农艺与气候因素对青藏高原栽培大麦淀粉含量空间分布的影响
2017-11-23钟志明王建林冯西博侯维海王改花大次卓嘎
钟志明,王建林,冯西博,付 刚,侯维海,王改花,大次卓嘎
1 中国科学院地理科学与资源研究所,北京 100101 2 西藏农牧学院, 林芝 860000
农艺与气候因素对青藏高原栽培大麦淀粉含量空间分布的影响
钟志明1,王建林2,*,冯西博2,付 刚1,侯维海2,王改花2,大次卓嘎2
1 中国科学院地理科学与资源研究所,北京 100101 2 西藏农牧学院, 林芝 860000
为了揭示不同环境因子对青藏高原栽培大麦籽粒淀粉含量(GSC)积累的影响程度,完善大麦GSC空间分异与环境因子的关系,明确青藏高原不同地区大麦品种GSC的环境效应,利用83个样点的地理、气候、土壤、农艺因子数据,研究了青藏高原栽培大麦GSC的分布特征,结果表明:(1)在地理水平方向上,青藏高原栽培大麦GSC的水平分布总体呈现出斑块状交错分布和南高北低的格局,并形成了以西藏拉孜、隆子、堆龙德庆、曲水、尼木、定日、萨迦、达孜、扎囊、日喀则、墨竹工卡、贡嘎、琼杰为中心的青藏高原中南部和以青海共和、贵德、海晏、同德为中心的青藏高原东北部等2个栽培大麦GSC高值区;(2)在地理垂直方向上,栽培大麦GSC的变化呈现出“S”型分布格局,即在海拔3300.0—3600.0 m以上,随着海拔的升高,栽培大麦GSC逐渐增加,在海拔4200.0 m与4500.0 m之间达到最高值,此后随着海拔的升高略有下降;(3)影响栽培大麦GSC的因子从大到小的顺序是穗密度﹥6月平均气温日较差>芒长﹥9月平均气温>1月平均气温>年日照时数﹥≥0℃积温>5月平均气温﹥8月平均气温日较差>8月平均气温>6月平均气温>≥10℃积温>6月平均月降水量>5月平均月降水量>7月平均相对湿度>8月平均相对湿度>7月平均气温。这一研究结果显示,对栽培大麦GSC影响最大的是基因型,其次是气候因素,土壤因素对GSC的影响不明显。影响栽培大麦GSC的农艺因子主要是穗密度和芒长,气候因子主要是拔节抽穗期的气温日较差和籽粒灌浆成熟期的平均气温,日照和降水的影响相对较小。
农艺因素;气候因素;大麦;淀粉含量;空间分布;青藏高原
大麦是全球栽培的第四大谷类作物,其适应性强,利用价值广泛,不仅广泛用于食品和饲料,还用于啤酒酿造。淀粉是大麦中存储的一种重要多糖,同时也是全球重要的食物和工业原料。淀粉是便宜的、可再生,能进行生物降解[1]。淀粉有直链淀粉和支链淀粉两类,前者为无分支的螺旋结构,后者以24—30个葡萄糖残基以α- 1,4-糖苷键首尾相连而成,在支链处为α- 1,6-糖苷键[2- 4]。不论是食用、饲用还是酿造啤酒,大麦籽粒淀粉含量(Grain Starch Content,GSC)均是一个十分重要的品质指标[5]。近年来,一些研究者认为大麦GSC不仅明显受其本身遗传因素的影响,还与栽培条件及产地的环境条件密切相关[6- 14]。但是,不同的环境因子是如何影响大麦GSC的,目前尚存在很大争议[15-22]。而且,这些研究多是在几个地点进行试验所得出的结论。截至目前,国内外尚未见到在大尺度上进行栽培大麦GSC与环境因子关系的研究报道。
青藏高原既是世界栽培大麦起源中心之一,又是中国重要的裸大麦(青稞)种植区。这里蕴藏着许多高GSC的种质资源[5,23-24],急需进一步挖掘与鉴别利用。迄今为止,除个别学者测定分析过部分西藏栽培大麦GSC外,尚未见到有关青藏高原栽培大麦GSC与环境因子关系的系统性研究报道。为此,本文试图以青藏高原为研究区,从大尺度上研究栽培大麦GSC的空间分异规律,以期为揭示不同环境(地理、气候、土壤)因子对青藏高原栽培大麦GSC积累的影响程度,完善大麦GSC空间分异与环境因子的关系,明确青藏高原不同地区大麦品种GSC的环境效应,为指导青藏高原大麦育种、栽培和加工提供理论依据。
1 材料和方法
1.1 研究区概况
