家兔ANGPTL4基因多态性及其与生长性状的关联性分析
2017-11-16李彦宏胡深强贾先波赖松家
李彦宏,胡深强,王 杰,贾先波,赖松家
(四川农业大学动物遗传育种研究所,四川 成都 611130)
家兔ANGPTL4基因多态性及其与生长性状的关联性分析
李彦宏,胡深强,王 杰,贾先波,赖松家*
(四川农业大学动物遗传育种研究所,四川 成都 611130)
【目的】探究家兔血管生成素样蛋白4(angiopoietin-like protein 4, ANGPTL4)基因第6外显子内的多态性及其与家兔部分生长性状的关联性。【方法】采用PCR产物测序法检测家兔ANGPTL4基因外显子内的单核苷酸多态性(SNPs)位点,并应用PCR-RFLP酶切分型技术对3个家兔品种(天府黑兔、伊拉兔和新西兰兔)共495个群体样本进行酶切分型,分析SNPs与3个家兔品种部分生长性状的关联性。【结果】ANGPTL4基因第6外显子中共检测发现3个遗传变异位点(c.823 G>A为非同义突变,c.867 T>C和c. 975 T>C均为同义突变),且完全处于连锁状态 (r2≥0.75),由此选择c.823 G>A位点作为遗传效应分析位点。关联性分析得出3个家兔群体内AG基因型和GG基因型与3个品种部分生长性状具有较高的遗传效应。【结论】ANPGTL4基因可以是与家兔生长发育相关的候选基因,以此为培育肉兔新品种提供辅助分子遗传选择依据。
ANGPTL4基因;多态性;生长性状;关联性分析;家兔
【研究意义】血管生成素样蛋白4(Angiopoietin-like protein4,ANGPTL4)被许多研究证实并归类于类血管生成素家族,是调控脂肪细胞分化的重要信号分子。而在家兔ANGPTL4基因研究中已被测出全长为4971 bp,且含7个外显子,同时可以编码411个氨基酸。早期研究发现ANGPTL4蛋白的表达具有选择性,分别在脂肪组织和胚胎中集中表达。而在不同来源的脂肪组织中表达也不一样,如只表达于内脏脂肪组织,其他器官脂肪组织(心脏、肝脏、肺、肾)少量表达[1]。ANGPTL4对脂肪组织的形成起抑制作用,其原因有可能是ANGPTL4蛋白产生水解作用,进一步缠绕折叠成卷曲螺旋结构域和纤维蛋白原样结构的小片段产物,从而引起脂肪细胞分化发生改变[2]。脂肪酸的氧化和解耦联作用已被证实对脂肪组织降解起重要作用。在脂肪组织形成的过程中,过氧化物酶体增殖物激活受体(PPAR-α、PPAR-β和PPAR-γ)可以在脂肪组织中产生解耦联作用降低脂质的含量,而ANGPTL4基因在脂肪组织中过表达后可以通过调节脂肪酶的活性去调节脂肪代谢,从而调节器官组织内的脂肪酸和甘油三酯的形成,使组织内脂肪含量得到有效降解[3-5]。【前人研究进展】Yoon等[6]的研究表明ANGPTL4是PPAR-α和PPAR-γ的下游因子,对血管生成和代谢具有重要促进作用。在肝脏、心脏、骨骼肌和肠中, PPAR-α、PPAR-γ和缺氧诱导因子HIF-1(hypoxiain duciblefa)可以通过激活ANGPTL4基因的转录,调控脂类物质的代谢[7]。ANGPTL4基因的cSNPs位点已在人类和小鼠中发现其与体脂率的高低有明显的相关性。