近岸典型生态系统大型底栖动物的次级生产力
——以瓯江口、洞头列岛为例
2017-11-23贾胜华廖一波曾江宁高爱根汤雁滨
贾胜华,廖一波,曾江宁,*,寿 鹿,黄 伟,高爱根,汤雁滨
1 国家海洋局海洋生态系统与生物地球化学重点实验室, 杭州 310012 2 国家海洋局第二海洋研究所, 杭州 310012
近岸典型生态系统大型底栖动物的次级生产力
——以瓯江口、洞头列岛为例
贾胜华1,2,廖一波1,2,曾江宁1,2,*,寿 鹿1,2,黄 伟1,2,高爱根1,2,汤雁滨1,2
1 国家海洋局海洋生态系统与生物地球化学重点实验室, 杭州 310012 2 国家海洋局第二海洋研究所, 杭州 310012
为更好了解和估算河口及岛屿生态系统大型底栖动物次级生产力的情况,以近岸两个典型生态系统河口-瓯江口、岛屿-洞头列岛为例,于2015年4月和10月在该海域进行大型底栖动物调查,采用3种Brey经验模型对大型底栖动物次级生产力进行估算,并分析次级生产力与环境因子的关系。结果表明:(1)以去灰干重(ash-free dry weight, AFDW)计,Brey经验公式估算瓯江口和洞头列岛海域年均次级生产力分别为0.48 g(AFDW) m-2a-1和0.70 g(AFDW) m-2a-1,Brey模型估算瓯江口和洞头列岛海域年均次级生产力分别为0.51 g(AFDW) m-2a-1和1.55 g(AFDW) m-2a-1,Brey模型估算瓯江口和洞头列岛海域年均次级生产力分别为0.25 g(AFDW) m-2a-1和0.99 g(AFDW) m-2a-1。(2)3种经验模型反映大型底栖动物年均次级生产力空间分布趋势一致:瓯江口海域有1高值区,位于瓯江口南部;洞头列岛海域有2高值区,分别位于洞头本岛东北部的三盘岛与花岗岛之间和东黄岛东南部的岛屿之间。(3)瓯江口海域大型底栖动物年均次级生产力主要贡献种为双鳃内卷齿蚕Aglaophamusdibranchis、长吻沙蚕Glycerachirori、异蚓虫Heteromastusfiliforms、焦河篮蛤Potamocorbulaustulata、薄云母蛤Yoldiasimilis、圆筒原盒螺Eocylichnabraunsi,它们对次级生产力的贡献率超54.2%;洞头列岛海域大型底栖动物年均次级生产力主要贡献种为双鳃内卷齿蚕、异蚓虫、薄云母蛤、绒毛细足蟹Raphidopusciliatus、隆线强蟹Eucratecrenata、脑纽虫Cerebratulinasp.、红狼牙鰕虎鱼Odontamblyopusrubicundus,它们对次级生产力的贡献率超57.1%。(4)次级生产力与环境因子的相关性分析显示化学需氧量、悬浮物和表层沉积物中值粒径是影响瓯江口海域大型底栖动物年均次级生产力的重要环境因子,而环境因子与洞头列岛海域大型底栖动物年均次级生产力相关关系不显著。(5)瓯江口海域Brey经验公式与Brey模型估算结果基本一致,洞头列岛海域Brey经验公式与Brey模型估算结果基本一致。
次级生产力;大型底栖动物;洞头列岛;瓯江口;Brey经验模型
大型底栖动物次级生产力是生态系统物质循环和能量流动量化的基础,是表征生态系统功能的一个重要参数[1- 2]。估算次级生产力不仅可帮助人们了解大型底栖动物的功能变化,而且对研究生态系统的物流、能流过程和海洋生物资源的持续利用以及评价海洋环境压力也具有重要意义[3- 5]。河口与岛屿是近岸海域两个典型的生态系统,河口区位于海洋与陆地交界处,受入海河流携带的陆源沉积物、有机物以及无机营养物质影响,海洋环境具有高营养化、水体层化、环流的特点,历来是研究的热点区域[6];海岛处于海岸带的前沿,在发展沿海经济、推动国防建设、保护海洋环境等方面具有重要意义。目前,国内报道岛屿与河口大型底栖动物次级生产力的研究较少[7],并且鲜有比较岛屿与河口两个典型生态系统次级生产力的报道。而且国内大型底栖动物次级生产力估算基本采用Brey经验公式[7- 12],估算结果比较粗略,仅有胶州湾潮间带[13]和辽东湾西部海域[14]两篇报道采用Brey估算模型[15],而采用Brey估算模型[1]计算大型底栖动物次级生产力国内尚无报道。
国外学者提出许多大型底栖动物次级生产力估算模型[3- 4,15- 18],并对不同模型进行了比较[4,19],这些模型或是建模数据范围较小,不一定适用于低纬度海域[18];或是采用生物寿命等不易确定的参数,难以应用到实践[4,16]。目前,Brey提出的经验模型在国内外大型底栖动物群落次级生产力估算中应用较为广泛[5,8,10,12- 14]。国外次级生产力估算模型已由Brey经验公式[3]发展到Brey估算模型[15],再到最近的Brey估算模型[1]。Brey估算模型[1]考虑的参数多达20个,模型更为精细、复杂,虽其计算表单设计有输入层、隐藏层和结果层,但只需输入数据,结果则自动显示,因此操作非常方便。