研究区涉及四川、甘肃、青海、西藏等4个省(区)主要栽培大麦种植区。研究区跨越山地草原、高寒草甸草原、高寒灌丛草甸、山地针叶林、山地灌丛草原、山地荒漠、半荒漠等7个自然植被带,年均日照时数1642—3393 h,年均温-0.6—12.9℃,年均降水量150.0—890.0 mm。研究对象为当地种植的栽培大麦(均为裸大麦)。
1.2 样品采集与分析
(1)样点布置:2014年6—9月,根据青藏高原大气温度和降水分布格局,于不同生态种植区(27°—38°N、79°—104°E)设置83个典型样点。在样点设置时,重点选择长势整齐、成片种植的栽培大麦田,所有样点的具体地理位置如图1所示。

图1 采样点分布图Fig.1 The locations of sampling sites
(2)植物样品采集与分析:采样在栽培大麦成熟期进行。采样前先用GPS定位、咨询当地农民了解样点所在农田种植的栽培大麦品种、施肥情况等基本信息。采样时在每个样点设5—6个样方,于每个样方(面积不小于20 m2)中随机采收栽培大麦籽粒2000 g,送回实验室风干后,用酶水解法(GB5009.9- 85)测定栽培大麦的GSC。同时,在每个样点,随机选择20个植株进行常规大田考种。
(3)土壤样品采集与分析:在每个样点进行植物采样的同时,设5—6个样方,于每个样方(面积不小于20.0 m2)中随机采集耕作层(0—30.0 cm)土壤样品3份,将其装入土壤布袋,送回实验室风干后,磨细过1 mm筛,测定土壤全氮、全磷、全钾、速效氮、速效磷、速效钾、pH值、有机质等理化指标。其中:土壤全氮和速效氮采用半微量凯氏法,土壤全磷和速效磷测定采用钼锑抗比色法,土壤全钾和速效钾测定采用原子吸收分光光度法,土壤有机质采用重铬酸钾氧化—外加热法、土壤pH值采用电位法进行测定。
1.3 数据分析与处理
首先将83个样点栽培大麦GSC数据分成5类(即<40,40—45,45—50,50—60和>60,单位:%),然后利用ARCGIS 9.3软件作其空间分布图。同时,采用DPS软件进行统计分析。运用单因素方差分析和最小显著差异法进行数据组间的差异性比较,用逐步回归分析法建立栽培大麦GSC与地理、气候(注:气候资料来源于中国国家气象信息中心)、土壤、农艺因子之间的关系方程,并用随机森林回归分析法综合分析不同因子对青藏高原栽培大麦GSC的影响程度。
2 结果与分析
2.1 青藏高原栽培大麦GSC的分布特征
2.1.1 青藏高原栽培大麦GSC的水平分布特征
从图2可以看出,栽培大麦GSC在青藏高原形成了两个高值区。其中:一个是介于87.0°—92.0°E、29.0°—30.0°N之间,以西藏拉孜、隆子、堆龙德庆、曲水、尼木、定日、萨迦、达孜、扎囊、日喀则、墨竹工卡、贡嘎、琼杰为中心的青藏高原中南部高值区,这一区域栽培大麦GSC平均在(62.1635±5.6130)%左右;另一个是介于100.0°—101.5°E、35.0°—36.5°N之间,以青海共和、贵德、海晏、同德为中心的青藏高原东北部高值区,这一区域栽培大麦GSC平均在(57.9762±2.0447)%左右。同时,从图2还可以看出,青藏高原栽培大麦GSC的水平分布总体呈现出斑块状交错分布和南高北低的趋势。

图2 青藏高原栽培大麦GSC水平分布状况Fig.2 GSC distribution conditions along the horizontal direction in Qinghai-Tibet Plateau
2.1.2 青藏高原栽培大麦籽粒淀粉的垂直分布特征
从表1可以看出,青藏高原栽培大麦GSC的平均值为(48.1675±9.9556)%,变异系数为19.69%。随着海拔升高,栽培大麦GSC的变化呈现出“S”型分布格局。即在海拔3300.0—3600.0 m以下, 栽培大麦GSC随着海拔的变化略有波动性变化,但是总体含量很低,平均值为(45.9538±9.2987)%。在海拔3300.0—3600.0 m以上,随着海拔的升高,栽培大麦GSC逐渐增加,在海拔4200.0—4500.0 m之间达到最高值,此海拔区间栽培大麦籽粒淀粉平均含量为(50.4275±10.7836)%,此后随着海拔的升高栽培大麦GSC略有下降。其中在3900.0—4500.0 m之间,栽培大麦GSC平均在(50.1431±10.7156)%左右。同时,从表1还可以看出,栽培大麦GSC变异系数随着海拔的升高而呈现有波动性变化。