legry等[4]在人的研究中发现ANGPTL4基因rs4076317位点的具有多态性,并与青年的体脂率显著性相关联,这些多态性位点均与PPARγ表达高低有关。Kim等[8]也在小鼠的中发现ANGPTL4基因表达水平与小鼠的体重升高呈正相关。【本研究切入点】因此,ANGPTL4基因对动物机体的脂肪能量代谢以及生长发育中起重要作用。由此ANGPTL4基因可能与家兔的生长性状可能存在密切联系。【拟解决关键问题】由此检测了家兔ANGPTL4基因的遗传变异位点,并分析了与3个家兔品种生长性状的联系,为家兔分子标记辅助育种提供理论依据。
1 材料与方法
1.1 试验材料
本试验所选3个肉兔品种: 新西兰兔172只、天府黑兔165只、伊拉兔158只。所有试验家兔均随机从四川农业大学教学基地选取健康无疾病、3代之内无亲缘关系的家兔。所选3个家兔群体的饲养和管理均保持一致,并做好防疫工作。所购饲料购自四川金川农饲料有限公司的肉兔全价颗粒饲料,试验期间家兔自由采食饮水,并测定35、42、56、70和84日龄5个阶段的个体体重数据。试验结束后剪取家兔部分耳组织块,放入添加有75 %酒精的1.5 mL的无菌EP管内,并尽快置于-20 ℃冰箱保存备用。
1.2 基因组DNA的提取
本试验使用DNA试剂盒(TIANGEN, Beijing)对3个家兔品种群体内共计495份样本的DNA进行了提取,并使用核酸蛋白分析仪分析DNA浓度(OD260/OD280在1.7~1.8)和1.5 %琼脂糖电泳实验分析DNA的提取效果,提取完毕使其储存于-20 ℃低温冰箱备用。
1.3 引物设计和PCR扩增
参照在NCBI上家兔ANGPTL4基因的DNA序列(GenBank 登录号:NW_003159393),利用Primer Premier5.0软件对ANGTL4基因外显子6设计引物(表1)。
随机选取30个DNA样本进行PCR扩增。PCR扩增体系为10 μl,其中包含5 μl 2×TaqPCR MasterMix,F/R引物各0.4 μl,DNA模板1 μl以及3.2 μl灭菌超纯水,PCR反应程序为:95 ℃预变性5 min,95 ℃变性30 s,退火30 s,72 ℃延伸50 s,38个循环;72 ℃延伸10 min,12 ℃保存。以上引物和所得PCR扩增产物均由成都市擎科生物有限公司进行引物的合成和产物的测序。
1.4 PCR产物测序和SNP位点筛选
随机选取3个家兔品种群体内30个DNA样本,对ANGPTL4基因的第6外显子进行PCR扩增,然后收集样品的PCR产物在成都市擎科生物有限公司进行纯化验证,并使用SeqMan软件和Chromas软件分析变异位点。应用Haplo View软件使所测SNP位点的连锁不平衡得到确定。
1.5 PCR-RFLP基因分型
