油茶象甲研究进展
2017-11-17朱桂兰曾爱平
马 玲,朱桂兰,曾爱平
(湖南农业大学植物保护学院,植物病虫害生物学与防控湖南省重点实验室,湖南 长沙 410128)
油茶象甲研究进展
马 玲,朱桂兰,曾爱平
(湖南农业大学植物保护学院,植物病虫害生物学与防控湖南省重点实验室,湖南 长沙 410128)
油茶象甲是我国特有的一种天然木本油料树种—油茶的蛀果害虫,分布于我国大部分油茶产区,危害油茶引起落果,给油茶生产带来严重损失。本文综述了油茶象甲的发生分布、危害及其影响因子、生物学特性、导致落果原因和综合防治等,旨在为油茶象甲的治理提供理论依据。
油茶象甲;综合防治;研究进展
油茶(Camellia spp.)又称茶籽树,一般指种子含油率较高的山茶属植物,是世界四大木本油料树种之一[1],历史悠久。目前全世界已知 280种,其中我国有 85%[2],主要分布在长江流域以南[3]。茶籽油是绿色健康高档的食用油[4],我国占世界油茶总资源的 80%[5]。油茶果实种仁营养丰富,脂肪含量 50% 以上,是动物重要的食物来源[6-7]。油茶生长期易受多种害虫危害[8-10],导致其减产甚至绝收,其中为害果实的害虫主要是油茶象甲[11]。油茶象甲(Curculio chinensis Chevrolat),又名山茶象、中华茶籽象,属鞘翅目(Coleoptera)象甲科(Curculionidae),是油茶专性蛀果害虫,名列我国林业危险性有害生物之中[12],大部分油茶产区均有分布[13],引起油茶品质下降[14],造成大量落果[15],最高可达96.6%[16],严重影响我国油茶产业的发展。
1 油茶象甲生物学研究
1.1 生活史
油茶象甲严重危害油茶,但由于生产中管理粗放,且油茶象甲生活隐蔽,对其生物学方面的研究一直较少。该虫生活史极不整齐,全国多数油茶产区 2 年 1 代,跨 3 个年头。第 1 年幼虫筑土室越冬,第 2 年以成虫越冬为主,第 3 年越冬成虫 4—6 月出土为害[15]。
1.2 生物学习性
马玲等[17]调查发现油茶象甲成虫雌雄比3.06 ∶ 1,具黑色和褐色两种体色型,黑色型通体黑色,褐色型仅前胸背板、喙基部黑色,其余淡褐色或红褐色,且黑色型为优势种群。
油茶象甲通过取食和产卵危害油茶果实。成虫取食时,最先用喙端的口器咬住果实,以此为着力点,身体在果实表面来回旋转,然后喙钻入果皮伸至种仁内部,取食种子汁液或咬食种仁。5 月中旬到 6 月上旬为成虫出土盛期,产卵盛期在 6 月中旬至 7 月上、中旬。成虫产卵时,通过产卵器将卵产入取食孔中,然后调转身体由喙将其推入种内或种仁间隙,卵期 10—15 天[15,18]。幼虫蛀食种仁,6 月下旬至 7 月上旬幼虫出果,出果盛期在 7 月中旬至 9 月中旬,幼虫单果发育,果内发育期为 16—21 天,平均 18.2 天[15],老熟后将果实咬出 3 ~ 5 mm 的圆孔脱果而出[18],于潮湿松软土壤滞育、化蛹及羽化[15]。虫蛀落果收集后幼虫逃逸数量逐天递减,逃逸高峰期集中在 6∶ 00—10∶ 00 及 16∶ 00—18∶ 00[15],体质量越大,幼虫逃逸时间越早[11]。一般认为,油茶象甲幼虫可分为 4 龄[15],但李志文等[18]野外种群研究表明,油茶象幼虫具 5 龄。成虫除白天活动外,晚上也有取食产卵行为[19],喜阴湿,有假死性,飞翔能力弱,产卵期在 150 天左右,寿命长达 250—300 天[15]。