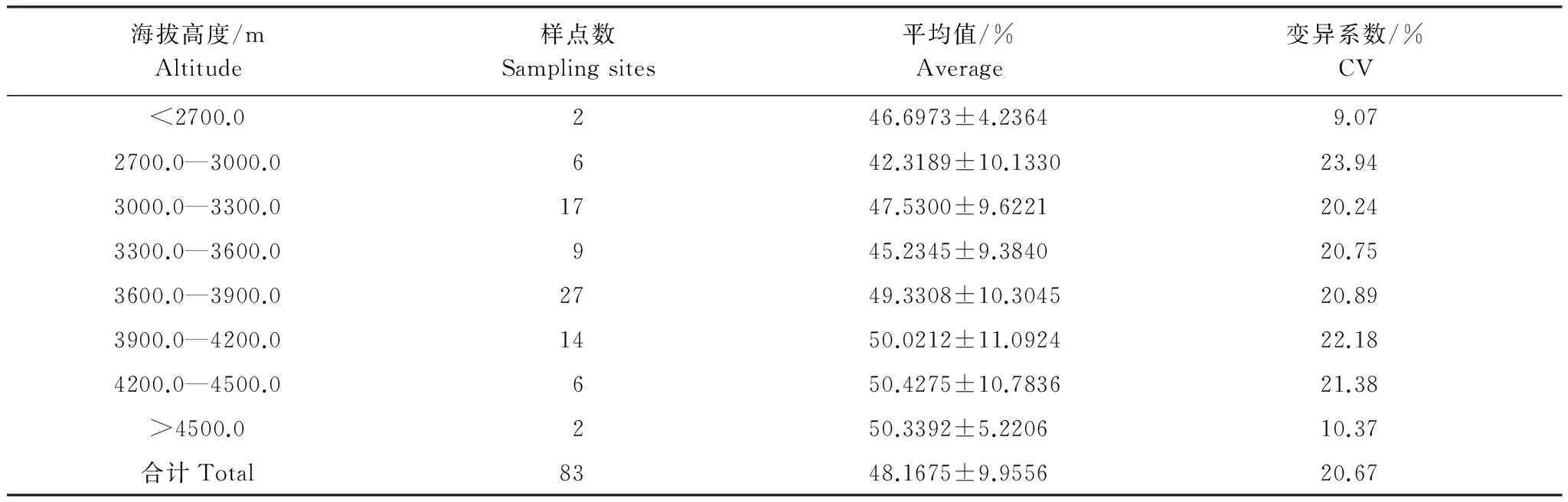
表1 不同海拔梯度栽培大麦GSC的分布状况
2.2 青藏高原栽培大麦GSC与生态因子的关系
2.2.1 栽培大麦GSC与地理因子的关系
基于逐步回归分析法,建立栽培大麦GSC与地理纬度(X1)、地理经度(X2)、海拔(X3)等地理因子的关系方程如下:
Y=26.7002+0.1311X1-0.1752X2+0.0040X3
(N=83R=0.1646R1=0.0517R2=-0.0327R3=0.1264) (1)
从式(1)可以看出,栽培大麦GSC(Y)与地理经度(X1)、海拔高度(X3)呈正相关关系,而与地理纬度(X2)呈负相关关系。根据标准误差检验,该回归方程未通过=0.05水平的显著性检验。表明,栽培大麦GSC随着地理经度和海拔高度的增加以及地理纬度的降低而增加。同时也表明,地理因子对栽培大麦GSC的影响未达到显著性水平(P<0.05)。
2.2.2 栽培大麦GSC与气候因子的关系
基于逐步回归分析法,建立栽培大麦GSC与年均日照时数(X1)、年蒸发量(X2)、≥0℃积温(X3)、≥10℃积温(X4)、年均气温(X5)、年均相对湿度(X6)、年均降水量(X7)、1月平均气温(X8)、5月平均气温(X9)、6月平均气温(X10)、7月平均气温(X11)、8月平均气温(X12)、9月平均气温(X13)、5月平均气温日较差(X14)、6月平均气温日较差(X15)、7月平均气温日较差(X16)、8月平均气温日较差(X17)、9月平均气温日较差(X18)、5月平均相对湿度(X19)、6月平均相对湿度(X20)、7月平均相对湿度(X21)、8月平均相对湿度(X22)、9月平均相对湿度(X23)、5月平均降水量(X24)、6月平均月降水量(X25)、7月平均降水量(X26)、8月平均降水量(X27)、9月平均降水量(X28) 等气候因子的关系方程如下:
Y=68.4096+0.0066X1-7.3785X3-3.0721X4+1.2481X6+12.2109X8-8.0992X9+12.6286X10-10.3583X11+
9.8997X12-9.5364X13-3.1067X15+3.2766X17-1.8666X21+0.3827X22-0.5219X24+0.8191X25
(N=83R= 0.5968*R1=0.3449**R3=-0.3538**R4=-0.3576**R6=0.1716R8=0.3728**R9= -0.2635*R10=0.3262**R11=-0.4007**R12=0.293*R13=-0.2426*R15=-0.3747**R17=0.3898**R21=-0.374**R22=0.3652**R24=-0.245*R25=0.2890*)
(2)
从式(2)可以看出,影响栽培大麦GSC(Y)的主要气候因子是年均日照时数(X1)、≥0℃积温(X3)、≥10℃积温(X4)、年均相对湿度(X6)、1月平均气温(X8)、5月平均气温(X9)、6月平均气温(X10)、7月平均气温(X11)、8月平均气温(X12)、9月平均气温(X13)、6月平均气温日较差(X15)、8月平均气温日较差(X17)、7月平均相对湿度(X21)、8月平均相对湿度(X22)、5月平均降水量(X24)、6月平均月降水量(X25),而年蒸发量(X2)、年均气温(X5)、年均降水量(X7)、5月平均气温日较差(X14)、7月平均气温日较差(X16)、9月平均气温日较差(X18)、5月平均相对湿度(X19)、6月平均相对湿度(X20)、9月平均相对湿度(X23)、7月平均降水量(X26)、8月平均降水量(X27)、9月平均降水量(X28)的影响则很小。
