中国小麦农家品种‘老白麦’抗条锈性遗传分析
2017-11-29郭继元张建周王建超王吐虹王凤涛蔺瑞明张忠军徐世昌
郭继元, 张建周, 王建超, 王吐虹, 王凤涛, 冯 晶, 蔺瑞明*, 张忠军, 徐世昌*
(1. 中国农业科学院植物保护研究所, 植物病虫害生物学国家重点实验室, 北京 100193; 2. 河南省农业科学院小麦研究所,郑州 450002; 3. 中国农业大学植物保护学院, 北京 100193)
中国小麦农家品种‘老白麦’抗条锈性遗传分析
郭继元1,3, 张建周2, 王建超1, 王吐虹1, 王凤涛1, 冯 晶1, 蔺瑞明1*, 张忠军3, 徐世昌1*
(1. 中国农业科学院植物保护研究所, 植物病虫害生物学国家重点实验室, 北京 100193; 2. 河南省农业科学院小麦研究所,郑州 450002; 3. 中国农业大学植物保护学院, 北京 100193)
小麦条锈病是小麦生产上重要的气传叶部病害。不断发掘和利用新抗源是持续控制条锈病流行危害的重要基础研究工作。‘老白麦’是我国小麦农家品种,对我国当前主要流行小种和致病类型均表现为高抗水平。本研究采用常规杂交分析方法,对‘老白麦’及其与感病品种‘Taichung 29’的杂交后代在成株期和苗期分别接种CYR32号小种和CYR33号小种,进行抗条锈性鉴定和统计分析。结果表明,‘老白麦’对CYR33号小种在苗期和成株期均表现近免疫,其全生育期抗条锈性由1对显性基因控制;对CYR32号小种在成株期表现近免疫,苗期表现高感,成株抗条锈性由1对显性基因控制,属细胞核遗传。研究结果表明‘老白麦’至少含有2对显性抗病基因,分别控制‘老白麦’对CYR33号小种的全生育期抗性和对CYR32号小种的成株抗性。基因推导分析认为‘老白麦’对CYR33的全生育期抗性基因可能为未知新基因。建议在抗病育种中加以有效合理利用,促进小麦品种中抗病基因多样化布局。
条锈病; 抗病性; 遗传分析; 基因推导
小麦条锈病是由条形柄锈菌Pucciniastriiformisf.sp.triticiErikss.侵染引起的世界性真菌病害,发生频繁,危害严重[1]。中国是世界上小麦条锈病最大的流行区,每年都有不同程度的发生,造成严重经济损失[2]。自20世纪50年代以来,曾发生15次条锈病中度以上流行,其中1950、1964、1990、2002年4次全国大流行,使小麦分别减产60亿、32亿、26.5亿、13亿kg[3]。研究和实践表明,培育和种植抗锈良种是防治小麦条锈病最经济、安全又环境友好的方法。但由于抗源单一化利用和条锈菌高度变异性,新的致病类型和毒性小种的产生与发展,致使小麦品种丧失原有的抗病性。20世纪50年代至90年代,导致全国范围内7次大的抗病品种更替[4],之后CYR32和CYR33的出现与发展又使一大批生产品种相继丧失抗病性。因此,不断发掘利用新抗源是持续控制条锈病流行危害的重要基础研究工作。