油茶象甲发生危害轻重与其本身的特性、林间小气候、油茶品种以及管理密切相关。单一种植林重于混林,郁闭度大的油茶林受害较重,老林受害重于幼林。在地形上,丘陵、半山坡危害重于山坡,阴坡多于阳坡。不同林木品种,受害程度亦稍有差异。油茶象甲幼虫呈聚集分布,油茶植株不同高度层次,分布密度有明显差异,下层危害最为严重,中上层较轻[5,20]。蔡守平等[21]报道,小果油茶比普通油茶受害程度高。早熟寒露品种受害较轻,迟熟霜降品种则较重。迟熟品种中,“紫红球”和“紫红桃”抗虫性较好、青皮类型受害最严重[22-23]。赵丹阳等[20]测试油茶象甲雌虫对油茶不同部位的趋性反应发现,4 月份油茶象甲雌虫对果实趋性低于嫩芽,而在 6 月份对果实趋性则大于嫩芽和成叶。
2 油茶象甲发生危害
2.1 野外危害调查方法
油茶象甲的危害调查普遍采用五点取样法或平行取样法;成虫采集利用其假死性采取震落方式,少数利用网捕法;通过采取果实或捡拾落果调查幼虫危害[11,21,24-25]。
2.2 影响油茶象甲危害的关键因子
油茶象甲以成虫或幼虫取食危害油茶果实。幼虫整个取食期均在单个果实内进行[11],故其危害性实际上取决于成虫取食、产卵行为。影响油茶象甲雌虫产卵的生物因子主要有寄主果实特征及天敌,非生物因子为温度。
动物对果实(种子)的扩散是一个复杂的生态学过程,常常受到果实特征的影响[26]。内食类昆虫需咬破寄主表面,取食植物体内部组织,因此植物的形态结构就会影响昆虫行为[27]。山茶属(Camellia)植物果实特征主要包括果实大小(果长和果径)、果实厚度(壳厚)、颜色、形状等[28]。其中果实大小、果实厚度与昆虫寄生的关系尤为密切[29-30]。
(1)果实大小(果径)。油茶象甲是寄主植物的专性蛀果昆虫,除成虫出土早期受蜜源植物如金银花(Lonicera sp.)和白背桐(Mallotus aelta)吸引外[31],无论幼虫还是成虫均以寄主果实为食。特别是幼虫,整个发育期均在果实内进行。油茶果实特征变异极其普遍[32],果实大小不同,营养物质含量相差甚大,因此果实大小成为幼虫发育的主要限制因子[33-34]。大果实出果的幼虫体质量更大,小果实出果的幼虫大小则受限制[11]。体质量越大,潜在适合度随之增加[35-36]。大量研究表明,昆虫为了提高后代适合度,亲代雌虫选择大果实产卵[37-42]。赵丹阳等[43]发现油茶象甲对油茶果径小于 13 mm 的果实不产卵。油茶象甲产卵高峰期野外调查和成虫室内测定,发现其偏好选择大果实产卵且具有“卵单产”特性[19]。但对小果油茶和普通油茶混合林调查结果表明,油茶果径 2.1 ~ 3.0 cm 的中小果实受害更重[21]。何立红等[44]对小果油茶落果研究发现,随着油茶果实大小的递减,油茶象甲幼虫寄生率相应增加。这可能是因为被寄生的油茶果实发育缓慢,非危害果实则发育相对较快,从而导致大果实危害率反而较低。
