近十年中国重要食药用菌研究进展
2017-11-08董彩虹刘奇正张娇娇
董彩虹 , 刘奇正, 张娇娇, 2
(1.中国科学院微生物研究所 真菌学国家重点实验室, 北京 100101;2.中国科学院大学, 北京 100039)
·大家专版·
董彩虹,博士,中国科学院微生物研究所研究员,博士生导师。2006年博士毕业于中国科学院微生物研究所,2014年晋升为研究员。现任中国菌物学会产业工作委员会副主任,中国菌物学会产业分会副秘书长,贵州省农业科学院、河南省科学院生物研究所特聘研究员,河北省食用菌协会高级顾问,国家中医药管理局人才交流中心虫草鉴定职业规范化培训专家委员会专家,中国虫草产业技术创新战略联盟冬虫夏草专业委员会专家组成员、杭州市钱江特聘专家,中国科学院"科学人讲坛"特邀专家。研究方向为珍稀食药用菌生物学研究,侧重于环境因素对食用菌生长发育和代谢产物产生的调控机理,涉及类群包括具有重要经济价值的珍稀种类冬虫夏草、蛹虫草、绣球菌、羊肚菌、灰树花和香蘑等。2015年以第一作者主笔“我国虫草产业发展现状、问题及展望—虫草产业发展金湖宣言”在《菌物学报》发表并在行业内引起强烈反响。2016年担任《菌物学报》特邀编辑组稿《冬虫夏草专刊》。作为主持人或主要学术骨干先后承担国家自然科学基金项目,国家科技支撑计划、863、973及公益性行业(农业)科研专项子课题,和多家企业建立了合作关系,选育的蛹虫草品种及栽培技术在多家企业生产。发表论文50余篇,包括Critical Reviews in Biotechnology, Applied Microbiology and Biotechnology, LWT-Food Science and Technology, Food and Bioproducts Processing, FEMS Microbiology Letters, Journal of Industrial Microbiology and Biotechnology, Journal of Applied Microbiology等。申请专利8项,其中6项已获得授权,参与编写《中国冬虫夏草》、《食用菌栽培学》。
近十年中国重要食药用菌研究进展
董彩虹1, 刘奇正1, 张娇娇1, 2
(1.中国科学院微生物研究所 真菌学国家重点实验室, 北京 100101;2.中国科学院大学, 北京 100039)
食用菌跻身我国第五大农作物,我国已经成为名副其实的食药用菌大国。本文对近十年来食药用菌重要种类名称变更、人工培植或栽培技术的突破和基因组学研究的进展进行了综述。随着分子系统学研究的深入,灵芝、黑木耳、毛木耳、桑黄等重要种类的名称得到更正和完善;冬虫夏草、羊肚菌、暗褐网柄牛肝菌等难栽培种类在全世界范围内首次实现人工栽培;大多数常见食药用菌种类完成了基因组解析。分析了分类和系统发育研究、生物学研究、组学研究与食药用菌产业发展的关系,基础研究促进食药用菌产业发展,同时产业发展为基础研究提出更多的科学问题。
食药用菌;人工栽培;名称变更;基因组学;产业
我国食药用菌产业发展迅速,2015年年产量3 476.15万吨(鲜品),产值2 516.38亿元,出口食药用菌类产品50.7万吨(干、鲜混计),创汇29.79亿美元。我国食用菌年产量已经多年占世界总产量的70%以上,成为名副其实的食药用菌大国。产业的发展与研究人员的努力密不可分,食药用菌基础科学的研究,面对的不再仅仅是真菌生物学,更多的是产业发展的需求[1]。本文对最近几年来我国食药用菌研究重要种类名称变更、人工培植或栽培技术的突破和基因组学研究等重要进展进行综述,以期促进基础研究和产业的协同发展。
1 食药用菌重要种类名称变更
我国食药用菌物种资源丰富,据戴玉成等[2]统计,中国食用菌共有966个分类单元(936种,23变种,3亚种4变型),并且还在不断增加;中国药用真菌473种[3],2016年出版的《中国大型菌物资源图鉴》[4]共记载大型菌物509属1819 种。过去我国有些食药用菌学名基于形态学研究采用了欧洲或美洲产地的名称,随着分子系统学研究的深入,这些名称正在逐渐得到更正和完善。
1.1灵芝
长期以来一直使用“Ganodermalucidum(Curtis) P. Karst.”作为“灵芝”的学名[5-6]。近几十年来,我国关于药用真菌的主要论著都将灵芝的拉丁名称处理为G.lucidum,如《中国药用真菌》[7]、《中国药用真菌图鉴》[8]、《中国经济真菌》[9]。灵芝相关的产品也基本标注其拉丁名称为G.lucidum。然而,多个独立的分子系统学与形态分类学相结合的研究结果表明,中国广泛栽培的“灵芝”与最初描述于欧洲的“G.lucidum”并不相同[10-11],而是一个新种,并命名为“灵芝GanodermalingzhiSheng H.Wu, Y.Cao & Y.C.Dai”[12]。这个名称应为我国广泛分布和栽培且具有重要药用价值的灵芝的合法科学名称。《中国药典》中记载的赤芝即为灵芝,其拉丁学名应为Ganodermalingzhi[13]。也有观点认为我国现在商业化栽培的物种大多数是四川灵芝GanodermaSichuanenseJ.D. Zhao & X.Q. Zhang[14]。