小麦农家品种是人们长期生产实践积累下来的生产品种,虽然绝大多数被当前生产品种所代替,但因其含有丰富的抗病性和抗逆性基因资源备受育种家的重视,特别是作为条锈病的重要抗源[5-7]。目前国内外对农家品种抗条锈性遗传研究已有大量报道,研究发现,‘白老芒麦’对Su-11的抗性由1对显性和2对隐性互补基因控制[8],‘红麦’(‘京2747’)对CYR19的抗性由1对显性基因控制[9],‘赤壳’(‘苏1900’)对CYR32的抗性由1对显性基因控制[10],‘换香头’、‘白麦(1-3)’和‘白麦(2-1)’成株抗性均由1个隐性基因控制[11],‘老芒麦’对Su-11和CYR32的抗性均由2对显性基因控制[12],‘红锁条’对CYR31和CYR32抗病性均由2对隐性独立或重叠基因控制[13]。‘老白麦’是我国小麦农家品种,具有良好的成株期抗条锈性,但其抗病性遗传基础尚不清楚。明确其抗条锈病基因组成及遗传和抗性特点,可促进抗源的有效合理利用,进一步提高抗病基因多样性,为小麦条锈病的持续控制提供抗病基因资源。
1 材料与方法
1.1 材料
1.1.1 供试品种
‘老白麦’是我国通过长期系选而成的小麦农家品种,来源于甘肃甘谷,具有较好的抗条锈性;感病品种‘Taichung 29’,至今尚未发现有抗条锈病基因。采用常规杂交方法,以‘Taichung29’为母本或父本分别与抗病品种‘老白麦’杂交和自交,分别获正交和反交的F1、F2代材料(群体)。
1.1.2 供试菌系
CYR32和CYR33的单孢菌系,为中国小麦条锈菌当前主要流行小种,在感病品种‘铭贤169’上繁殖,并经中国小麦条锈菌鉴别寄主鉴定后备用。基因推导所用菌系来自国内外25个毒性谱不同的条锈菌系,即66E2、3E6、66E0、64E0、96E0、3E48、0E32、32E0、67E16、1E32、114E210、1E28、8E132、2E180、98E16、3E108、2E66、70E2、2E80、2E64、2E0、42E0、3E128、3E0、34E0。以上材料均由中国农业科学院植物保护研究所麦类真菌病害研究组提供。
1.2 试验方法
1.2.1 苗期抗病性鉴定
在中国农业科学院植物保护研究所人工气候室内(温度昼14~18℃/夜10~12℃,光周期L∥D=14 h∥10 h,光照强度6 000 lx,RH75%)进行。将‘Taichung 29’和‘老白麦’及其杂交后代播于直径9 cm的花盆中,双亲播种30粒,F1代各播种20粒,F2代群体播种300粒,每盆播种10粒。出苗后待第一片叶完全展开时用扫抹法分别接种CYR32和CYR33,在10℃的黑暗接种间内保湿24 h,然后转移到人工气候室中潜育发病,接菌后待‘Taichung 29’充分发病,调查记载侵染型。侵染型采用“0-4”型常规六级标准[1]。
1.2.2 成株期抗病性鉴定
于2011年-2013年在中国农业科学院植物保护研究所廊坊基地CYR32鉴定病圃进行。采用等行距条播方式,行距0.3 m,行长2.0 m,每行播种20粒,双亲各播种1行,F1代播种2行,正反交F2代群体均播种300粒。每畦两端分别播种2行‘铭贤169’作为保护行,试验小区周围播种1行‘铭贤169’作诱发行。选用条锈菌流行小种CYR32于4月上旬在田间喷雾接种感病品种‘铭贤169’作诱发行,在5月下旬当感病对照品种‘铭贤169’发病严重度达到80%以上时,分别调查记载供试材料的旗叶和倒二叶的侵染型。侵染型采用“0-4”型常规六级标准[1]。