本试验使用在线软件WatCut(http://watcut.uwaterloo.ca/template.php?act=snp_new)对所需限制性内切酶的进行初步选择,并使用Primer Premier5.0确定其特异性,由此本试验优先选择Cfr42I酶(SacII)(货号:ER0202)(购自赛默飞世尔科技公司)使遗传变异位点得到准确的酶切分型结果。酶切体系为10 μl:10×FastDigest Green Buffer 1.0 μl,20×Oligonucleotide(0.01 mM) 0.5 μl,PCR产物3.0 μl,Cfr42I (SacII)酶 0.3 μl,ddH2O 5.2 μl。然后酶切体系轻轻摇匀离心后静置于37 ℃恒温水浴锅中60~80 min,完毕后取出所得混合物,采用琼脂糖凝胶电泳(1.5 %)检测PCR产物片段酶切结果,并在胶图上做好基因型标记,以便用于统计分析。
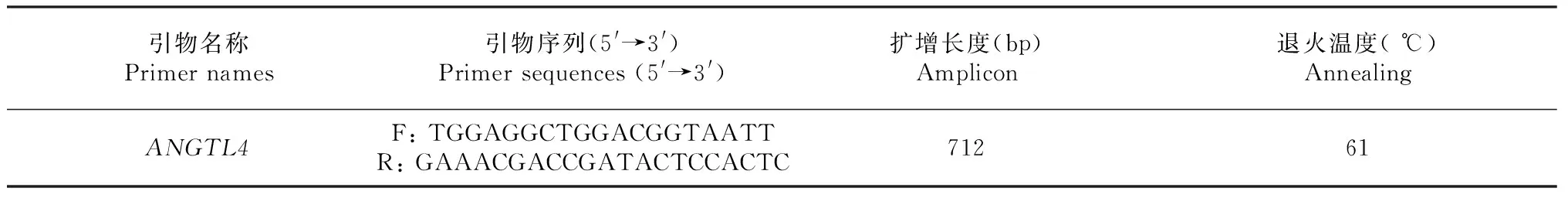
表1 PCR引物引物信息
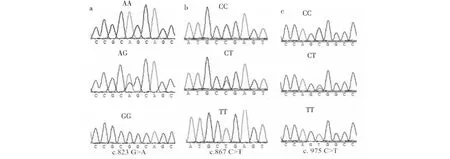
小写字母a、b和c分别代表c.823 G>A、c.867 T>C和c. 975 T>C The lowercase abc represent the sites c.823 G>A, c.867 T>C and c. 975 T>C, respectively图1 ANGPTL4基因测序结果Fig.1 The sequencing consequence of ANGPTL4 gene

图2 ANGPTL4蛋白结构域预测Fig.2 The domain prediction of ANGPTL4 protein
1.6 数据处理
本试验使用SPSS 22.0软件中的一般线性模型(general linear model,GLM)程序用于分析ANGPTL4基因的SNP位点与3个家兔品种的生长性状之间的遗传效应,数据分析结果用最小二乘均数±标准误(mean ± SE)表示。
Yijk=μ+Gi+Bj+Sk+eijk
其中:Yijk和Yjk代表个体生长性状测量值;μ代表群体平均数;Gi代表基因型效应;Bj代表品种效应;Sk代表性别效应;eijk代表随机误差。
2 结果与分析
2.1 ANGPTL4基因SNP位点检测
如图1所示,第6外显子内存在3个遗传变异位点(c.823 G>A、c.867 T>C和c. 975 T>C),其中c.823 G>A为错义突变,c.867 T>C和c. 975 T>C均为同义突变。
2.2 家兔ANGPTL4基因遗传位点分析
如图2所示,3个遗传变异位点(c.823 G>A,c.867 T>C和c. 975 T>C)均位于FBG结构域中,这3个突变位点可能会对ANGPTL4蛋白的功能产生影响,由此需要进一步对SNPs进行连锁平衡分析。
如图3所示,c.823 G>A与c.867 T>C、c. 975 T>C处于完全连锁状态,且具有相似的遗传效应,其中c.823 G>A与其余两个SNPs连锁程度非常强(r2值分别为0.89和0.82)。由此本试验最后选择c.823 G>A点作为ANGPTL4基因的候选SNP进行后续研究。
2.3 PCR-RFLP分型
利用PCR-RFLP分别对天府黑兔、伊拉兔和新西兰兔共计495份样本DNA中ANGPTL4基因c.823 G>A位点进行酶切分型。所使用限制性内切酶为Cfr42I (SacII)(货号:ER0202)购于赛默飞世尔科技公司(Thermo Fisher Scientific)。使用1.5 %琼脂糖凝胶电泳所得酶切分型结果如图4所示,依次分别呈现GG基因型、AA基因型和AG基因型。