同时,从式(2)还可以看出,栽培大麦GSC与年均日照时数(X1)、1月平均气温(X8)、6月平均气温(X10)、8月平均气温(X12)、8月平均气温日较差(X17)、8月平均相对湿度(X22)、6月平均月降水量(X25)的偏相关系数达到显著或极显著正相关差异水平,与≥0℃积温(X3)、≥10℃积温(X4)、5月平均气温(X9)、7月平均气温(X11)、9月平均气温(X13)、6月平均气温日较差(X15)、7月平均相对湿度(X21)、5月平均降水量(X24)的偏相关系数达到显著或极显著负相关差异水平,但是与年均相对湿度(X6)的偏相关关系未达到显著性(P<0.05)差异。
2.2.3 栽培大麦GSC与土壤因子的关系
基于逐步回归分析法,建立栽培大麦GSC与土壤有机质(X1)、全氮(X2)、全磷(X3)、全钾(X4)、速效氮(X5)、速效磷(X6)、速效钾含量(X7)和土壤pH值(X8)等土壤因子的数学模型:
Y=55.6769-7.1357X4-0.0260X7(N=83R=0.2523R4=-0.145R7=-0.1929)
(3)
从式(3)可以看出,影响栽培大麦GSC(Y)的主要土壤因子是土壤全钾(X4)、速效钾含量(X7),而土壤有机质(X1)、全氮(X2)、全磷(X3)、速效氮(X5)、速效磷(X6)、和土壤pH值(X8)的影响则很小。同时从式(3)还可以看出,栽培大麦GSC与土壤全钾(X4)、速效钾含量(X7)呈负相关关系。根据标准误差检验,该回归方程未通过=0.05水平的显著性检验。表明,栽培大麦GSC随着土壤全钾(X4)、速效钾含量(X7)的减少而增加。同时也表明,土壤因子对栽培大麦GSC的整体影响未达到显著性水平(P<0.05)。
2.2.4 栽培大麦GSC与农艺因子的关系
基于逐步回归分析法,建立栽培大麦GSC与株高(X1)、穗长(X2)、穗密度(X3)、穗形(X4)、穗姿(X5)、总小穗数(X6)、不孕小穗数(X7)、穗粒数(X8)、芒长(X9)、籽粒颜色(X10)、籽粒形状(X11)、护颖宽度(X12)、千粒重(X13)等农艺因子的数学模型:
Y=1.7731+1.8331X3-3.8122X5+0.1279X6+1.0718X9-0.5077X10+0.1916X13
(N=83R= 0.4717**R3= -0.3207**R5= -0.1885R6=0.1803R9= 0.3683R10= -0.1219R13=0.1683**)
(4)
从式(4)可以看出,影响栽培大麦GSC(Y)的主要农艺因子是穗密度(X3)、穗姿(X5)、总小穗数(X6)、芒长(X9)、籽粒颜色(X10)、千粒重(X13),而株高(X1)、穗长(X2)、穗形(X4)、不孕小穗数(X7)、穗粒数(X8)、籽粒形状(X11)、护颖宽度(X12)的影响则很小。同时从式(4)还可以看出,栽培大麦GSC与芒长(X9)的偏相关系数达到极显著正相关差异水平,与穗密度(X3)的偏相关系数达到极显著负相关差异水平,但是与穗姿(X5)、总小穗数(X6)、千粒重(X13)、籽粒颜色(X10)的偏相关关系均未达到显著性(P<0.05)差异。根据标准误差检验,该回归方程通过了=0.01水平的显著性检验。表明,栽培大麦GSC随着芒长(X9)的增加和穗密度的减少而显著增加。同时也表明,农艺因子对栽培大麦GSC的整体影响达到极显著性水平(P<0.01)。
2.2.5 栽培大麦GSC与综合因子的关系
通过以上分析可以看出,栽培大麦GSC与年均日照时数、1月平均气温、6月平均气温、8月平均气温、8月平均气温日较差、8月平均相对湿度、6月平均月降水量、芒长的偏相关系数达到显著或极显著正相关差异水平,与≥0℃积温、≥10℃积温、5月平均气温、7月平均气温、9月平均气温、6月平均气温日较差、7月平均相对湿度、5月平均降水量、穗密度的偏相关系数达到显著或极显著负相关差异水平。但是,这些因子对栽培大麦GSC的影响程度如何尚不清楚。为此,我们运用随机森林回归分析法,对栽培大麦GSC影响显著的年均日照时数、1月平均气温、6月平均气温、8月平均气温、8月平均气温日较差、8月平均相对湿度、6月平均月降水量、≥0℃积温、≥10℃积温、5月平均气温、7月平均气温、9月平均气温、6月平均气温日较差、7月平均相对湿度、5月平均降水量、穗密度、芒长等17个变量进行了随机森林回归分析。从表2可以看出,这17个变量对栽培大麦GSC影响从大到小的顺序是穗密度﹥6月平均气温日较差>芒长﹥9月平均气温>1月平均气温>年日照时数﹥≥0℃积温>5月平均气温﹥8月平均气温日较差>8月平均气温>6月平均气温>≥10℃积温>6月平均月降水量>5月平均月降水量>7月平均相对湿度>8月平均相对湿度>7月平均气温。