(2)果实生理状态。植物受害后,释放的挥发物会发生改变,生理状态随之而变,从而影响昆虫行为,导致昆虫对不同生理状态下的寄主植物偏好性不同。油茶象甲危害非病害果显著高于病害果实[44],选择危害健康状态果实,可能是油茶健康果含有某种(类)化学物吸引油茶象甲,而油茶病害果实的某些挥发物则对油茶象甲存在驱避作用,譬如西印度甘薯小象甲偏好选择未受害块茎[45],马铃薯甲虫对被沙漠蝗为害的马铃薯植株更具趋向性[46]。另一方面,病害果实通常表面有病斑,种子发霉,果实品质差,这可能影响油茶象甲行为选择。
(3)油茶象甲天敌。目前有关油茶象甲天敌的报道很少[20,25],已有研究表明,啮齿动物能识别虫蛀果实,并取食该虫蛀种子剩余可利用的资源,包括其内的象甲[6]。在都江堰实验林场,小泡巨鼠(Leopoldamys edwardsi)有搬运取食普通油茶的现象[47]。在益阳赫山区小果油茶林,落果中油茶象甲幼虫有选择地被某种鼠类取食,该鼠类也取食较小的油茶果实[48]。
(4)温度。温度是昆虫生长发育的重要因素,在繁衍过程中起着决定性作用。明确气候对昆虫生存和繁殖的作用,可以预测其种群发展动态,为害虫综合防治预测预报提供理论依据[49-50]。不同昆虫种类之间,温度的作用各异。发育温度内,低温通常会延长各虫态发育时间,高温则会缩短昆虫发育甚至致死[51]。潘华平[52]研究表明 3—4月、7—8 月平均气温和平均相对湿度以及 5—6 月≥ 10 ℃ 的有效积温影响油茶象甲危害发生。马玲等[17]发现 7 月下旬至 8 月初持续高温(日最高温度>35 ℃)导致油茶象甲成虫大量死亡,成虫日死亡数与日最高气温显著线性正相关。
2.3 油茶象甲导致果实脱落原因
被象甲寄生的果实成熟前下落是一种普遍现象[53-54]。受油茶象甲危害的果实在果长、果径、果重及籽重方面显著小于非受害果[21]。有学者认为对植物而言,停止虫蛀果实生长,主动防御将节约下来的能量更好地分配到健康果实中,对种群更新具重要意义。而李志文等[48]研究表明,油茶象甲蛀果为害引起果实正常生理受阻才导致其脱落,并非符合主动防御假说。
3 油茶象甲综合防控现状
油茶象甲是我国重要木本油料植物—油茶的蛀果害虫,该虫果内产卵,土中化蛹,生活隐蔽,天敌较少,防治特别困难。近年来,油茶象甲防控措施尚未完善。
3.1 林业防治
选育抗虫油茶品种,抚育幼林,改造老林,垦复培育,及时采果,捡拾落果对其集中焚烧,减少小年发生[15]。
3.2 物理防治