1.2黑木耳
黑木耳在我国已经有1 000多年栽培历史,多年来我国黑木耳的学名一直使用模式产地为欧洲的Auriculariaauricular-judae,国内外涉及中国黑木耳的主要期刊和论文几乎均使用上述学名[2-3,5,9,15]。近年来,北京林业大学戴玉成团队关于黑木耳分类及系统发育的研究表明,A.auricula-judae实际为一复合种,该复合种在全球范围内有4个种和一个未确定种。我国广泛分布和栽培的黑木耳与欧洲的A.auricular-judae不同,应为一新种,学名为AuriculariaheimuerF. Wu, B.K. Cui & Y.C. Dai[16-17]。
1.3毛木耳
毛木耳,又称黄背木耳,白背木耳,是我国广泛栽培的食用菌种类之一。过去中国文献引用毛木耳的拉丁学名基本是Auriculariapolytricha(Mont.) Sacc.,该种的模式产地为北美洲。Looney 等[18]结合分子系统学研究表明,Auriculariapolytricha正确名称应该为Auricularianigricans(Sw.) Birkebak, Looney & Sánchez-García,并被广泛接受。吴芳[17]研究发现目前中国尚未发现长毛木耳Auricularianigricans的分布,在中国甚至世界范围内广泛分布的毛木耳拉丁学名为AuriculariacorneaEhrenb。近年来成功驯化并推广的玉木耳为毛木耳的白色变种[19]。
1.4桑黄
桑黄作为著名传统药用真菌在中国已经有2 000多年的历史,但真正的桑黄种类及学名长久以来并没有定论。《中国的真菌》[15]定名为针裂蹄,拉丁学名Phellinuslinteus(Berk.& M. A. Curtis) Teng,《中国药用真菌》[7]命名为Phellinusigniarius(L.) Quél.,中文名称为“火木层孔菌”,《中国药用真菌图鉴》[8]称为裂蹄针层孔菌。《中国真菌志锈革孔菌科》中记载,目前国内可以称为桑黄的有两种:P.igniarius和P.baumii[20]。日、韩学者几十年来一直采用P.linteus作为桑黄的学名[21]。
2012年台湾真菌学家吴声华团队基于形态特征和ITS序列分析,将桑黄处理为纤孔菌属(Inonotus),并作为新种InonotussanghuangSheng H. Wu, T. Hatt. & Y.C. Dai[22]发表。Zhou 等[23]基于ITS rDNA和nLSU rDNA 序列分析结果及形态特征,将桑黄类群重新进行归类,建立桑黄孔菌新属并命名为SanghuangporusSheng H. Wu, L.W. Zhou &Y.C. Dai,模式种为Sanghuangporussanghuang(Sheng H. Wu, T. Hatt. & Y.C. Dai) Sheng H. Wu, L.W. Zhou & Y.C. Dai。因此,桑黄的学名也从发表新种时的Inonotussanghuang改为Sanghuangporussanghuang[21]。
1.5冬虫夏草
冬虫夏草菌于1843年由英国真菌学家Berkeley定名为SphaeriasinensisBerk.[24];1878年,意大利学者Saccardo又将其转到Cordyceps(Fr.) Link,名称为Cordycepssinensis(Berk.) Sacc.[25];2007年,Sung等基于多基因系统学分析的结果将广义的虫草属CordycepsFr. sensu lato分归为3个不同的科,冬虫夏草菌归到线虫草科(Ophiocordycipitaceae G.H. Sung, J.M. Sung, Hywel-Jones & Spatafora)线虫草属(OphiocordycepsPetch),科和属都被重新修订,名称为Ophiocordycepssinensis(Berk.) G. H. Sung, J. M. Sung, Hywel-Jones & Spatafora[26],O.sinensis的名称已被广泛接受。
除此之外,还有一些种类的名称发生了变更,如猪肚菇,曾被错误鉴定为大杯伞Clitocybemaxima[27],其学名应为巨大革耳[Panusgiganteus(Berk.) Corner≡LentinusgiganteusBerk.][28],2011年转移至Pleurotusgiganteus(Berk.) Karun. & K.D. Hyde[29]。牛樟芝现名为Taiwanofunguscamphoratus(M. Zang & C.H. Su) Sheng H. Wu, Z.H. Yu, Y.C.Dai & C.H. Su,我国工厂化栽培的绣球菌为广叶绣球菌SparassislatifoliaY.C. Dai & Zheng Wang[30]等。正确认识物种是研究和开发的基础,因此,我国食药用真菌分类和系统发育研究还有待于进一步深入展开。