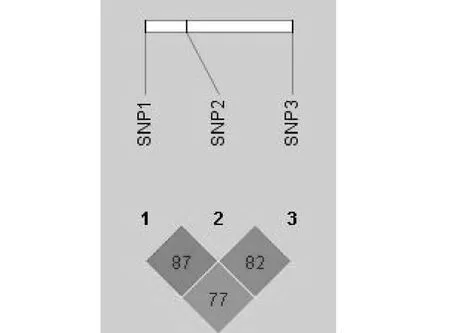
SNP1-3分别代表c.823 G>A、c.867 T>C和c. 975 T>CSNP1-3 represent c.823 G>A, c.867 T>C and c. 975 T>C图3 ANGPTL4基因SNPs 连锁不平衡分析Fig.3 Linkage disequilibrium analysis of SNPs in ANGPTL4 gene
2.4 家兔ANGPTL4基因SNP位点与生长性状的相关性
通过统计所得c.823 G>A位点的基因频率、基因型频率和遗传参数及卡方检验χ2的统计结果。
如表2所示,c.823 G>A位点分别在3个家兔品种内发现G和A等位基因和AA、AG和GG基因型,其中G等位基因和GG基因型均具有优势效应。而c.823 G>A遗传多态性位点在每个家兔群体内的PIC均为中度多态(0.25
2.5 c.823 G>A位点与家兔生长性状的关联性分析
对c.823 G>A位点与3个品种家兔群体样本的35~84日龄阶段的个体重进行关联性分析(表3),表明在天府黑兔群体中AG基因型样本群体在5个生长阶段个体重显著高于其它2个基因型,但GG群体日增重呈极显著(P<0.01);在伊拉兔样本群体内,AG型群体样本的体重均值在70日龄阶段高于其它两个,并且日增重高于AA(P<0.05)。而GG型个体在后期3个阶段(56~84日龄)的体重均值高于AA型和AG型,且日增重极显著(P<0.01);在新西兰兔群体内,两组样本群体(AG型和GG型)在84日龄阶段体重均值和日增重均呈显著(P<0.05)。
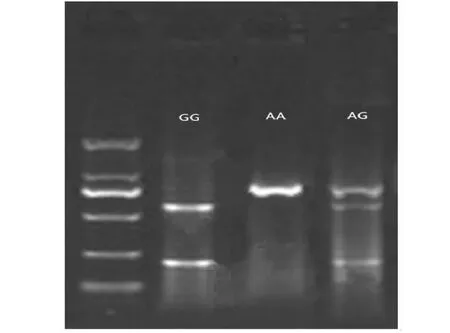
图4 ANGPTL4基因c.823 G>A位点酶切分型结果Fig.4 The typing result of ANGPTL4 gene c.823 G>A loci by RFLP
3 讨 论
3.1 ANGPTL4基因多态性分析
本试验发现ANGPTL4基因的第6个外显子中存在3个变异位点(c.823 G>A,c.867 T>C和c. 975 T>C)均位于ANGPTL4基因所特有的FBG结构域,该结构域位于纤维蛋白原中的β和γ链的C端,其分别由α、β和γ 3条多肽链的N端通过二硫键连接组成[9]。3条链的C端各自形成一个球状区域,在血液凝集过程中凝集下游信号因子的作用,其中β和γ链的C端结构域基本相同,即为FBG结构域[10]。而这3个SNPs均位于FBG结构域,由此推测SNPs对ANGPTL4蛋白结构域的功能具有一定影响,从而对家兔生长发育具有一定影响。而进一步分析发现3个SNPs位点(c.823 G>A、c.867 T>C和c. 975 T>C)高度连锁,由此这3个位点具有相似的遗传效应。因此,本试验最终选择非同义突变c.823 G>A位点作为候选SNP进行后续研究。
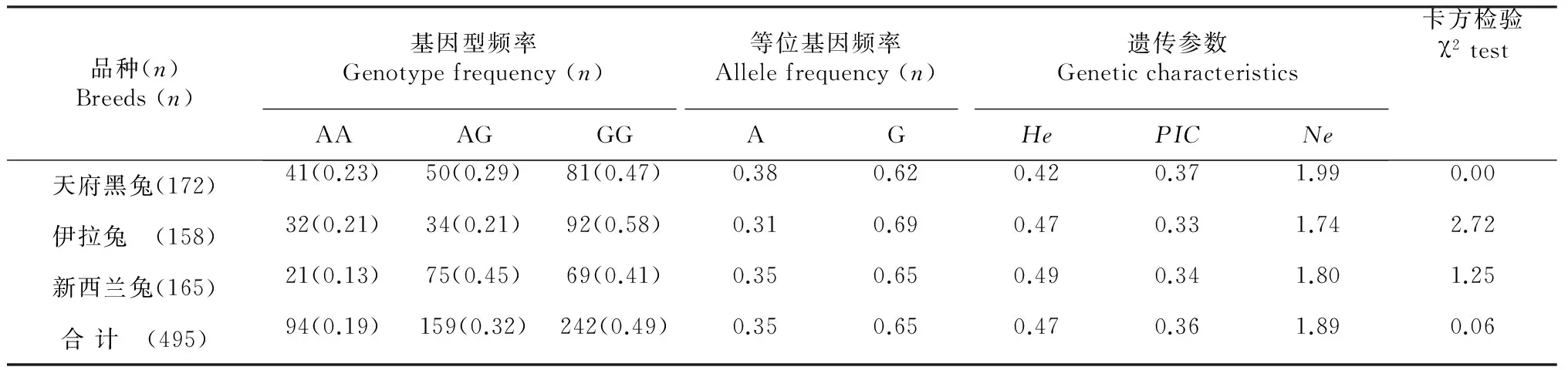
表2 ANGPTL4基因SNP c.823 G>A位点基因频率与基因型频率