植物挥发物或聚集信息素是一种高效防治害虫的方法。在油茶幼林中间作大豆、花生对调控油茶炭疽病、软腐病、烟煤病以及油茶尺蠖、毒蛾等油茶主要病虫害效果明显[55-56]。王宗楷等[31]在广西桂林利用金银花(Lonicera sp.)和白背桐(Mallotus aelta)成功引诱油茶象甲。将特定配比的糖醋液挂在油茶树上,对油茶象甲也有很好的诱集作用[22]。
3.3 化学防治
油茶象甲防治期间,可选用绿色威雷、乐果乳油、敌百虫、醚菊酯乳油、三唑磷乳油、敌敌畏等药剂进行喷雾,密集油茶林可使用烟雾剂熏杀(各农药的有效作用浓度见表 1)[15,22,57-58]。幼虫出果期于地面撒施药粉、石灰或喷洒 90% 敌百虫 500 倍液对其灭杀[15]。
3.4 生物防治
虫生真菌等对害虫的防治同样起重要作用。绿僵菌和白僵菌对象甲具有持续控制作用[59-62],其中油茶林喷施菌株 FJMa 201101 后,油茶象甲幼虫死亡率随着时间推移逐渐提高,喷菌后 180 天,平均死亡率超过 70%[61]。
4 展望
油茶象甲的生活史大部分在土中,果内产卵,土中化蛹,生活隐蔽,土壤翻耕对其影响很大。但由于人们对于防治知识认识不足疏于管理,再加上劳动力成本增加,垦复翻耕和捡拾落果等的林间防治越来越少,因此,防治油茶象甲应更重视林业防治,营林护林,及时清理落果。油茶产业绿色发展要求安全无公害的防治措施,减少环境污染。针对油茶象甲老熟幼虫入土1年以上才化蛹羽化的特性,土中治虫相比于其他生活史阶段更为容易。防治油茶象甲期间,应重点对生物杀虫剂白僵菌、绿僵菌的研发,筛选出油茶象甲幼虫致病力优良菌株。但生物菌株开发成本高,效果缓慢,环境因素影响较大,推广应用存在难度。
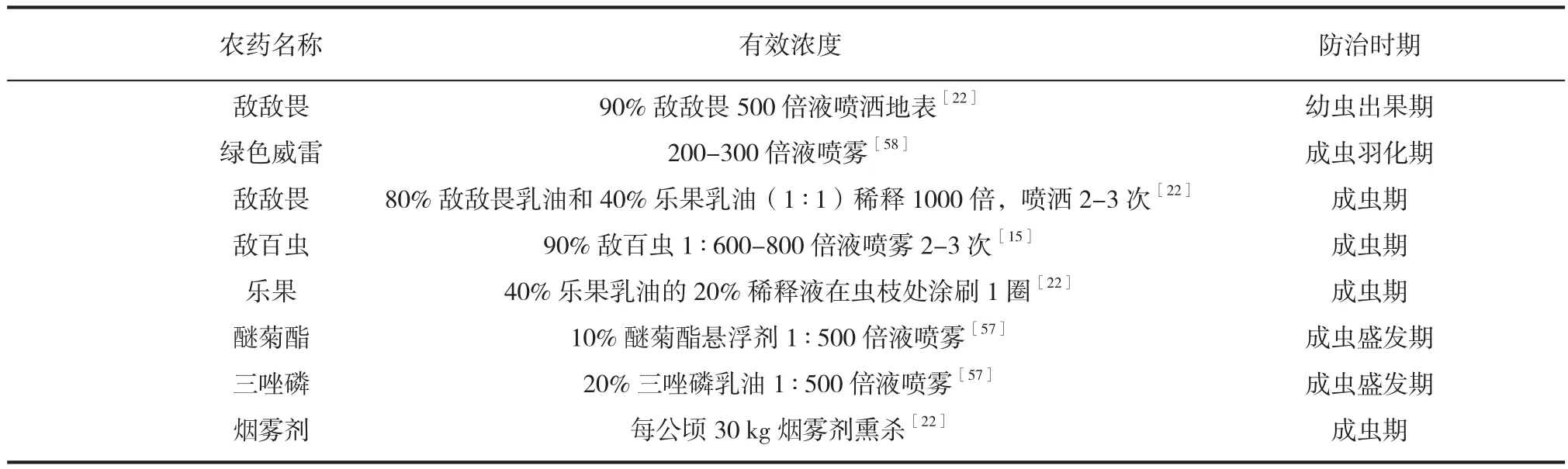
表1 防治油茶象甲常用农药及其有效浓度Tab.1 Common pesticide and its effective concentration of controlling Curculio chinensis Chevrolat
鞘翅目昆虫能够分泌聚集信息素,并且已有利用信息素与挥发物成功防治的案例。如 Bartelt和 Hossain 利用植物挥发物配制一种取食引诱剂,用于治理澳大利亚梨园甲虫[63];刺激多种叶甲取食的葫芦素,与杀虫剂和引诱剂混合制成毒饵,能较好的防治食根叶甲[64]。国内外利用化学信息物质对油茶象甲防治研究鲜见报道,因此对油茶象甲信息素与油茶树或山茶属植物的活性研究需要迫切。如果能将信息素和挥发物结合,大量诱捕油茶象甲,既能防治害虫又可减少化学农药污染,避免害虫抗药性和再猖獗。因此,综合应用信息素诱捕和监测,对防控油茶象甲具有广阔的应用前景。同时,油茶象甲作为一种专性油茶蛀果害虫,在已有研究的基础上,运用生理生化和分子生物学技术,揭示油茶象甲与寄主植物协同进化是今后的又一个重要研究方向。
[1]庄瑞林.中国油茶(第2版)[M].北京:中国林业出版社,2008.
[2]张宏达.中国植物志[M].北京:科学出版社,1998.
[3]刘 凌,泽桑梓,季 梅,等.中国油茶林害虫和螨类及其天敌研究综述[J].西部林业科学,2013,42(1):96-104.