1.2.3 基因推导分析
1973年Browder[14]依据F1or的基因对基因理论[15]提出了初步的抗病基因推导方法。Dubin[16]进一步完善了该方法,并指出根据寄主和病原相互作用产生的高低反应型和系谱分析相结合进行基因推导,同时提出六条基本原则。本研究利用来自国内外25个毒性谱不同条锈菌单孢菌系,以30个小麦抗条锈病单基因系或已知基因载体品系作为标准品系,对供试品种进行抗性鉴定,比较分析供试品种与标准品系相对应的侵染型组合,分析供试品种的抗性谱及其所携带的抗条锈基因(组合)。
2 结果与分析
2.1 苗期和成株期抗条锈性评价
‘老白麦’在苗期和成株期分别接种条锈菌单孢菌系CYR32和CYR33进行抗性鉴定。结果显示,‘老白麦’对CYR33在苗期和成株期均表现近免疫,侵染型为0;型;对CYR32在成株期表现近免疫,侵染型为0型,而苗期表现高感,侵染型为4型。表明 ‘老白麦’对CYR33具有全生育期抗性,对CYR32具有成株抗性。感病亲本‘Taichung 29’对CYR32和CYR33在苗期和成株期均表现高感,侵染型为4型。
2.2 苗期和成株期抗条锈性遗传分析
2.2.1 对CYR32的成株期抗条锈性遗传分析
由表1结果看出,‘老白麦’表现抗病,侵染型为0型,‘Taichung29’表现感病,侵染型为4型,F1代表现抗病,侵染型0型,F2代群体发生了抗感分离。根据双亲及杂交后代侵染型级别及各级侵染型的数目,将0~2型划为抗病类型,3~4型划为感病类型。在其265株的正交F2代群体中,抗病株和感病株分别为204株和61株,经χ2检验,符合由1对显性基因控制的理论比例(χ2{3∶1}=0.554 7,0.30≤P≤0.50);在其273株反交的F2代群体中,抗病株和感病株分别为210株和63株,经χ2检验,也符合由1对显性基因控制的理论比例(χ2{3∶1}=0.538 5,0.30≤P≤0.50)。表明‘老白麦’对CYR32的成株抗病性由1对显性基因控制,属细胞核遗传。
表1‘老白麦’及杂交后代在成株期对CYR32的抗性遗传分析
Table1Geneticanalysisofwheatlandrace‘Laobaimai’andtheiroffspringpopulationsatadultstagetoCYR32ofPucciniastriiformisf.sp.tritici
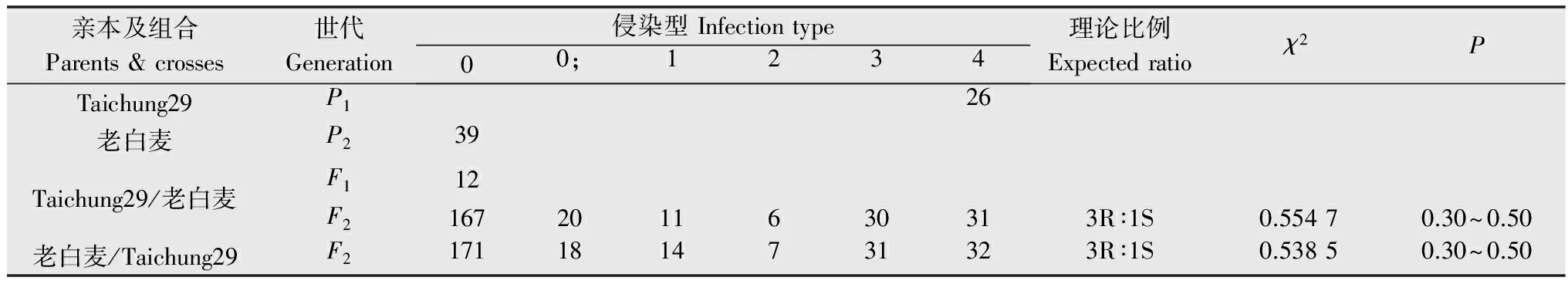
亲本及组合Parents&crosses世代Generation侵染型Infectiontype00;1234理论比例Expectedratioχ2PTaichung29P126老白麦P239Taichung29/老白麦F112F21672011630313R∶1S0.55470.30~0.50老白麦/Taichung29F21711814731323R∶1S0.53850.30~0.50