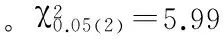
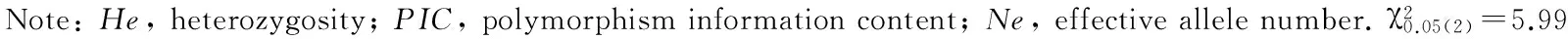
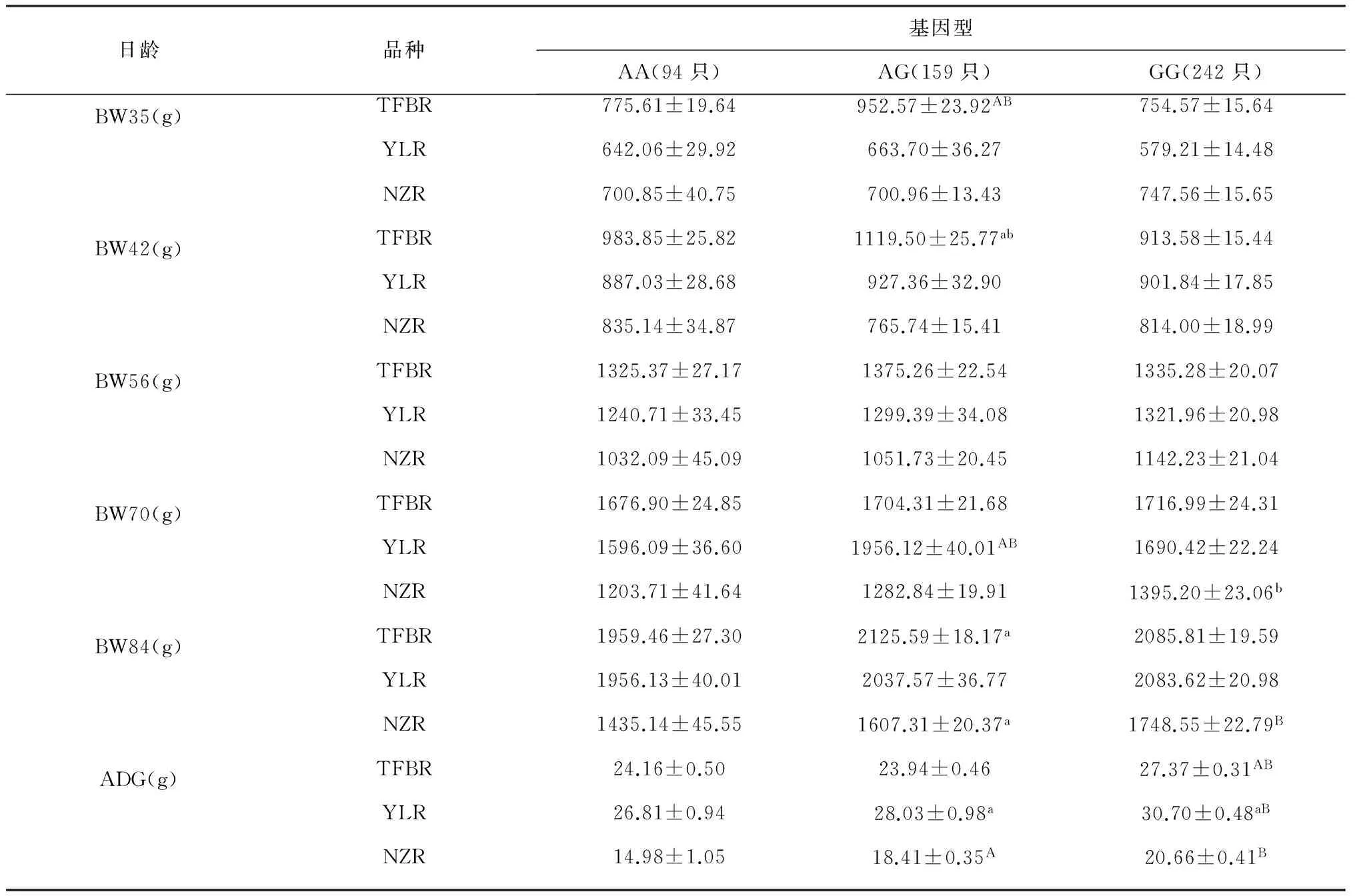
表3 ANGPTL4基因c.823 G>A位点对生长性状的遗传效应分析
注:35 W、42 W、56W、70 W和84 W分别表示35日龄、42日龄、56日龄、70日和84日龄体重;ADG表示35~84日龄平均日增重;小写字母(ab)和大写字母(AB)分别表示差异性显著(P<0.05)和差异极显著(P<0.01);TFBR、YLR和NZR分别表示天府黑兔、伊拉兔和新西兰兔。
Note: 35 W, 42 W, 56, 70 W, and 84 W represent the body weight of 35, 42, 56, 70 and 84 days of age respectively; ADG represents the average daily gain from 28 to 84 days of age. Lower case letters (ab) and capital letters (AB) indicated significant difference (P<0.05) and significant difference (P<0.01); TFBR, YLR and NZR represent the rabbit breed Tianfu black rabbit, IRA rabbit and New Zealand rabbit, respectively.
以参照多态信息含量(polymorphism information content,PIC)指标为依据,对c.823 G>A位点在3个家兔品种内的基因多态性分析表明该位点在各个群体中均表现为中度多态(0.25
3.2 ANGPTL4基因与家兔生长性状相关性分析
ANGPTL4基因在机体的生长代谢过程中具有重要作用,其可以调节血管的生成、脂类物质的代谢以及糖类代谢平衡[12]。侯飞等通过检测多个牛品种(南阳牛、郏县红牛、鲁西牛等)的ANGPTL4基因编码区发现存在4个遗传变异位点,其中,1422 T>C为错义突变,而其它3个均为同义突变。通过关联性分析表明c. 1422 T>C位点多态性与不同牛品种的体重呈正相关[13]。张静等通过检测ANGPTL4基因非编码区在广灵大尾羊和小尾寒羊的遗传变异位点分析发现存在8个SNPs,其中-1691 C>G和-577 T>G 2个颠换的等位基因频率在品种之间存在显著差异。而-1691 C>G和-1625 G>A均以正向超显性遗传方式影响绵羊尾长、宰前体重和胴体重,而以负向超显性效应的方式影响广灵大尾羊的屠宰率[14]。而在本试验中发现家兔群体样本中非同义突变c.823 G>A位点对3个家兔品种的后期增重具有较高的遗传效应,2种基因型(AG和GG)在3个家兔品种中的增重优势明显,且GG基因型所占群体的日增重极显著(P<0.01)。因此,c.823 G>A位点与3个家兔品种的生长发育相关性明显。