研究海域位于27°41′—28°13′N,120°52′—121°16′E的浙江南部,包括洞头列岛海域、瓯江口海域,兼有岛屿、河口2个典型生态系统。目前关于该海域大型底栖动物次级生产力研究尚无报道,并且没有比较两个区域次级生产力的研究,本文拟采用Brey经验公式[3]、Brey估算模型[1]以及Brey估算模型[15]分别计算瓯江口海域及洞头列岛海域大型底栖动物次级生产力,比较两个研究区域的次级生产力大小和3种估算方法产生的结果差距,并分析其与环境因子的关系,以期为河口、岛屿两个典型生态系统大型底栖动物功能研究、生态环境保护提供理论基础。
1 材料与方法
1.1 站位分布与样品采集处理
大型底栖动物采样于2015年4月(春季)和10月(秋季)进行,共布设35个调查站位(图1)。其中,S8、S14—S17、S25—S30、S32—S34调查站位位于瓯江口海域;S1—S7、S9—S13、S18—S24、S31、S35调查站位位于洞头列岛海域。采用0.1 m2的Van Veen型抓斗式底质采样器取样,每站重复取样2次,合并为一个样品。使用孔径为0.5 mm的网筛分选样品,采用5%的福尔马林溶液固定所获生物样品,带回实验室进行种类鉴定。样品的采集、保存、计数、称量和资料整理均按照《海洋调查规范》(GB/T 12763.6——2007)[20]进行。
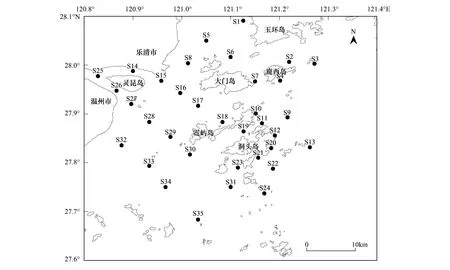
图1 洞头列岛及邻近海域大型底栖动物采样站位Fig.1 Sampling stations of macrozoobenthos in Dongtou Islands and adjacent sea area
1.2 数据处理
根据经验模型对数据的要求,需将生物量湿重数据转换为去灰干重生物量或能值。Brey经验公式[3]中生物量湿重(WW)的9/50为去灰干重(AFDW)[8]。Brey估算模型[1]和Brey估算模型[15]中,生物量湿重、去灰干重、能值的转换关系[13-15]见表1。
1.3 次级生产力估算经验模型
1.3.1Brey经验公式[3]
计算所有物种的年均去灰干重生物量和年均个体去灰干重,按照公式(1)逐种计算大型底栖动物年均次级生产力,将所有物种年均次级生产力相加得到该站位大型底栖动物群落年均次级生产力。
lgP=a+b1lgB+b2lgW
(1)
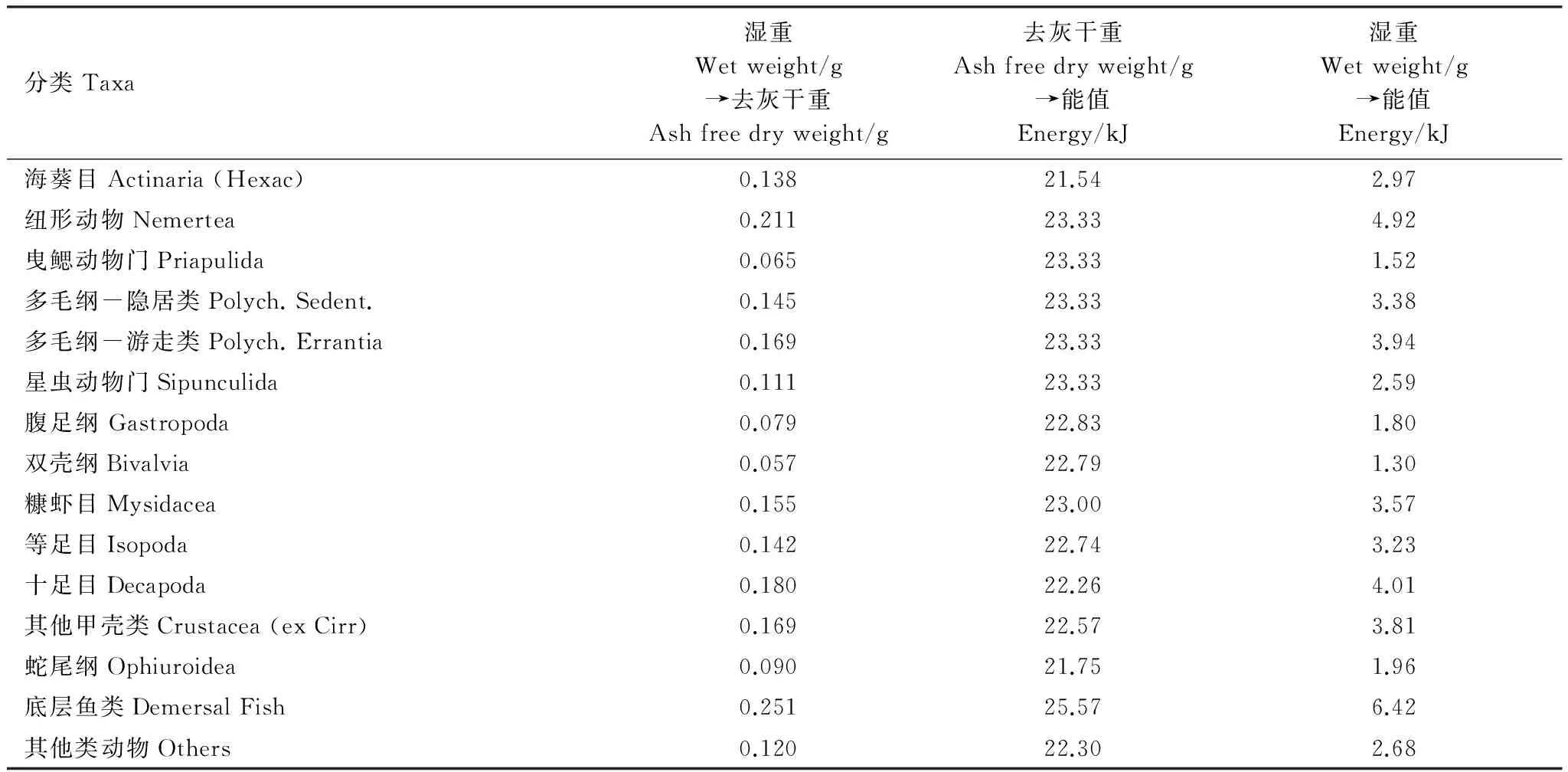
表1 大型底栖动物湿重生物量、去灰干重和能值的转换关系[13-15]
大型底栖动物不同类群的经验公式系数a,b1和b2见表2。其中:P为大型底栖动物年均次级生产力,单位为g(AFDW) m-2a-1;B为大型底栖动物年均去灰干重,单位为g(AFDW)/m2;W为大型底栖动物个体的平均去灰干重,单位为g(AFDW)/个。

表2 Brey经验公式中不同类群的3个系数值[3]
a:系数a,Coefficient a;b1:系数b1,Coefficient b1;b2:系数b2,Coefficient b2
1.3.2Brey估算模型[4- 5,15,22]
与Brey经验公式[3]相比,Brey估算模型[15]除考虑大型底栖动物的密度、生物量等生物因素,还考虑到水深、水温、生境等环境因素,见公式(2)。
lgP=7.947+lgB-2.294lgM-2409.856/(T+273)+0.168/D+0.194SubT+
0.180InEpi+0.277MoEpi+0.174Taxon1-0.188Taxon2+0.33Taxon3-0.062Habitat1+582.851lgM/(T+273)
(2)
其中,P为大型底栖动物年均次级生产力(kJ m-2a-1);B为年平均生物量对应的能值(kJ/m2);M为年平均个体体重能值(kJ);T为平均底层水温(℃);D为平均水深(m);SubT:潮下带值为1,潮间带为0;InEpi:底内生物值为1,底表生物为0;MoEpi:移动生物值为1,固着生物为0;Taxon1:环节动物或甲壳动物值为1,其他类群为0;Taxon2,棘皮动物值为1,其他类群为0;Taxon3:昆虫值为1,其他类群为0;Habitat1:湖泊生境值为1,其他生境值为0。
1.3.3Brey估算模型[1,23]
在Brey估算模型[15]多元线性回归估算模型基础上,Brey估算模型[1]增加大型底栖动物食性、移动性等参数提出多参数人工神经网络(Artificial Neural Network, ANN)估算模型,可对大型底栖动物次级生产力进行更好的预测[23],见公式(3)、(4)、(5)。由于模型参数较多,各参数对应系数值可参考2012-01版本计算表单[1]。模型计算表单具有3个层次:输入层、隐藏层和结果层,输入层包含平均个体体重能值、水深、水温、不同类群、移动性、食性、生境等20个参数,结果层为P/B的对数值。只需输入每个物种的参数,结果自动显示,操作比较简便。计算时先把20个待输入数据放入其他表格中,再拷贝到计算表单中。