2.2.2 对CYR33的抗条锈性遗传分析
由表2结果看出,抗病亲本‘老白麦’对CYR33表现近免疫,侵染型为0;型,‘Taichung 29’表现高感,侵染型为4型,F1代表现抗病,侵染型为0;型,F2代群体发生了抗感分离。根据双亲及杂交后代侵染型级别及各级侵染型的数目,将0~2型划为抗病类型,3~4型划为感病类型。在其282株F2代群体中,抗病株和感病株分别为220株和62株,经χ2检验,符合由1对显性基因控制的理论比例(χ2{3∶1}=1.366 4,0.20≤P≤0.30)。表明‘老白麦’对CYR33的全生育期抗性由1对显性基因控制。
表2‘老白麦’及杂交后代在苗期对条锈菌系CYR33的遗传分析
Table2Geneticanalysisofwheatlandrace‘Laobaimai’andtheiroffspringpopulationsatseedlingstagetoCYR33ofPucciniastriiformisf.sp.tritici

亲本及组合Parents&crosses世代Generation侵染型Infectiontype00;1234理论比例Expectedratioχ2PTaichung29P119老白麦P226Taichung29/老白麦F112F2521575615473R∶1S1.36640.20~0.30
2.3 ‘老白麦’苗期抗条锈性基因推导分析
在温室利用25个条锈菌系对‘老白麦’和已知基因标准品系进行苗期抗条锈性鉴定。结果显示,感病对照品种‘铭贤169’对25个条锈菌系均高度感病,侵染型为4型;30个已知基因品系和供试品种‘老白麦’对25个条锈菌系的相互作用的侵染型组合及其与标准品系比对分析结果见表3。抗性谱比对分析结果显示,‘老白麦’对25个条锈菌系的抗性谱均与30个已知基因品系的抗性谱不同。表明‘老白麦’含有与供试的已知基因品系不同的抗条锈病基因。
表330个已知基因品系和供试品种接种25个条锈菌系的侵染型结果1)
Table3Infectiontypesof30lineswithknownresistancegenesandtestedcultivarsinoculatedwith25pathotypesofPucciniastriiformisf.sp.tritici
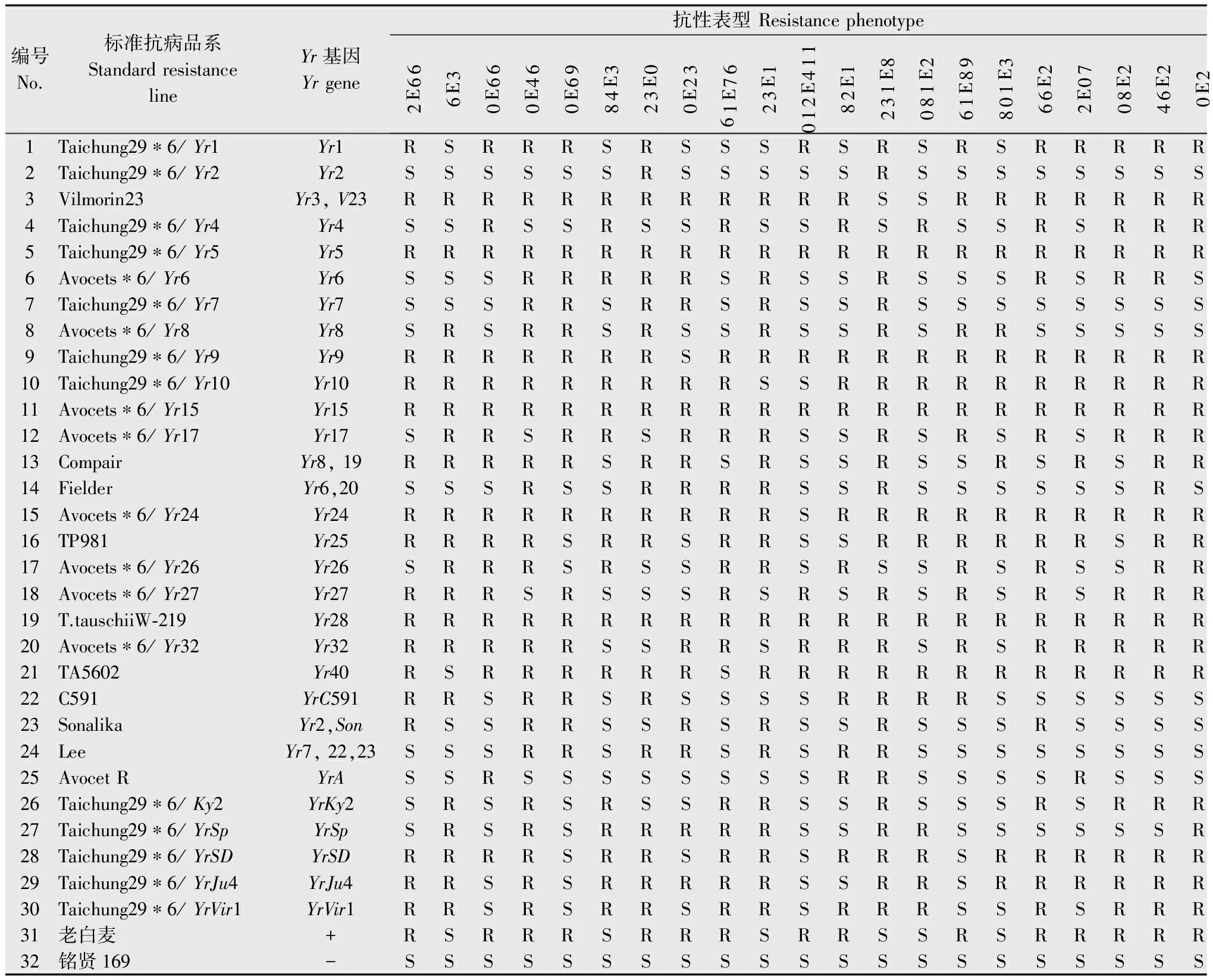
编号No.标准抗病品系StandardresistancelineYr基因Yrgene抗性表型Resistancephenotype66E23E666E064E096E03E480E3232E067E161E32114E2101E288E1322E18098E163E1082E6670E22E802E642E042E03E1283E034E01Taichung29∗6/Yr1Yr1RSRRRSRSSSRSRSRSRRRRRRSSR2Taichung29∗6/Yr2Yr2SSSSSSRSSSSSRSSSSSSSSSSSS3Vilmorin23Yr3,V23RRRRRRRRRRRRSSRRRRRRRSRRR4Taichung29∗6/Yr4Yr4SSRSSRSSRSSRSRSSRSRRRRSRR5Taichung29∗6/Yr5Yr5RRRRRRRRRRRRRRRRRRRRRRRRR6Avocets∗6/Yr6Yr6SSSRRRRRSRSSRSSSRSRRSSRSR7Taichung29∗6/Yr7Yr7SSSRRSRRSRSSRSSSSSSSSSRSS8Avocets∗6/Yr8Yr8SRSRRSRSSRSSRSRRSSSSSSSSS9Taichung29∗6/Yr9Yr9RRRRRRRSRRRRRRRRRRRRRRRSR10Taichung29∗6/Yr10Yr10RRRRRRRRRSSRRRRRRRRRRRRRR11Avocets∗6/Yr15Yr15RRRRRRRRRRRRRRRRRRRRRRRRR12Avocets∗6/Yr17Yr17SRRSRRSRRRSSRSRSRSRRRRRRR13CompairYr8,19RRRRRSRRSRSSRSSRSRSRRRRRR14FielderYr6,20SSSRSSRRRRSSRSSSSSSRSSRSS15Avocets∗6/Yr24Yr24RRRRRRRRRRSRRRRRRRRRRRRRR16TP981Yr25RRRRSRRSRRSSRRRRRRSRRRRRR17Avocets∗6/Yr26Yr26SRRRSRSSRRSRSSRSRSSRRRRRR18Avocets∗6/Yr27Yr27RRRSRSSSRSRSRSRSRSRRRRRSR19T.tauschiiW⁃219Yr28RRRRRRRRRRRRRRRRRRRRRRRRR20Avocets∗6/Yr32Yr32RRRRRSSRRSRRRSRSRRRRRRRRR21TA5602Yr40RSRRRRRRSRRRRRRRRRRRRRRRR22C591YrC591RRSRRSRSSSSRRRRSSSSSSSRSS23SonalikaYr2,SonRSSRRSSRSRSSRSSSRSSSSSSSS24LeeYr7,22,23SSSRRSRRSRSRRSSSSSSSSSSSS25AvocetRYrASSRSSSSSSSSRRSSSSRSSSRSSS26Taichung29∗6/Ky2YrKy2SRSRSRSSRRSSRSSSRSRRRSSRS27Taichung29∗6/YrSpYrSpSRSRSRRRRRSSRRSSSSSSRRRRS28Taichung29∗6/YrSDYrSDRRRRSRRSRRSRRRSRRRRRRSRRS29Taichung29∗6/YrJu4YrJu4RRSRSRRRRRSSRRSRRRRRRRRRS30Taichung29∗6/YrVir1YrVir1RRSRSRRSRRSRRRSSRSRRRRRRS31老白麦+RSRRRSRRRSRRSSRSRRRRRRSSR32铭贤169-SSSSSSSSSSSSSSSSSSSSSSSSS
1) R: 抗病; S: 感病;+: 未知基因。
R: Resistant; S: Susceptible;+: Unknown gene.
