2014,36(6):9-16.
[17]张兰,韦小丽,黎泽超,等.不同地理种源棕榈种子形态,萌发特性及内含物质比较[J].种子,2016,35(10):36-40.
[18]王培蒂,秦国峰.马尾松种子发育过程中种仁贮藏物质的积累与变化[J].林业科学研究,1996,9(2):146-151.
海菜花的花、叶均可观赏,是我国最适宜池塘、湖泊、河流绿化的水生栽培植物。繁殖是淡水大型沉水植物分布和丰富度格局变化的关键,繁殖成功是植物适应环境变化的综合体现。在淡水大型沉水植物的有性繁殖过程中,种子萌发是繁殖成功的关键过程之一[12]。
用赤霉素和吲哚丁酸(IBA)处理海菜花种子,可明显提高种子的发芽率、发芽势和发芽指数,缩短种子萌发的时间,比对照提前了2~4 d,随着浓度的增大而提高;浓度为75 mg/L时,发芽率、发芽势和发芽指数较高,对海菜花种子的萌发具有显著的促进作用,所以可以采用该浓度指导生产。
不同浓度水杨酸和α-萘乙酸处理对海菜花种子萌发均有抑制作用,浓度愈高,抑制作用愈强。相同浓度的α-萘乙酸比水杨酸的抑制作用强。
海菜花是一种较好的水生资源植物,但对环境要求较高,尤其是对水体污染较为敏感。海菜花生长快,特别是从第3年起生长尤快,寿命亦长,因此,海菜花可作为公园、学校、厂矿的池塘、河流、湖泊等水生绿化植物,然而目前尚未见海菜花育苗生产报道。本研究结果可为海菜花的综合开发利用提供技术指导和帮助。
参考文献:
[1]杨丽云,和加卫.飘然若仙海菜花[J].植物杂志,2000(5):5.
[2]蒋柱檀,李恒,刀志灵,等.云南传统食用植物海菜花的民族植物学研究[J].内蒙古师范大学学报(自然科学汉文版),2010,39(2):163-167.
[3]朱静,杨亚维,郭爱伟,等.云南几大湖泊海菜花营养成分分析[J].安徽农业科学,2010,38(24):1 292-1 295.
[4]梁士楚,李桂荣.靖西海菜花,谁来护卫[J].湿地科学与管理,2007,3(4):15.
[5]王苗,尹延震.海菜花种子的萌发实验研究[J].环境科学与管理,2014,39(11):112-115.
[6]王斌.波叶海菜花种子储存条件与萌发初步研究[J].种子,2011,30(9):5-7.
[7]赵素婷,厉恩华,杨娇,等.光照对海菜花(Otteliaacuminata)种子萌发、幼苗生长及生理的影响[J].湖泊科学,2014,26(1):107-112.
[8]王玄,袁思安,张建军,等.金莲花种子萌发特性研究[J].中国农学通报,2012,28(7):185-189.
[9]张志良,瞿伟菁,李小芳.植物生理学实验指导[M].北京:高等教育出版社,2009.
[10]丁秀英,张军,苏宝林.水杨酸在植物抗病中的作用[J].植物学通报,2001,18(2):163-168.
[11]Ratea DN,Cuencab JV,Bowmanb GR,et al.The Gain-of-function arabidopsis acd mutant reveals novel regula-tion and function of the salicylic acid signaling pathway incontrolling cell death,defenses and cell growth[J].Plant Cell,1999,11:1 695-1 708.
[12]Titus J E,Hoover D.Toward predisting reproductive success in subm ersed freshwater angiosperms[J].Aquatic Botany,1991,41:111-136.
Seed Phenotypic Variation Regularity Research from Plus Trees ofPinusmassoniana
WANGHaoyun1,2,ZHUXiaokun1,2,XIEWeibin3,WUFeng1,2
2017-01-26
国家自然科学基金(31660201)。
王好运(1990— ),男,河南濮阳人;硕士研究生,主要从事林木栽培生理生态与分子生物学研究;E-mail:475833435@qq.com。
吴 峰,副教授,硕士生导师,主要从事林木遗传育种研究;E-mail:fwu@gzu.edu.cn。
营养物质; 选择
10.16590/j.cnki.1001-4705.2017.08.069
S 791.248
A
1001-4705(2017)08-0069-04