表2 不同因子对栽培大麦GSC影响的重要程度
3 结论与讨论
在地理水平方向上,青藏高原栽培大麦GSC的水平分布总体呈现出斑块状交错分布和南高北低的格局,并形成了以西藏拉孜、隆子、堆龙德庆、曲水、尼木、定日、萨迦、达孜、扎囊、日喀则、墨竹工卡、贡嘎、琼杰为中心的青藏高原中南部和以青海共和、贵德、海晏、同德为中心的青藏高原东北部等2个栽培大麦GSC高值区;在地理垂直方向上栽培大麦GSC的变化呈现出“S”型分布格局,即在海拔3300.0—3600.0 m以下, 栽培大麦GSC虽有波动性变化,但是总体含量很低,在海拔3300.0—3600.0 m以上,随着海拔的升高,栽培大麦GSC逐渐增加,在海拔4200.0—4500.0 m达到最高值,此后随着海拔的升高栽培大麦GSC略有下降。表明,今后要选择高β淀粉的青藏高原栽培大麦品种,可能从这些区域的地方品种中进行选择可能会有更大的收获。要发展青藏高原青稞淀粉产业,可能在这些地区进行规划种植栽培大麦可能会有更好的收效。
农艺、气候、土壤等因子对栽培大麦GSC影响从大到小的顺序是穗密度﹥6月平均气温日较差>芒长﹥9月平均气温>1月平均气温>年日照时数﹥≥0℃积温>5月平均气温﹥8月平均气温日较差>8月平均气温>6月平均气温>≥10℃积温>6月平均月降水量>5月平均月降水量>7月平均相对湿度>8月平均相对湿度>7月平均气温。表明,对栽培大麦GSC影响最大的是基因型,其次是是气候因素,是二者共同作用的结果,土壤因素对GSC的影响不明显。影响栽培大麦GSC的农艺因子主要是穗密度和芒长,气候因子主要是拔节抽穗期的气温日较差和籽粒灌浆成熟期的平均气温,日照和降水的影响相对较小。这一研究结果支持大麦籽粒淀粉含量虽然主要受品种特性,即遗传因素的控制,但环境因素对它也有明显影响的研究结果[5- 14]。
栽培大麦GSC与年均日照时数、6月平均月降水量的偏相关系数均达到显著正相关差异水平,与9月份平均气温的偏相关系数均达到显著负相关差异水平,与土壤含氮量的偏相关系数未达到显著性差异水平。这与Erbs等对德国大麦的研究表明,随着灌浆期温度的升高和降水的减少,作物淀粉含量下降[8];Ahmed等对西藏大麦的研究表明,干旱条件下,大麦淀粉含量下降[9];Macleod和Duffus认为大麦籽粒发育中,英国大麦淀粉含量随着温度的升高而减少[25];Wei等对中国大麦的研究表明,随着温度的升高,淀粉含量减少[26]; Reinhardt等对德国大麦的研究研究表明,随着温度的升高,作物淀粉含量下降[27]的结论一致,但是与Holopainen等对英国二棱大麦(Hordeumvulgare)的研究表明,随着日照时间的缩短,大麦淀粉含量增多,且二者差异未达到显著水平[28];Erbs等对德国大麦的研究表明,随着施氮量的增加,淀粉含量显著下降[28]的结论相反。
[1] Goldstein A, Annor G, Putaux J L, Hebelstrup K H, Blennow A, Bertoft E. Impact of full range of amylose contents on the architecture of starch granules. International Journal of Biological Macromolecules, 2016, 89: 305- 318.