ANGPTL4基因的c.823 G>A位点使丝氨酸(Ser)→甘氨酸(Gly),从而有可能改变了ANGPTL4蛋白的构象和功能。这Ser和Gly在人体内属于非必需氨基酸,而非必需氨基酸在生物代谢过程中也具有重要影响,其中Gly是一种抑制性较强的神经递质,在信息传导过程中发挥重要作用。当细胞收到刺激后,它通过对神经传导过程中的Na+/K+离子通道的启闭,从而抑制细胞的信号传导。而研究证实Gly与动物细胞的免疫和应激有密切联系,其可能是通过抑制Ca+信号转导、阻止炎症因子被激活、减少氧应激反应等其它协同保护机制[15-16]。因此,在家兔的能量代谢中,ANGPTL4基因c.823 G>A位点有可能直接或间接影响了基因的转录翻译过程以及蛋白质的构象,进一步使蛋白功能增强,从而使家兔在不同的生长阶段的日增重起促进作用[17-18],鉴于本试验所选取的品种和样本量有限,其潜在的生物学效应仍需进一步研究。
4 结 论
ANGPTL4基因的第6外显子上存在3个SNPs,其中c.823 G>A为非同义突变,c.867 T>C和c. 975 T>C均为同义突变,且3个位点完全连锁。c.823 G>A遗传变异分析发现存在A和G等位基因和AA、AG和GG基因型,其中G和GG均具有优势遗传效应。该c.823 G>A位点的关联性分析表明2种基因型(AG和GG)与所选3个家兔群体样本部分生长性状呈显著(P<0.05),且GG基因型的平均日增重呈极显著(P<0.01)。因此,ANGPTL4基因可以作为改良家兔生产性能的候选基因。
[1]Klopper J P, Berenz A, Hays W R, et al. In vivo and microarray analysis of rexinoid-responsive anaplastic thyroid carcinoma[J]. Clinical Cancer Research An Official Journal of the American Association for Cancer Research, 2008, 14(2):589-96.
[2]Mandard S, Zandbergen F, Van S E, et al. The fasting-induced adipose factor/angiopoietin-like protein 4 is physically associated with lipoproteins and governs plasma lipid levels and adiposity[J]. Journal of Biological Chemistry, 2006, 281(2):934-44.
[3]Sukonina V, Lookene A, Olivecrona T, et al. Angiopoietin-like protein 4 converts lipoprotein lipase to inactive monomers and modulates lipase activity in adipose tissue[J]. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103(46):17450-17455.