log(P/B)=a0+a1H1+a2H2
(3)
H1=TanH(b0+b1logM+b2/T+b3logD+b4Mollusca+b5Annelida+b6Crustacea+b7Echinodermata+b8Insecta+b9Infauna+b10Sessile+b11Crawler+b12FacSwim+b13Herbivor+b14Omnivor+b15Carnivor+b16Lake+b17River+b18Marine+b19Subtidal+b20Exploited)
(4)
H2=TanH(c0+c1logM+c2/T+c3logD+c4Mollusca+c5Annelida+c6Crustacea+c7Echinodermata+c8Insecta+c9Infauna+c10Sessile+c11Crawler+c12FacSwim+c13Herbivor+c14Omnivor+c15Carnivor+c16Lake+c17River+c18Marine+c19Subtidal+c20Exploited)
(5)
其中,P为大型底栖动物年均次级生产力(kJ m-2a-1);B为年平均生物量对应的能值(kJ/m2);M为年平均个体体重能值(J);T为平均底层水温(℃);D为平均水深(m);Mollusca:软体动物值为1,其他为0;Annelida:环节动物值为1,其他为0;Crustacea:甲壳动物值为1,其他为0;Echinodermata:棘皮动物值为1,其他为0;Insecta:昆虫值为1,其他为0;Infauna:底内生物值为1,其他为0;Sessile:固着生物值为1,其他为0;Crawler:爬行动物值为1,其他为0;FacSwim:兼性游泳生物值为1,其他为0;Herbivor:草食动物动物值为1,其他为0;Omnivor:杂食动物值为1,其他为0;Carnivor:肉食动物值为1,其他为0;Lake:湖泊生境值为1,其他生境值为0;River:河流生境值为1,其他生境值为0;Marine:海洋生境值为1,其他生境值为0;Subtidal:潮下带值为1,其他生境值为0;Exploited指鱼类,通常设置为0。
本文将春、秋两季大型底栖动物栖息密度和生物量取平均值,然后分别采用3种经验模型逐种计算大型底栖动物年均次级生产力。Brey经验公式[3]运用Excel软件进行数据处理,Brey估算模型[1]和Brey估算模型[15]从网站(http://www.thomas-brey.de/science/virtualhandbook/)下载计算表单,按要求进行数据处理。为便于经验模型计算结果间比较,将Brey估算模型[1]和Brey估算模型[15]计算结果按照表1转换关系换算为去灰干重(g(AFDW) m-2a-1)。采用SPSS 16.0软件进行相关性分析。
2 结果
2.1 种类组成
瓯江口海域两航次共采集到大型底栖动物81种。其中,多毛类39种,占48.1%;甲壳动物23种,占28.4%;软体动物9种,占11.1%;棘皮动物4种,占4.9%;其他类6种(包括鱼类、螠虫动物、纽形动物、海葵),占7.4%。洞头列岛海域两航次共采集到大型底栖动物111种。其中,多毛类50种,占45.0%;甲壳动物30种,占27.0%;软体动物18种,占16.2%;棘皮动物5种,占4.5%;其他类8种(包括鱼类、星虫动物、纽形动物、曳鳃动物),占7.2%。可以看出,瓯江口海域与洞头列岛大型底栖动物种类组成占比例最多的均为多毛类、甲壳动物和软体动物。
2.2 不同海域大型底栖动物栖息密度、生物量及次级生产力
瓯江口海域大型底栖生物年均栖息密度为150.18 个/m2,年均生物量为2.06 g/m2。洞头列岛海域大型底栖生物年均栖息密度为214.29 个/m2,年均生物量为4.96 g/m2。以去灰干重(AFDW)计,Brey经验公式[3]估算瓯江口和洞头列岛海域年均次级生产力分别为0.48 g(AFDW) m-2a-1和0.70 g(AFDW) m-2a-1,Brey模型[15]估算瓯江口和洞头列岛海域年均次级生产力分别为0.51 g(AFDW) m-2a-1和1.55 g(AFDW) m-2a-1,Brey模型[1]估算瓯江口和洞头列岛海域年均次级生产力分别为0.25 g(AFDW) m-2a-1和0.99 g(AFDW) m-2a-1(表3)。Brey经验公式[3]与Brey估算模型[15]对瓯江口海域大型底栖动物次级生产力估算结果基本一致,Brey模型[1]估算结果比前两者的平均值低49.5%;Brey经验公式[3]与Brey估算模型[1]对洞头列岛海域大型底栖动物次级生产力估算结果基本一致,Brey模型[15]估算结果比前两者的平均值高45.5%。3种模型估算结果均表明,洞头列岛海域大型底栖动物年均次级生产力高于瓯江口海域。