3 讨论
研究结果表明,‘老白麦’对CYR33在苗期和成株期均表现近免疫,其全生育期抗条锈性由1对显性基因控制;对CYR32在成株期表现近免疫,而在苗期则表现高感,其成株抗条锈性由1对显性基因控制,这意味着‘老白麦’至少含有对CYR33全生育期抗性和对CYR32成株抗性的2对显性抗条锈病基因,分别控制‘老白麦’在苗期和成株期对CYR33和CYR32的抗性。基因推导分析显示,控制对CYR33全生育期抗性的‘老白麦’抗性谱与供试的已知基因载体品系的抗性谱不同,表明‘老白麦’含有与已知基因载体品系所含的抗条锈病基因不同;另据系谱分析,‘老白麦’是通过系选而成的农家品种,同时尚未发现有证据表明供试的已知基因载体品系与‘老白麦’有亲缘关系,故认为对CYR33的全生育期抗性基因为未知新基因。‘老白麦’含有成株抗条锈病基因和全生育期抗条锈病基因,可抗我国当前主要流行小种CYR32和CYR33,是可利用的重要抗源材料。建议在今后抗病育种中加以有效合理利用,促进抗病基因多样化布局。
[1] 李振歧, 曾士迈. 中国小麦锈病[M]. 北京: 中国农业出版社, 2002: 1-362.
[2] 陈万权, 康振生, 马占鸿, 等. 中国小麦条锈病综合治理理论与实践[J]. 中国农业科学, 2013(20):4254-4262.
[3] 陈刚. 小麦条锈病区域流行的相关性分析[D].北京:中国农业大学, 2005.
[4] 万安民, 牛永春, 吴立人. 中国小麦品种抗条锈性丧失及其治理对策[C]∥中国国际农业科技年会,1999.
[5] 代君丽, 井金学, 李振岐, 等. 我国小麦农家品种抗条锈性评价及抗病品种的遗传分析[C]∥中国植物病理学会学术年会论文集. 杨凌:西北农林科技大学出版社,2007:144-149.
[6] 蔺瑞明, 郄彦敏, 冯晶, 等. 中国小麦农家品种抗条锈病的鉴定与评价[J]. 沈阳农业大学学报, 2010(5):535-539.
[7] 代君丽, 牛永春. 小麦农家品种大籽糙抗条锈性的遗传分析[J]. 遗传, 2003(3):311-313.
[8] 代君丽, 牛永春, 井金学, 等. 我国小麦农家品种白老芒麦的抗条锈性遗传分析[J]. 西北农业学报, 2003(1):121-122.
[9] 巫娟,牛永春,檀根甲.小麦农家品种红麦(京2747)主效抗条锈病基因的RAPD标记[J].植物遗传资源学报,2006(3):260-263.
[10] Liu Fanghui, Niu Yongchun, Deng Hui, et al. Mapping of a major stripe rust resistance gene in Chinese native wheat variety Chike using microsatellite markers [J]. Journal of Genetics and Genomics, 2007(12):123-130.