[4]Legry V, Bokor S, Cottel D, et al. Associations between common genetic polymorphisms in angiopoietin-like proteins 3 and 4 and lipid metabolism and adiposity in European adolescents and adults[J]. Journal of Clinical Endocrinology & Metabolism, 2009, 94(12):5070-5077.
[5]Gealekman O, Burkart A, Chouinard M, et al. Enhanced angiogenesis in obesity and in response to PPARgamma activators through adipocyte VEGF and ANGPTL4 production[J]. Ajp Endocrinology and Metabolism, 2008, 295(5):1056-1064.
[6]Jitrapakdee S, Slawik M, Medina-Gomez G, et al. The peroxisome proliferator-activated receptor-gamma regulates murine pyruvate carboxylase gene expression in vivo and in vitro[J]. Journal of Biological Chemistry, 2005, 280(29):27466-27476.
[7]Belanger A J, Lu H, Date T, et al. Hypoxia up-regulates expression of peroxisome proliferator-activated receptor gamma angiopoietin-related gene (PGAR) in cardiomyocytes: role of hypoxia inducible factor 1alpha[J]. Journal of Molecular and Cellular Cardiology, 2002, 34(7):765-774.
[8]Kim H K, Youn B S, Shin M S, et al. Hypothalamic Angptl4/Fiaf Is a Novel Regulator of Food Intake and Body Weight[J]. Diabetes, 2010, 59(11):2772-2780.
[9]Weisel J W, Stauffacher C V, Bullitt E, et al. A model for fibrinogen: domains and sequence[J]. Science, 1985, 230(4732):1388-1391.