表3 瓯江口海域和洞头列岛海域大型底栖动物年均次级生产力
AFDW:去灰干重,ash-free dry weight
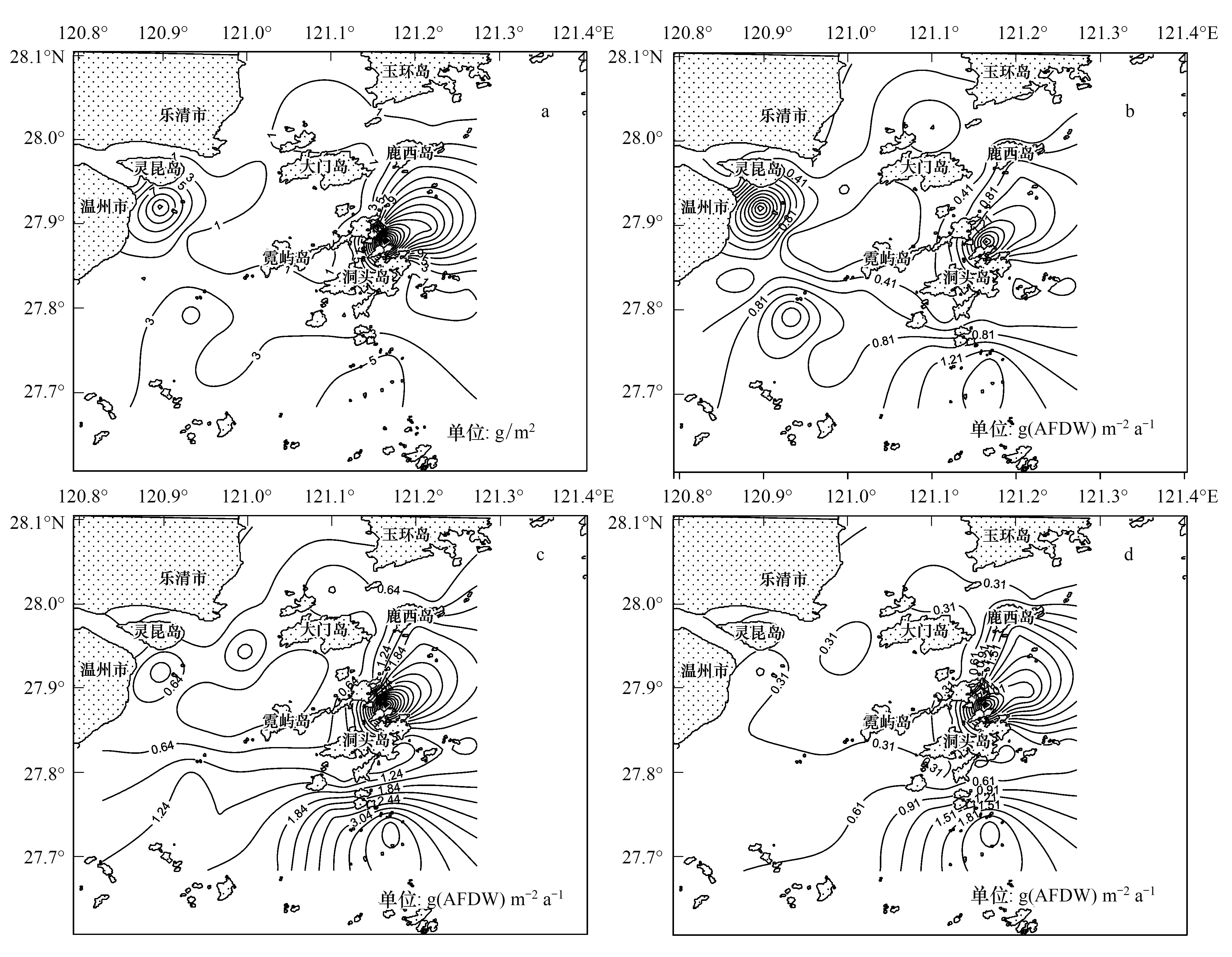
图2 大型底栖动物年均生物量(a)和Brey经验公式[3](b)、Brey估算模型[15](c)、Brey估算模型[1](d)年均次级生产力空间分布Fig.2 Distribution of the mean biomass (a) and productivity of macrobenthos estimated by Brey′s empirical formula[3] (b), Brey′s empirical model[15] (c), and Brey′s empirical model[1] (d)
3种经验模型反映大型底栖动物年均次级生产力空间分布趋势一致,如图2:瓯江口海域有1个高值区,出现在瓯江口南部海域;洞头列岛海域有2个高值区,分别出现在洞头本岛东北部的三盘岛与花岗岛之间海域和东黄岛东南部的岛屿之间海域,与大型底栖动物平均生物量分布规律相似。根据Brey经验公式[3]、Brey估算模型[15]和Brey估算模型[1]计算结果,年均次级生产力最低值均出现在瓯江口北部的S14站位,分别为0.01、0.04 g(AFDW) m-2a-1和0.01 g(AFDW) m-2a-1,原因是该站位大型底栖动物物种数少,生物量极低,仅有少量多毛类。Brey经验公式[3]计算结果表明,瓯江口南部海域S27站位大型底栖动物年均次级生产力最高,为2.62 g(AFDW) m-2a-1,原因是该站位出现了生物量和栖息密度均较大的焦河篮蛤Potamocorbulaustulata。该站位焦河篮蛤个体大,数量多,3种经验模型计算表明该种对该站位年均次级生产力贡献率均达到90%以上。Brey估算模型[15]和Brey估算模型[1]计算结果均表明S11站位大型底栖动物年均次级生产力最高,分别为6.25 g(AFDW) m-2a-1和4.97 g(AFDW) m-2a-1,原因是该站位大型底栖动物物种数多,达28种,栖息密度和生物量均较大。
2.4 大型底栖动物年均次级生产力的主要贡献物种
3种经验模型估算大型底栖动物年均次级生产力排前5位的物种基本一致(表4和表5),包括多毛类、软体动物、甲壳动物、鱼类和纽形动物。瓯江口海域大型底栖动物年均次级生产力主要贡献种为双鳃内卷齿蚕Aglaophamusdibranchis、长吻沙蚕Glycerachirori、异蚓虫Heteromastusfiliforms、焦河篮蛤P.ustulata、薄云母蛤Yoldiasimilis、圆筒原盒螺Eocylichnabraunsi,它们对该海域大型底栖动物年均次级生产力的贡献率超54.2%;洞头列岛海域大型底栖动物年均次级生产力主要贡献种为双鳃内卷齿蚕、异蚓虫、薄云母蛤、绒毛细足蟹Raphidopusciliatus、隆线强蟹Eucratecrenata、脑纽虫Cerebratulinasp.、红狼牙鰕虎鱼Odontamblyopusrubicundus,它们对该海域大型底栖动物年均次级生产力的贡献率超57.1%。
表43种经验模型估算瓯江口海域大型底栖动物年均次级生产力排前5位的物种
Table4Top5speciescontributetosecondaryproductivityofmacrobenthosinOujiangRiverestuaryestimatedbythreeempiricalmodels,respectively

经验模型Empiricalmodels次级生产力排前5的物种Thetop5speciesofsecondaryproductivity合计贡献率ThetotalcontributionrateBrey经验公式[3]Brey'sempiricalformula[3]焦河篮蛤、薄云母蛤、长吻沙蚕、圆筒原盒螺、异蚓虫82.9%Brey估算模型[15]Brey'sempiricalmodel[15]焦河篮蛤、长吻沙蚕、薄云母蛤、异蚓虫、双鳃内卷齿蚕58.1%Brey估算模型[1]Brey'sempiricalmodel[1]焦河篮蛤、长吻沙蚕、薄云母蛤、异蚓虫、双鳃内卷齿蚕54.2%
其中,双鳃内卷齿蚕、异蚓虫、薄云母蛤、绒毛细足蟹、圆筒原盒螺、长吻沙蚕属于分布范围较广、栖息密度和生物量均较高的物种,而隆线强蟹、焦河篮蛤属于个别站位栖息密度和生物量极高,在其他站位栖息密度和生物量偏低的物种。红狼牙鰕虎鱼、脑纽虫则属于出现频率和栖息密度较低,但个体大、生物量高的物种。