[2] Pérez S, Bertoft E. The molecular structures of starch components and Their contribution to the architecture of starch granules: A comprehensive review. Starch, 2010, 62(8): 389- 420.
[3] Wang S J, Copeland L. Effect of acid hydrolysis on starch structure and functionality: a review. Critical Reviews in Food Science and Nutrition, 2015, 55(8): 1081- 1097.
[4] Hermansson A M, Svegmark K S. Developments in the understanding of starch functionality. Trends in Food Science & Technology, 1996, 7(11): 345- 353.
[5] 王建林. 西藏高原作物栽培学. 北京: 中国农业出版社, 2012.
[6] Griffey C, Brooks W, Kurantz M, Thomason W, Taylor F, Obert D, Moreau R, Flores R, Sohn M, Hicks K. Grain composition of Virginia winter barley and implications for use in feed, food, and biofuels production. Journal of Cereal Science, 2010, 51(1): 41- 49.
[7] Fox G, Borgognone G M, Flinn P, Poulsen D. Genetic and environmental analysis of NIR 3. Feed quality predictions on genotypes of barley (HordeumvulgareL.). Field Crops Research, 2011, 120(3): 380- 386.
[8] Erbs M, Manderscheid R, Jansen G, Seddig S, Pacholski A, Weigel H J. Effects of free-air CO2enrichment and nitrogen supply on grain quality parameters and elemental composition of wheat and barley grown in a crop rotation. Agriculture, Ecosystems & Environment, 2010, 136(1/2): 59- 68.
[9] Ahmed I M, Cao F B, Han Y, Nadira U A, Zhang G P, Wu F B. Differential changes in grain ultrastructure, amylase, protein and amino acid profiles between Tibetan wild and cultivated barleys under drought and salinity alone and combined stress. Food Chemistry, 2013, 141(3): 2743- 2750.
[10] Novo P, Vojislava M I, Jasmina S, Milan M. Effect of growing season and variety on quality of spring two-rowed barley. Genetika, 2014, 46(1): 59- 73.