[4]丛玲美.茶油品质控制过程中主要质量指标变化规律的研究[D].北京:中国林业科学研究院,2007.
[5]周国英,宋光桃,李 河.油茶病虫害防治现状及应对措施[J].中南林业科技大学学报:自然科学版,2007,27(6): 179-182.
[6]肖治术,张知彬,王玉山.小泡巨鼠对森林种子选择和贮藏的观察[J].兽类学报,2013,23(3):208-213.
[7]Abe H,Matsuki R,Ueno S,et al.Dispersal of Camellia japonica seeds by Apodemus speciosus revealed by maternity analysis of plants and behavioral observation of animal vecto rs[J].Ecology Research, 2006(21):732-740.
[8]洪 伟,罗顺跃,陈顺立,等.油茶主要病虫害生态空间分布规律的研究[J].应用生态学报,1992,3(4):308-312.
[9]黄敦元,余江帆,郝家胜,等.不同生境油茶林油茶史氏叶蜂的发生与危害程度比较[J].中南林业科技大学学报,2010, 30(1):59-64.
[10]束庆龙.中国油茶栽培与病虫害防治[M].北京:中国林业出版社,2009.
[11]李志文,何立红,马 玲,等.果实大小对油茶象幼虫生长的影响[J].应用生态学报,2014,25(12):3580-3586.
[12]国家林业局.国家林业局 2013 年第 4 号公告[EB/OL].http∶//www.forestry.gov.cn/uploadfile/main/2013-1/file/2013-1-14-fc1f 01fa05cb445ea6febed5fdfd86b9.pdf
[13]徐 磊,潘涌智,黄阳洋.油茶象的潜在分布区预测[J].福建林业科技,2011,38(4):55-58.
[14]李苗苗,张 威,吕军美,等.茶籽象为害对油茶果产量和茶油品质的影响[J].植物保护,2016,42(5):65-68.
[15]周石涓.油茶象的生物学及其防治[J].昆虫学报,1981, 24(1):48-52.
[16]蒋三俊.油茶象鼻虫的防治[J].特种经济动植物,2009, 12(8):54-54.
[17]马 玲,曾爱平,李志文.油茶象生物学初步研究[C].华中昆虫研究,2015(11)∶ 132-138.
[18]李志文,何立红,夏 姣,等.油茶象幼虫虫龄的划分[J].昆虫学报,2015,58(2):181-189.
[19]李志文,何立红,杨柳君,等.小果油茶上油茶象的产卵策略[J].昆虫学报,2015,58(9):981-988.
[20]赵丹阳,秦长生,徐金柱,等.油茶象甲成虫对油茶寄主选择性研究[J].中国农学通报,2015,31(17):100-104.
[21]蔡守平,何学友,李志真,等.油茶象危害油茶果实的初步研究[J].福建林业科技,2011,38(2):14-16.
[22]周 宏.油茶象生物学特性与防治方法[J].安徽林业科技,2007(3):48-48.
[23]郑学慧,郎南军,黄 钰.油茶病虫害发生的现状及其环境因子研究[J].西部林业科学,2011,40(4):106-110.
[24]赵丹阳,秦长生,揭育泽,等.广东省油茶病虫害种类及发生动态调查[J].安徽农业科学,2012,40(29):14267-14270.
[25]李 密,周 刚,彭争光,等.湖南油茶害虫风险性评估及危险性等级划分[J].中国农学通报,2014,30(19):277-283.
[26]Bonal R,Muñoz A.Seed growth s uppression constrains the growth of seed parasites:premature acorn abscission reduces Curculio elephas larval size[J].Ecological Entomology,2008,33(1):31-36.
[27]朱更瑞,王新卫,方伟超,等.桃园梨小食心虫发生规律及成虫对不同诱捕器的趋性研究[J].果树学报,2014(2):282-287.
[28]何 方,吕芳德.湖南油茶品种分类及优良类型的研究[J].经济林研究,1985,3(1):1-26.
[29]Hughes M,Dobric N,Scott I C,et al.T he Hand1, Stra13 and Gcm1 t ranscription factors override FGF sig naling to promote terminal differentiation of trophoblas t stem cells[J].Developmental Biology,2004,271(1):26-37.