表53种经验模型估算洞头列岛海域大型底栖动物年均次级生产力排前5位的物种
Table5Top5speciescontributetosecondaryproductivityofmacrobenthosinDongtouIslandsestimatedbythreeempiricalmodels,respectively

经验模型Empiricalmodels次级生产力排前5的物种Thetop5speciesofsecondaryproductivity合计贡献率/%ThetotalcontributionrateBrey经验公式[3]Brey'sempiricalformula[3]薄云母蛤、异蚓虫、脑纽虫、红狼牙鰕虎鱼、绒毛细足蟹57.1Brey估算模型[15]Brey'sempiricalmodel[15]异蚓虫、脑纽虫、双鳃内卷齿蚕、绒毛细足蟹、红狼牙鰕虎鱼57.7Brey估算模型[1]Brey'sempiricalmodel[1]脑纽虫、红狼牙鰕虎鱼、绒毛细足蟹、异蚓虫、隆线强蟹59.2
2.5 大型底栖动物年均次级生产力与环境因子的关系
将3种经验模型估算大型底栖动物年均次级生产力与环境因子进行Pearson相关性分析。由表6和表7可知,3种经验模型估算年均次级生产力之间相关性极显著(P<0.01)。瓯江口海域Brey模型[15]估算结果与化学需氧量、悬浮物显著负相关(P<0.05),Brey模型[1]估算结果与悬浮物显著负相关(P<0.05)、与沉积物中值粒径显著正相关(P<0.05)。洞头列岛海域大型底栖动物年均次级生产力与环境因子相关关系不显著。
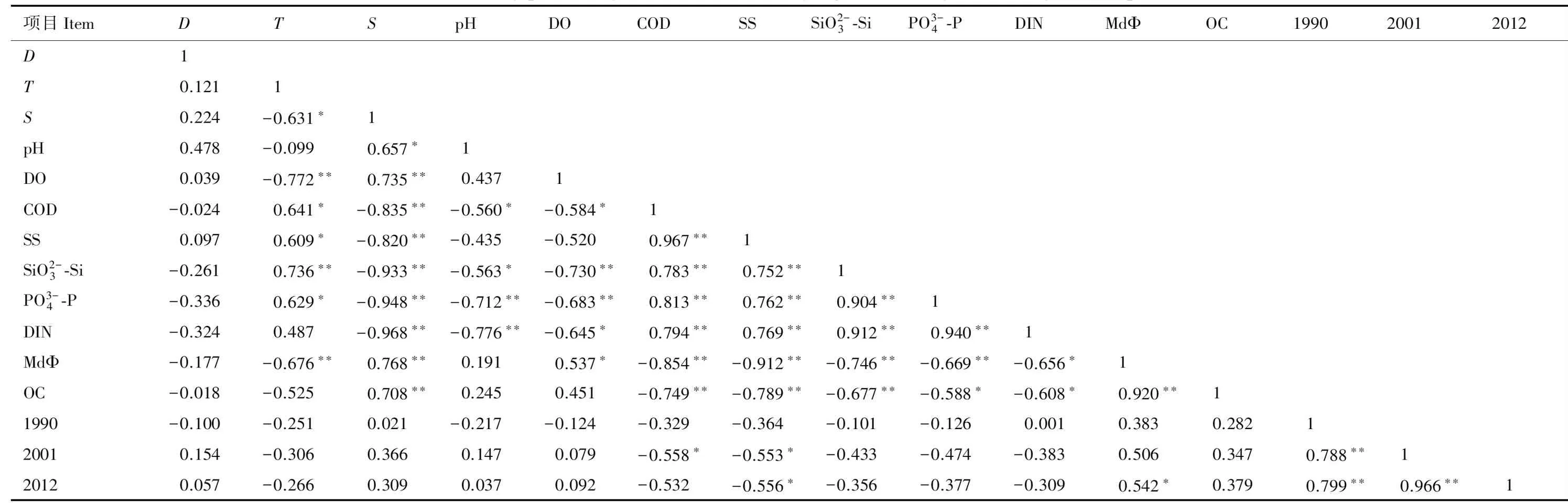
表6 3种经验模型估算瓯江口海域大型底栖动物年均次级生产力与环境因子的相关性
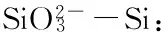

表7 3种经验模型估算洞头列岛海域大型底栖动物年均次级生产力与环境因子的相关性
3 讨论
3.1 次级生产力分布格局分析及与其他海域的比较
次级生产力的空间分布主要受大型底栖动物群落结构和栖息地环境条件影响[23]。瓯江口除南部以外的其他海域,年均次级生产力较低。河口海域由于受入海河流的影响,沉积速率较高,使底质处于强烈的扰动之中,在一定程度限制某些大型底栖动物的生存,如多毛类和部分甲壳类,活动能力强,对底质变化不敏感的物种才能生存,造成该区域大型底栖动物种类较少,因此次级生产力一般较低[7,14]。如辽东湾西部海域[14]受六股河影响,大型底栖动物的次级生产力有由近岸海域向远岸海域递增的趋势;受长江径流影响,河口区海底环境极不稳定,而距离河口较远的海域沉积环境趋于稳定,因此长江口[7]大型底栖动物次级生产力具有自长江入海口向东递增的趋势;越靠近深圳河河口[9],大型底栖动物的次级生产力越低。另外,近年来实施的温州浅滩围垦促淤工程在灵昆岛东侧到洞头霓屿岛西侧的滩涂上,建成南北两条围涂大堤,把灵昆岛与霓屿岛连成一片,该海域大部分为新淤出底质,大型底栖动物物种数少、生物量低。长江口滩涂围垦对大型底栖动物的影响研究[24]表明,围垦促淤可导致大型底栖动物物种减少,群落结构改变,生物量明显降低。因此,围垦促淤可能导致该海域次级生产力降低。另外,焦河篮蛤是咸淡水双壳类动物,为河口常见种,对环境适应能力较强。瓯江口南部海域年均次级生产力高的原因与厦门鼓浪屿附近海域[25]较为相似,鼓浪屿位于九龙江河口区,附近海域因出现大量光滑河篮蛤Potamocorbulalaevis导致次级生产力很高。
洞头本岛东北部的三盘岛与花岗岛之间海域和东黄岛东南部的岛屿之间海域因距离河口较远,沉积环境相对稳定,适宜于大型底栖动物生存。另外,岛屿生态系统有较高的生产力,海流流过岛屿时产生海底局部上升流使有机质均匀混合,为底栖动物提供丰富饵料,且该局部上升流或海流分支可为底栖动物提供多样性的栖息环境[6],岛屿之间的海域,大型底栖动物物种数、生物量一般较高[26]。因此,该海域大型底栖动物次级生产力较高。
与其他海域比较,瓯江口和洞头列岛海域大型底栖动物次级生产力处于较低水平,低于象山港港口海域[27]0.99 g(AFDW) m-2a-1、胶州湾海域[8]3.41 g(AFDW) m-2a-1、长江口及邻近海域[7]3.52 g(AFDW) m-2a-1及辽东湾西部海域[14]3.82 g(AFDW) m-2a-1的调查结果。历史调查资料[28]显示,洞头列岛海域属低生物量、低密度海域,该海域是浙江大型底栖动物年平均生物量最低的海域,栖息密度仅高于六横岛海域。河口海域由于底质不稳定可能次级生产力本身较低,如象山港口门附近站位[27]次级生产力仅为0.32 g(AFDW) m-2a-1。另外,瓯江口和洞头列岛海域次级生产力低可能与近年来围垦促淤有关,如研究表明[25]九龙江口海域底质受疏浚扰动,不利于大型底栖动物栖息,年次级生产力仅为0.31 g(AFDW) m-2a-1。而且相对依据全年资料估算的次级生产力数值,本文依据春秋两个季节资料估算的结果可能与实际次级生产力水平存在偏差。