[11] Ahmed Z, Tetlow I J, Ahmed R, Morell M K, Emes M J. Protein interactions among enzymes of starch biosynthesis in high-amylose barley genotypes reveal differential roles of heteromeric enzyme complexes in the synthesis of A and B granules. Plant Science, 2015, 233: 95- 106.
[12] Ahmadi A, Baker D A. The effect of water stress on the activities of key regulatory enzymes of the sucrose to starch pathway in wheat. Plant Growth Regulation, 2001, 35(1): 81- 91.
[13] Ahmed I M, Dai H X, Zheng H X, Cao F B, Zhang G P, Sun D F, Wu F B. Genotypic differences in physiological characteristics in the tolerance to drought and salinity combined stress between Tibetan wild and cultivated barley. Plant Physiology and Biochemistry, 2013, 63: 49- 60.
[14] Balla K, Rakszegi M, LI Z Y, Békés F, Bencze S, Veisz O. Quality of winter wheat in relation to heat and drought shock after Anthesis. Czech Journal of Food Science, 2011, 29(2): 117- 128.
[15] Chen Z, Newman I, Zhou M, Mendham N, Zhang G, Shabala S. Screening plants for salt tolerance by measuring K+ux: a case study for barley. Plant, Cell & Environment, 2005, 28(10): 1230- 1246.
[16] Brandt C, Balko C, Eichler-Löbermann B. Interactive effects of soil water content and phytin supply on phosphorus nutrition of different crops species. Landbauforschung, 2011, 61(4): 317- 325.
[17] Clausen S K, Frenck G, Linden L G, Mikkelsen T N, Lunde C, Jørgensen R B. Effects of single and multifactor treatments with elevated temperature, CO2and ozone on oilseed rape and barley. Journal of Agronomy and Crop Science, 2011, 197(6): 442- 453.
[18] Ferris R, Ellis R H, Wheeler T R, Hadley P. Effect of high temperature stress at anthesis on grain yield and biomass of field-grown crops of wheat. Annals of Botany, 1998, 82(5): 631- 639.
[19] Gusmao M, Siddique K H M, Flower K, Nesbitt H, Veneklaas E J. Water deficit during the reproductive period of grass pea (LathyrussativusL.) reduced grain yield but maintained seed size. Journal of Agronomy Crop Science, 2012, 198(6): 430- 441.
[20] del Moral G L F, Rharrabti Y R, Martos V, Royo C. Environmentally induced changes in amino acid composition in the grain of durum wheat grown under different water and temperature regimes in a Mediterranean environment. Journal of Agricultural and Food Chemistry, 2007, 55(20): 8144- 8151.
[21] Goupy P, Hugues M B, Boivin P, Amiot M J. Antioxidant composition and activity of barley (Hordeumvulgare) and malt extracts and of isolated phenolic compounds. Journal of the Science of Food and Agriculture, 1999, 79(12): 1625- 1634.
[22] Hejgaard J, Boisen S. High-lysine proteins in Hiproly barley breeding: Identification, nutritional significance and new screening methods. Hereditas, 2009, 93(2): 311- 320.
[23] 马得泉. 中国西藏大麦遗传资源. 北京: 中国农业出版社, 2000.
[24] 王建林, 栾运芳, 大次卓嘎, 胡单. 西藏栽培大麦变种组成和分布规律研究. 中国农业科学, 2006, 39(11): 2163- 2169.
[25] Macleod L C, Duffus C M. Temperature effects on starch granules in developing barley grains. Journal of Cereal Science, 1988, 8(1): 29- 37.
[26] Wei K, Dai F, Wu F B, Zhang G P. The Variation of β-amylase activity and protein fractions in barley grains as affected by genotypes and post-Anthesis temperatures. Journal of the Institute of Brewing, 2009, 115(3): 207- 213.
[27] Reinhardt D, Jansen G, Seddig S, Eichler-Loebermann B. Temperature stress during flowering time affects yield and quality parameters of waxy barley. Landbauforschung Volkenrode, 2013, 63(1): 79- 83.
[28] Holopainen U R M, Wilhelmson A, Home S, Poutanen K, Shewry P R. Day-length effects on protein localisation affect water absorption in barley (Hordeumvulgare) grains. Food and Agriculture, 2012, 92(15): 2944- 2951.