[10]Lu J, Teh C, Kishore U, et al. Collectins and ficolins: sugar pattern recognition molecules of the mammalian innate immune system[J]. Biochimica et Biophysica Acta. General Subjects, 2002, 1572(3):387-400.
[11]Botstein D, White R L, Skolnick M, et al. Construction of a genetic linkage map in man using restriction fragment length polymorphisms[J]. American Journal of Human Genetics, 1980, 32(3):314-331.
[12]Zhu H, Li J, Qin W, et al. [Cloning of a novel gene,ANGPTL4 and the functional study in angiogenesis][J]. National Medical Journal of China, 2002, 82(2):94-99.
[13]侯 飞. 黄牛ANGPTL4、GPIHBP1基因SNPs检测及其与生长性状的关联分析[D]. 西北农林科技大学, 2011.
[14]张 静, 景炅婕, 张 方,等. 绵羊ANGPTL4基因的多态性及其与尾型和屠宰性状的关联研究[J]. 中国畜牧杂志, 2016, 52(5):12-18.
[15]Li X, Bradford B U, Wheeler M D, et al. Dietary glycine prevents peptidoglycan polysaccharide-induced reactive arthritis in the rat: role for glycine-gated chloride channel[J]. Infection and immunity, 2001, 69(9):5883-5891.
[16]Stachlewitz R F, Li X, Smith S, et al. Glycine inhibits growth of T lymphocytes by an IL-2-independent mechanism[J]. Journal of Immunology, 2000, 164(1):176-182.
[17]Camargo G M F D, Porto-Neto L R, Kelly M J, et al. Non-synonymous mutations mapped to chromosome X associated with andrological and growth traits in beef cattle[J]. BMC Genomics, 2015, 16(1):1-10.
[18]Noguchi K, Morishima Y, Takahashi S, et al. Impact of nonsynonymous mutations of factor X on the functions of factor X and anticoagulant activity of edoxaban[J]. Blood Coagulation and Fibrinolysis An International Journal in Haemostasis & Thrombosis, 2015, 26(2).
PolymorphismsofANGPTL4GeneandItsAssociationwithGrowthTraitsinRabbits
LI Yan-hong, HU Shen-qiang, WANG Jie, JIA Xian-bo, LAI Song-jia*
(Institute of Animal Genetics and Breeding, Sichuan Agricultural University, Sichuan Chengdu 611130,China)
【Objective】 The aim of this study was to investigate the association between polymorphisms in the angiopoietin like protein 4 (ANGPTL4) gene and growth traits of domestic rabbits. 【Method】 The polymerase chain reaction (PCR) products were first screened for single nucleotide polymorphisms (SNPs) of rabbitANGPTL4 gene by direct DNA sequencing, and the PCR-restriction fragment length polymorphism (RFLP) assay was further used for genotyping of a total of 459 samples from three meat rabbit breeds (Tianfu Black, IRA and New Zealand rabbits). Thereafter, the association of polymorphisms in this gene with growth traits was evaluated. 【Result】 Our results showed that three SNPs, including a non-synonymous mutation c.823 G>A and two synonymous mutations c.867 T>C, and c. 975 T>C, were detected in exon 6 ofANGPTL4. Owing to a completely linkage relationship (r2≥ 0.75) among these three SNPs, only the c.823 G>A was selected for analysis of genetic effects. The association analysis of its genotypes with growth traits revealed that the genotypes of AG and GG were strongly correlated with some growth traits in three breeds of rabbits. 【Conclusion】 These data suggested thatANPGTL4 could be a candidate gene affecting rabbit growth and the SNP c.823 G>A might be used as a potential genetic marker in rabbit breeding.
ANGPTL4 gene; Polymorphism; Growth traits; Association analysis; Rabbit
1001-4829(2017)10-2382-06
10.16213/j.cnki.scjas.2017.10.037
2016-12-26
国家兔产业技术体系建设(CARS-44-A-2)
李彦宏(1990-),男,硕士研究生,从事家兔遗传育种研究方向,417590007@qq.com,*为通讯作者,E-mail: laisj5794@163.com。
S829.1
A
(责任编辑 李 洁)