3.2 次级生产力与环境因子的关系
环境因子如水深、水温、溶解氧等通过影响动物代谢速率和物种生活史,进而影响次级生产力[23],分析影响次级生产力的环境因子对全面理解海洋生态系统的功能具有重要意义[18]。由大型底栖动物次级生产力估算经验模型参数可知,除栖息密度和生物量外,次级生产力还与水深、底温密切相关[1,17-18],二者可分别影响食物质量和动物代谢速率[18]。许多研究表明[3- 4,8,10-11],大型底栖动物次级生产力随水深增加而降低。此外,影响大型底栖动物次级生产力的环境因子还有很多,如底质类型、盐度、溶解氧、沉积物中有机质含量、叶绿素a含量、总氮、总磷[10,12,18]等。袁伟等[10]对胶州湾西部海域大型底栖动物次级生产力与环境因子的相关性分析研究表明,叶绿素a是影响该海域次级生产力的重要因子。严娟等[12]采用典范对应分析长江口潮间带大型底栖动物群落次级生产力与环境变量的关系,发现水体总氮、总磷、盐度、沉积物有机质、溶解氧、叶绿素a均影响次级生产力,但不同断面影响因子不尽一致。
本研究发现化学需氧量、悬浮物、沉积物中值粒径是影响瓯江口海域大型底栖动物年均次级生产力的重要环境因子,环境因子与洞头列岛海域大型底栖动物年均次级生产力相关关系不显著。河口生态系统处于海洋与陆地交接处,陆源污染物可通过河口流入海洋。瓯江口海域可能由于近年来沿岸工业发展导致该海域有机污染物含量增加,有机污染通过微生物生化作用分解和氧化,大量消耗水中溶解氧,使水质变黑发臭[29],进而影响大型底栖动物的生存。且河口海域的悬浮物颗粒量可影响海水透明度和真光层厚度,从而影响浮游生物光合作用和初级生产力[30],进而影响次级生产力。通常认为沉积物粒度是影响大型底栖动物物种分布、栖息密度和生物量的重要环境因子[31],本研究发现瓯江口海域次级生产力与沉积物中值粒径显著正相关,证实了这一结论。岛屿间可能由于地理环境复杂,相关性分析结果表明环境因子与洞头列岛海域大型底栖动物次级生产力相关关系不显著。不同海域环境因素不同,且各个环境因子对底栖动物不同类群的作用也不同,要深刻理解影响大型底栖动物次级生产力的关键因素,需要对特定研究海域进行长期数据积累和对比分析,该领域有待进一步深入研究。
3.3 次级生产力估算经验模型的比较
次级生产力估算经验模型是对单个种群次级生产力研究的总结,极大地促进该领域研究的进步,使大型底栖动物群落次级生产力估算成为可能。由Brey经验公式[3]到Brey估算模型[15]再到Brey估算模型[1],模型越来越复杂、精细,表现在两方面:一是模型考虑的参数由2个增加到20个,并考虑环境因子;二是湿重生物量、去灰干重、能值转换由所有底栖动物相同转化比例精细到不同类群不同转换比例。另外,从建模方法看,Brey经验公式[3]和Brey估算模型[15]建模采用多元线性回归法,而Brey估算模型[1]采用人工神经网络法(Artificial Neural Network, ANN)。多元线性回归法对独立变量间的交互性非常敏感,而人工神经网络法可以模拟自变量和因变量间复杂的、非线性的、间断性的关系[1]。在预测大型底栖动物P/B值上,人工神经网络法优于多元线性回归法[32]。
经验模型只是对群落次级生产力的估算,并非真实值。3种经验模型所得次级生产力估算值存在一定差异,但在反映空间分布趋势和计算大型底栖动物次级生产力排前5的物种上大致相同。由于群落次级生产力的真实值较难测定,可将不同估算模型结果相吻合的数据认为是次级生产力的可靠估算值[13],本研究中Brey经验公式[3]与Brey估算模型[15]对瓯江口海域大型底栖动物次级生产力估算结果基本一致,Brey经验公式[3]与Brey估算模型[1]对洞头列岛海域大型底栖动物次级生产力估算结果基本一致,可认为Brey经验公式[3]和Brey估算模型[15]对瓯江口海域的估算结果比较可靠,Brey经验公式[3]和Brey估算模型[1]对洞头列岛海域的估算结果比较可靠。
致谢:感谢刘小涯、于培松、张海峰、蔡小霞、赵晨辉、陈文深等协助采样并提供相关理化参数。
[1] Brey T. A multi-parameter artificial neural network model to estimate macrobenthic invertebrate productivity and production. Limnology and Oceanography, 2012, 10(8): 581- 589.