[11] 郄彦敏, 蔺瑞明, 冯晶, 等. 4个小麦农家品种对条锈菌成株抗性遗传分析[J]. 沈阳农业大学学报, 2010(6):673-676.
[12] 骆惠生, 曹世勤, 贾秋珍, 等. 农家品种老芒麦抗条锈基因遗传分析[J]. 甘肃农业科技, 2007(7):10-12.
[13] 宁利园, 郄彦敏, 王凤涛, 等. 中国小麦农家品种红锁条和白蚂蚱的抗条锈性遗传分析[J]. 植物保护学报, 2015(2):145-152.
[14] Browder L E.Probable genotype of someTriticumaestivum‘Agent’ derivatives for reaction toPucciniareconditaf.sp.tritici[J]. Crop Science, 1973, 13: 203-206.
[15] Flor H H.Current status of the gene-for-gene concept [J]. Annual Review of Phytopathology, 1971, 9: 275-296.
[16] Dubin H J, Johnson R, Stubbs R W. Postulated genes for resistance to stripe rust in selected CIMMYT and related wheats[J]. Plant Disease, 1989, 73(6): 472-475.
(责任编辑: 田 喆)
GeneticanalysisoftheresistanceofChinesewheatlandrace‘Laobaimai’tostriperust
Guo Jiyuan1,3, Zhang Jianzhou2, Wang Jianchao1, Wang Tuhong1, Wang Fengtao1, Feng Jing1, Lin Ruiming1, Zhang Zhongjun3, Xu Shichang1
(1.StateKeyLaboratoryforBiologyofPlantDiseasesandInsectPests,InstituteofPlantProtection,ChineseAcademyofAgriculturalSciences,Beijing100193,China; 2.WheatResearchInstitute,HenanAcademyofAgriculturalSciences,Zhengzhou450002,China; 3.CollegeofPlantProtection,ChinaAgriculturalUniversity,Beijing100193,China)
Stripe rust is one of the most destructive airborne leaf diseases of wheat. It is important to continuously develop and utilize new resources resistant to stripe rust, and often regarded as the critical basis for durably controlling the epidemic disease. Chinese wheat landrace ‘Laobaimai’ is highly resistant to most presently predominant races and pathotypes ofPucciniastriiformisf.sp.tritici. In this study, by using crossing and genetic analysis, resistant parent ‘Laobaimai’, susceptible ‘Taichung 29’ and their progeny populations derived from the crosses of these two cultivars were all inoculated with the two races CYR32 and CYR33 at seedling and adult stages respectively to identify their disease resistance levels, and statistical analysis was carried out. Based on the results, ‘Laobaimai’ was nearly immune to race CYR33 at both seedling and adult stages, containing a single dominant over-all resistant gene. On the other hand, ‘Laobaimai’ was highly susceptible to race CYR32 at seedling stage, while nearly immune to it at adult stage, indicating that its adult resistance was controlled by a single nuclear-genetic-type dominant gene. Accordingly, ‘Laobaimai’ contained at least two dominant resistance genes, over-all resistant gene for race CYR33 and adult resistance for race CYR32, respectively. Through gene postulation analysis, it was suggested that ‘Laobaimai’ possibly contained an unknown over-all resistance gene to race CYR33. We propose that the stripe rust resistance germplasm ‘Laobaimai’ could be reasonably used in wheat resistance breeding practice for promoting the diverse distribution of the resistance genes in wheat cultivars.
stripe rust; disease resistance; genetic analysis; gene postulation
2017-01-26
2017-02-28
国家自然科学基金(31261140370,31272033);国家重点基础研究发展计划(“973”计划)(2013CB127700);国家重点研发计划-粮丰工程(2016YFD0300705);农业部作物种质资源保护子项目(2017NWB036-12);中国农业科学院基本科研业务费专项(Y2017P706)
* 通信作者 E-mail: rmlin@ippcaas.cn;shichangxu317@163.com
S 435.121, S 432.21
A
10.3969/j.issn.0529-1542.2017.06.010