EffectsofagronomicandclimaticfactorsonthespatialdistributionofstarchconcentrationinbarleycultivatedontheQinghai-TibetPlateau
ZHONG Zhiming1, WANG Jianlin2, *, FENG Xibo2, FU Gang1, HOU Weihai2, WANG Gaihua2, Dacizhuoga2
1InstituteofGeographicSciencesandNaturalResourcesResearch,ChineseAcademyofSciences,Beijing100101,China2CollegeofPlantScience,TibetInstituteofAgricultureandAnimalHusbandry,Linzhi860000,China
Starch concentration is an important quality character of barley grain. In recent years, some researchers have realized that barley grain starch concentration (GSC) is affected by genetic factors, as well as by its cultivation and environmental conditions. However, the effect of different environmental factors on barley GSC remains unclear. In addition, previous studies have mostly been conducted at several sites, and large-scale studies have not addressed the relationship between cultivated barley GSC and environment factors. The Qinghai-Tibet Plateau is an important region for growing hull-less barley in China, and the relationship between barley GSC and environment factors on the Qinghai-Tibet Plateau have not been reported. Therefore, the present study investigated the distribution of barley GSC on the Qinghai-Tibet Plateau using geography, climate, soil, and agronomy data from 83 sampling sites, the cultivated barley GSC distribution characteristics in Qinghai-Tibet Plateau was studied in this research work. The aim of the study was to identify the relative effects of different environmental factors on GSC, to clarify the relationship between the distribution of barley GSC and environmental factors, and to get explicit GSC acclimation of barley varieties planted in different areas to environment. The results showed that (1) GSC exhibited a macrocosm pattern of spatial distribution along the horizontal direction on the Qinghai-Tibet Plateau, with staggered patches of different value and a tendency of “high in the south and low in the north.” In addition, two regions of higher GSC (south-central and north-eastern) included the Lazi, Longzi, Duilongdeiqin, Qushui, Nimu, Dingri, Sajia, Dazi, Zhanang, Shigatse, Mozhugongka, Gongga, and Qiongjie Counties in Tibet and the Gonghe, Guide, Haiyan, and Tongde Counties in the Qinghai Province. (2) Along the vertical direction, GSC exhibited an S-shaped distribution pattern. Indeed, GSC increased gradually with increasing altitude from 3300.0—3600.0 m, reached its maximum between 4200.0 m and 4500.0 m, and thereafter slightly decreased with increasing altitude. (3) The relative effects of the environmental factors on barley GSC were as follows: spike density > average diurnal temperature range in June > length of the awn > average temperature in September > average temperature in January > annual sunshine hours > accumulated temperature (≥0 ℃) > average temperature in May > average diurnal temperature range in August > average temperature in August > average temperature in June > accumulated temperature (≥10 ℃) > average precipitation in June > average precipitation in May > average relative humidity in July > average relative humidity in August > average temperature in July. Therefore, genotype had the greatest impact on the GSC of cultivated barley, followed by climatic factors, and the effects of soil factors effect were not apparent. Spike density and length of the awn were the most important agronomic factors, and diurnal temperature range during the period of elongating and heading and average temperature during the period of filling and maturation were the main climatic factors, whereas the effects of sunshine hours and precipitation were relatively small. These findings support previous reports that barley GSC is mainly controlled by the characteristics (i.e., genetic factors) of individual barley varieties, although the effects of environmental factors were also apparent.
agronomic factor; climatic factor; barley; starch concentration; spatial distribution; Qinghai-Tibet Plateau
国家自然科学基金 (31360300,31370458,31560362); 西藏自治区重点项目(XZXTCX- 2015-03,XBTSZWXK- 2015-01);西藏农牧学院雪域英才工程人才发展支持计划项目(2015XYB01);作物学科建设项目(2015ZWXKJS)
2016- 08- 31; < class="emphasis_bold">网络出版日期
日期:2017- 07- 11
*通讯作者Corresponding author.E-mail: xzwangjl@126.com
10.5846/stxb201608311782
钟志明,王建林,冯西博,付刚,侯维海,王改花,大次卓嘎.农艺与气候因素对青藏高原栽培大麦淀粉含量空间分布的影响.生态学报,2017,37(21):7197- 7205.
Zhong Z M, Wang J L, Feng X B, Fu G, Hou W H, Wang G H, Dacizhuoga.Effects of agronomic and climatic factors on the spatial distribution of starch concentration in barley cultivated on the Qinghai-Tibet Plateau.Acta Ecologica Sinica,2017,37(21):7197- 7205.