[2] Dolbeth M, Cusson M, Sousa R, Pardal M A. Secondary production as a tool for better understanding of aquatic ecosystems. Canadian Journal of Fisheries and Aquatic Sciences, 2012, 69(7): 1230- 1253.
[3] Brey T. Estimating productivity of macrobenthic invertebrates from biomass and mean individual weight. Meeresforsch, 1990, 32(4): 329- 343.
[4] Cusson M, Bourget E. Global patterns of macroinvertebrate production in marine benthic habitats. Marine Ecology Progress Series, 2005, 297: 1- 14.
[5] Petracco M, Cardoso R S, Corbisier T N, Turra A. Secondary production of sandy beach macrofauna: an evaluation of predictive models. Estuarine, Coastal and Shelf Science, 2012, 115: 359- 365.
[6] 寿鹿, 曾江宁, 廖一波, 赵永强, 江志兵, 陈全震, 高爱根, 杨家新. 瓯江口海域大型底栖动物分布及其与环境的关系. 应用生态学报, 2009, 20(8): 1958- 1964.
[7] 刘勇, 线薇薇, 孙世春, 吴耀泉. 长江口及其邻近海域大型底栖动物生物量、丰度和次级生产力的初步研究. 中国海洋大学学报, 2008, 38(5): 749- 756.
[8] 李新正, 王洪法, 张宝琳. 胶州湾大型底栖动物次级生产力初探. 海洋与湖沼, 2005, 36(6): 527- 533.
[9] 金亮, 蔡立哲, 周细平, 徐华林. 深圳湾北岸泥滩大型底栖动物次级生产力研究. 台湾海峡, 2007, 26(3): 415- 421.
[10] 袁伟, 张志南, 于子山. 胶州湾西部海域大型底栖动物次级生产力初步研究. 应用生态学报, 2007, 18(1): 145- 150.
[11] 王宗兴, 孙丕喜, 刘彩霞, 张新军, 代欣欣, 周颖敏. 桑沟湾大型底栖动物的次级生产力. 应用与环境生物学报, 2011, 17(4): 495- 498.
[12] 严娟, 庄平, 侯俊利, 张涛, 刘鉴毅, 冯广朋, 杨刚. 长江口潮间带大型底栖动物次级生产力及其影响因子. 应用与环境生物学报, 2012, 18(6): 935- 942.
[13] 张崇良, 徐宾铎, 任一平, 薛莹, 纪毓鹏. 胶州湾潮间带大型底栖动物次级生产力的时空变化. 生态学报, 2011, 31(17): 5071- 5080.
[14] 纪莹璐, 赵宁, 杨传平, 季相星, 王振钟, 隋吉星, 曲方圆, 于子山. 辽东湾西部海域大型底栖动物群落次级生产力初探. 中国海洋大学学报, 2015, 45(4): 53- 58.
[15] Brey T. Population dynamics in benthic invertebrates. A virtual handbook. 2001. http://www.thomas-brey.de/science/virtualhandbook/navlog/index.html.
[16] Robertson A I. The relationship between annual production: biomass ratios and lifespans for marine macrobenthos. Oecologia, 1979, 38(2): 193- 202.
[17] Schwinghamer P, Hargrave B, Peer D, Hawkins C M. Partitioning of production and respiration among size groups of organisms in an intertidal benthic community. Marine Ecology Progress Series, 1986, 31(2): 131- 142.
[18] Tumbiolo M L, Downing J A. An empirical model for the prediction of secondary production in marine benthic invertebrate populations. Marine Ecology Progress Series, 1994, 114: 165- 174.
[19] Dolbeth M, Lillebø A I, Cardoso P G, Ferreira S M, Pardal M A. Annual production of estuarine fauna in different environmental conditions: an evaluation of the estimation methods. Journal of Experimental Marine Biology and Ecology, 2005, 326(2): 115- 127.
[20] 国家海洋局. GB/T 12763.6—2007 海洋调查规范 第6部分: 海洋生物调查. 北京: 中国标准出版社, 2008.
[21] 国家海洋局. GB 17378.4—2007 海洋监测规范 第4部分: 海水分析. 北京: 中国标准出版社, 2008.
[22] Soliman Y S, Rowe G T. Secondary production ofAmpeliscamississippianaSoliman and Wicksten 2007 (Amphipoda, Crustacea) in the head of the Mississippi Canyon, northern Gulf of Mexico. Deep Sea Research Part II: Topical Studies in Oceanography, 2008, 55(24/26): 2692- 2698.
[23] Fuhrmann M M, Pedersen T, Ramasco V, Nilssen E M. Macrobenthic biomass and production in a heterogenic subarctic fjord after invasion by the red king crab. Journal of Sea Research, 2015, 106: 1- 13.