[30]Toju H,Sota T.Imbalance of predator and prey armament:geographic clines in phenotypic interface and natural selection[J].The American Naturalist,2006,167(1):105-117.
[31]王宗揩,丘凤波,尤其儆.山茶象的植物诱集[J].植物保护,1980,6(2):11-13.
[32]黄 勇.小果油茶与普通油茶居群遗传结构及种间杂交渐渗[J].应用生态学报,2011,24(8):2345-2352.
[33]Lafferty K D,Kuris A M.Trophic strategies, animal diversity and body size[J].Trends in Ecology & Evolution,2002,17(11):507-513.
[34]Desouhant E,Debouzie D.Ploye H,et al.Clutch size manipulations in the chestnut weevil,Curculio elephas:fitness of oviposition strategies[J].Oecologia,2000,122(4):493-499.
[35]Reitz S R,Adler P H.Fecundity and ovipoasition of Eucelatoria bryani,a gregarious parasitoid of Helicoverpa zea and Heliothis virescens[J].Entomologia Experimentalis Et Applicata,1995, 75(2):175-181.
[36]Bonal R,Hernández M,Ortego J,et al.Positive cascade effects of forest fragmentation on acorn weevils mediated by seed size enlargement[J].Insect Conservation & Diversity,2012,5(5):381-388.
[37]Fenner M,Cresswell J,Hurley R,et al.Relationship between capitulum size and pre-dispersal seed predation by ins ect larvae in common Asteraceae[J].Oecologia,2002,130(1):72-77.
[38]Gao S,Tang Y,Ke W,et al.Rela tionships between body size and parasitic fitness and offspring performance of Sclerodermus pupariae Yang et Yao (Hymenoptera:Bethylidae)[J].Plos One,2016,11(7):1-14.
[39]詹月平,周 敏,贺 张,等.寄主大小及寄生顺序对蝇蛹佣小蜂寄生策略的影响[J].生态学报,2013,33(11): 3318-3323.
[40]高珏晓,孟 玲,李保平.广大腿小蜂对菜粉蝶蛹体型大小的产卵选择及后代发育表现[J].生态学杂志,2010,29(2):339-343.
[41]刘文静,汪广垠,牛可坤,等.槲栎种子雨进程中昆虫的捕食特征[J].昆虫学报,2010,53(4):436-441.
[42]王 学,肖治术,张知彬,等.昆虫种子捕食与蒙古栎种子产量和种子大小的关系[J].昆虫学报,2008,51(2):161-165.
[43]赵丹阳,秦长生,徐金柱,等.油茶象甲形态特征及生物学特性研究[J].环境昆虫学报,2015b,19(1):3-6.
[44]何立红,李志文,刘劲军,等.油茶象危害与小果油茶果实特征的相关性[J].林业科学,2014,50(12):151-155.
[45]Yasuda K.Attractiveness of different plant organs of sweet potato and damaged tuber to West-Indian sweet potato weevil,Euscepes postfasciatus(Fairmaire)(Coleoptera:Curculionidae)in wind tunnel[J].Japanese Journal of Applied Entom olgy and Zoolgy, 1996,40(2):160-161.
[46]Dicke M,Sabelis M W,Takabayashi J,et al.Plant strategies of manipulating predatorprey interactions through allelochemicals:Prospects for application in pest control[J].Journal of Chemical Ecology,1990,16(11):3091-118.
[47]Xiao Z S,Zhang Z P,Krebs C J.Long-term seed survival and dispersal dynamics in a rodent-dis persed tree:testing the predator satiation hypothesis and the predator dispersal hypothesis[J].Journal of Ecology,2013,101(5):1256-1264.
[48]李志文,孙寒松.湖南益阳小果油茶林小果油茶、油茶象和啮齿动物的相互作用[J].昆虫学报,2016,59(10):1123-1132.
[49]Carey J R.Applied demography for biologists with specia lemphasis on insects[J].Oxford University Press,1993,118(2):243-243.
[50]Enkegaard A.The poinsettia strain of the cotton whitefly,Bemisia tabaci(Homoptera:Aleyrodidae),biological and demographic parameters on poinsettia(Euphorbia pulcherrima)in relation to temperature[J].Bulletin of Entomological Research,1993,83(4):535-546.
[51]丁岩钦.昆虫种群数学生态学原理与应用[M].北京:科学出版社,1980.
[52]潘华平.油茶主要病虫害防控区域划分及其预测预报研究[D].长沙:中南林业科技大学,2011.
[53]Boucher D H,Sork V L.Early drop of nuts in response to insect infestation[J].Oikos,1979,33(3):440-443.
[54]Gurnell J.Tree seed production and food conditions for rodents in an oak wood in southern England[J].Forestry,1993,66(3): 291-315.
[55]陈守常,田泽君,郭隆錫.油茶炭疽病的发生与蔓延[J].植物保护学报,1965(3):207-218.
[56]文亚雄,刘君昂,刘小平,等.不同间作模式对油茶幼林病虫害发生的影响[J].经济林研究,2016,34(1):129-134.
[57]杜月飞.油茶象发生现状与防治对策[J].防护林科技,2014(10):123-124.
[58]邓卓喜.油茶病虫害发生现状及其防治措施[J].热带林业,2011,39(4):42-46.
[59]Adane K,Moore D,Archer S A.Preliminary st udies on the use of beauveria bassiana to control Sitophilus zeamais(Coleoptera:Curculionidae)in the Laboratory[J].Journal of Stored Products Research,1996,32(2):105-113.
[60]Shah F A,Gaffney M,Ansari M A,et al.Neem seed cake enhances the efficacy of the insect pathogenic fungus Metarhizium anisopliae for the control of black vine weevil,Otiorhynuchs sulcatus (Coleoptera:Curculionidae)[J].Biological Control,2008,44(1):111-115.
[61]邓小军,周国英,刘君昂,等.湖南油茶林丛枝菌根真菌多样性及其群落结构特征[J].中南林业科技大学学报∶自然科学版,2011,31(10):38-42.
[62]何学友,蔡守平,杜月飞,等.感染油茶象幼虫的高致病力金龟子绿僵菌菌株筛选[J].林业科学,2015(8):52-59.
[63]Bartelt R J,Hossain M S.Development of s ynthetic foodrelated attractant for Carpophilus davidsoni and its effectiveness in the stone fruit orchards in southern Australia[J].Journal of Chemical Ecology,2006,32(10):2145-2162.
[64]Metcalf R L,Ferguson J E,L ampman R,et al.Dry cucurbitacin-containing baits for controlling diabroticite beetles(Coleoptera:Chrysomelidae)[J].Journal of Economic Entomology,1987, 80(4):870-875.
(文字编校:龚玉子)
Research advance on Curculio chinensis Chevrolat
MA Ling,ZHU Guilan,ZENG Aiping
(Hunan Provincial Key Laboratory for Biology and Control of Plant Diseases and Insect Pests,College of Plant Protection,Hunan Agricultural University,Changsha 410128,China)
The camellia weevil,Curculio chinensis Chevrolat,belongs to Coleoptera Curculionidae,and it is a vital pest attacking fruits of the oil tea camellia in China.Camellia weevil is distributed in oil-tea camellia production of China.Its infestation always leads to tremendous fruit drop of plants,which causes a huge los s of camellia output.This article preliminary summarizes the research advance on the occurrence and distribution,harm and its impact factors,biological characteristics,reasons of causing fruit drop,as well as integrated pest management of the camellia weevil.It aims at providing theoretical basis for identification and prediction as well as comprehensive management of Curculio chinensis Chevrolat.
Curculio chinensis Chevrolat;comprehensive management;research advance
S 763.7
A
1003-5710(2017)03-0084 -06
10.3969/j.issn.1003-5710.2017.03.015
2017-04-05
湖南省现代农业产业技术体系(湘农业联[2012]278号);湖南省科技厅项目(2011FJ4298);湖南省教育厅项目(15C0672);湖南农业大学创新实验(XCX17025)
马 玲(1993-),女,在读硕士,湖南省汨罗市人,研究方向为农业昆虫与害虫防治
曾爱平,教授;E-mail:apzengchina@163.com