[24] 马长安, 徐霖林, 田伟, 吕巍巍, 赵云龙. 围垦对南汇东滩湿地大型底栖动物的影响. 生态学报, 2012, 32(4): 1007- 1015.
[25] 周细平, 蔡立哲, 梁俊彦, 傅素晶, 林和山, 黄昆. 厦门海域大型底栖动物次级生产力的初步研究. 厦门大学学报: 自然科学版, 2008, 47(6): 902- 906.
[26] 高爱根, 杨俊毅, 陈全震, 胡锡钢, 董永庭, 胡月妹. 达山岛、平岛、车牛山岛邻近海域大型底栖生物分布特征. 海洋学报, 2003, 25(6): 135- 141.
[27] 王金辉, 杨春旺, 孙亚伟, 杨元利. 象山港大型底栖动物的生物多样性和次级生产力研究. 天津农学院学报, 2006, 13(2): 24- 28.
[28] 国家海洋局第二海洋研究所. 浙江省海岛海域生物资源综合调查报告. 杭州: 国家海洋局第二海洋研究所, 1993.
[29] 郑青, 韩海波, 周保学, 李金花, 白晶, 蔡伟民. 化学需氧量(COD)快速测定新方法研究进展. 科学通报, 2009, 54(21): 3241- 3250.
[30] 赵冬梅, 高爱国, 李超, 郭占荣, 张延颇. 闽江下游及河口海域水体中悬浮物的分布特征. 海洋地质前沿, 2012, 28(6): 35- 39.
[31] Nanami A, Saito H, Akita T, Motomatsu K I, Kuwahara H. Spatial distribution and assemblage structure of macrobenthic invertebrates in a brackish lake in relation to environmental variables. Estuarine, Coastal and Shelf Science, 2005, 63(1/2): 167- 176.
[32] Brey T, Jarre-Teichmann A, Borlich O. Artificial neural network versus multiple linear regression: predictingP/Bratios from empirical data. Marine Ecology Progress Series, 1996, 140: 251- 256.
Secondaryproductivityofmacrobenthosintypicalcoastalecosystems:OujiangRiverestuaryandDongtouIslandsasexamples
JIA Shenghua1,2, LIAO Yibo1,2, ZENG Jiangning1,2,*, SHOU Lu1,2, HUANG Wei1,2, GAO Aigen1,2, TANG Yanbin1,2
1KeylaboratoryofMarineEcosystemandBiogeochemistry,StateOceanicAdministration,Hangzhou310012,China2SecondInstituteofOceanography,StateOceanicAdministration,Hangzhou310012,China
To better understand and estimate the secondary productivity of macrobenthos in estuary ecosystems and island ecosystems, we conducted investigations on the macrobenthos in two typical coastal ecosystems, Oujiang River estuary and Dongtou Islands, in the Eastern China sea, in April and October 2015. In Oujiang River estuary, 81 species were found, including 39 polychaete species, 23 crustacean species, 9 mollusk species, 4 echinodermata species, and 6 other species. In Dongtou Islands, 111 species were found, including 50 polychaete species, 30 crustacean species, 18 mollusk species, 5 echinodermata species, and 8 other species. Three different empirical formulas were applied to calculate the secondary productivity of these macrobenthos. The relationships between the secondary productivity and environmental factors were also analyzed. The results showed that: (1) The average annual secondary productivity of macrobenthos, measured as ash-free dry weight and calculated using three empirical formulas, i.e., Brey′s empirical formula (1990) and Brey′s empirical models (2001 and 2012), were 0.48, 0.51, and 0.25 g m-2a-1, respectively, in the Oujiang River estuary and 0.70, 1.55, and 0.99 g m-2a-1, respectively, in the Dongtou Islands. (2) The spatial distribution of the average annual secondary productivity estimated by the three empirical formulas was consistent. A zone with high average annual secondary productivity of macrobenthos was found in the southern waters of the Oujiang River estuary. Two such zones were found in the Dongtou Islands, one located in the sea area between Sanpan Island and Huagang Island, northeast of the main island of Dongtou, and the other in the sea area between the islands southeast of the Donghuang Island. (3) Six species,Aglaophamusdibranchis,Glycerachirori,Heteromastusfiliformis,Potamocorbulaustulata,Yoldiasimilis, andEocylichnabraunsi, mainly contributed—more than 54.2% of the total—to the average annual secondary productivity of macrobenthos, in the Oujiang River estuary. In the Dongtou Islands,A.dibranchis,H.filiformis,Y.similis,Raphidopusciliatus,Eucratecrenata,Cerebratulinasp., andOdontamblyopusrubicunduscontributed the most—more than 57.1% of the total—to the average annual secondary productivity of macrobenthos. (4) Correlation analysis indicated that chemical oxygen demand, suspended solids, and median size of surface sediments were the important environmental factors affecting the average annual secondary productivity of macrobenthos in the Oujiang River estuary. In contrast, environmental factors had no measurable impact on the average annual secondary productivity of macrobenthos in the Dongtou Islands. (5) The average annual secondary productivity of macrobenthos estimated by Brey′s empirical formula (1990) was consistent with that estimated by Brey′s empirical model (2001), for the Oujiang River estuary, and with that estimated by Brey′s empirical model (2012), for the Dongtou Islands.
secondary productivity; macrobenthos; Dongtou Islands; Oujiang River estuary; Brey′s empirical model
海洋公益性行业科研专项(201405007,201505004- 3,201505027- 4);国家海洋局第二海洋研究所基本科研业务费专项 (JG1311,JG1312,JG1412,JG1616);浙江省自然科学基金(LY14D060007);国家科技支撑计划(2015BAD08B01)
2016- 09- 18; < class="emphasis_bold">网络出版日期
日期:2017- 07- 11
*通讯作者Corresponding author.E-mail: jiangningz@126.com
10.5846/stxb201609181872
贾胜华,廖一波,曾江宁,寿鹿,黄伟,高爱根,汤雁滨.近岸典型生态系统大型底栖动物的次级生产力——以瓯江口、洞头列岛为例.生态学报,2017,37(21):7140- 7151.
Jia S H, Liao Y B, Zeng J N, Shou L, Huang W, Gao A G, Tang Y B.Secondary productivity of macrobenthos in typical coastal ecosystems: Oujiang River estuary and Dongtou Islands as examples.Acta Ecologica Sinica,2017,37(21):7140- 7151.