2 食药用菌重要种类人工培植或栽培技术的突破
2.1冬虫夏草
人工培植是解决冬虫夏草自然资源有限而市场需求巨大的矛盾的途径,一直是科学研究和产业发展的热点问题。广东东阳光药业经过十余年研发,突破了冬虫夏草菌培养技术、寄主昆虫虫种选择和规模化繁殖、侵染及大规模成草等关键技术,成功实现了冬虫夏草培植产业化(图1A)[31]。
《菌物学报》2016年设冬虫夏草专刊,介绍了冬虫夏草培植分生孢子高产菌株筛选[32]、培养基和培养条件优化[33]、寄主昆虫品系选育[34]、土壤和植物对寄主昆虫规模化饲养的影响[35]及侵染途径[36]等方面的研究。魏江春等[37]对培植的冬虫夏草通过分子系统学分析进行了物种鉴定,证实了冬虫夏草培植的真实性。培植和野生冬虫夏草主要化学成分和蛋白图谱比较分析显示两者没有显著差异[38-39]。冬虫夏草不同发育时期蛋白质组iTRAQ质谱分析证明了昆虫寄主蛋白逐渐被真菌蛋白替代的变化过程[40]。冬虫夏草人工培植的成功,不论对科学还是产业,都不愧为里程碑式的创新性成果。此外,最近也有报道冬虫夏草野生抚育获得阶段性成果(http://news.sina.com.cn/c/2017-05-17/doc-ifyfekhi7958337.shtml)。
2.2羊肚菌
1982年Ower等[41]报道在室内获得了羊肚菌子实体,随后Ower及其团队在1985至1988年之间先后申请了3个羊肚菌人工栽培的美国专利(US Patents 4594809、4757640、4866878)并获得授权[42-44]。这些专利技术在美国进行了羊肚菌室内商业化栽培,但是因品种退化和细菌污染2008年全面停产[45],至今未恢复生产。

图1 近年来我国成功实现人工栽培的重要食药用菌Fig.1 Species cultivated successfully in China recentlyA:人工栽培冬虫夏草(广东东阳光药业有限公司提供); B:羊肚菌的大田栽培; C:工厂化栽培暗褐网柄牛肝菌(张春霞提供); D:人工栽培牛樟芝A: Ophiocordyceps sinensis; B: Morchella spp.; C: Phlebopus portentosus; D: Taiwanofungus camphoratus
我国从上世纪八十年代开始,多家单位进行了羊肚菌的人工栽培研究,也有报道室外人工栽培成功,但均不能稳产甚至很难重复。直到2000年,外营养添加技术(营养袋)的发明对室外羊肚菌人工栽培产生了积极的推动作用(图1B)[46]。我国羊肚菌人工栽培面积从2002年3 000亩发展到2016年突破2万亩,全国大部分省份都有栽培,栽培的主要种类为梯棱羊肚菌MorchellaimportunaM. Kuo, O′Donnell & T. J. Volk、六妹羊肚菌M.sextelataM. Kuo及少量七妹羊肚菌M.eximiaBoud.[45]。但是目前为止,还是存在产量不稳定的问题。
室外栽培受气候条件、自然灾害等影响较大,目前多家单位对羊肚菌的室内栽培进行了探索,但是目前为止还没有规模化室内栽培的报道。
2.3暗褐网柄牛肝菌
牛肝菌目中大多数科属的种类都属于外生菌根菌,需要用合成菌根苗的方法进行人工栽培。暗褐网柄牛肝菌(Phlebopusportentosus)隶属于牛肝菌目(Boletales)、小牛肝菌科(Boletinellaceae)、网柄牛肝菌属(Phlebopus),国内主要分布在云南和四川,广西和海南也有分布。云南省热带作物科学研究所经过多年的研究,在温室和菇房内用栽培腐生菌的方法成功培养出暗褐网柄牛肝菌子实体(图1C)[47-48],因此认为其可能是一种腐生菌[49]。云南景洪宏臻农业科技有限公司于2015年4月成功进行工厂化生产,日产鲜牛肝菌达到500公斤左右(http://travel.sina.com.cn/china/2015-04-10/2203303666.shtml)。云南省热带作物科学研究所在凤凰木和柚子树下野外仿生栽培也取得了突破[50-51]。
2.4牛樟芝
牛樟芝又名樟芝、樟菇、牛樟菇、红樟芝,气味芳香,味辛、苦,生长环境较苛刻,腐生于台湾特有树种牛樟树树干的中空部或枯死倒伏牛樟树阴暗潮湿的树干表面。由于其寄主的专一性,生长缓慢以及不易人工栽培等因素,牛樟芝极其珍稀昂贵,被誉为“森林红宝石”。
野生资源枯竭加之其良好的保健功能,牛樟芝价格高涨,推动了人工培育技术的发展。目前其人工培养方法主要有4种,分别为椴木栽培法、固体培养法、液体发酵法、皿培式培养法,椴木栽培法近年来取得了突破(图1D)。以原有宿主牛樟树椴木为培养基栽培牛樟芝,培养成本高。近年来,也发展了以苹果木、杉木、福建本地的小叶红心樟椴木等为基质的椴木栽培方式[52-53]。
3 食药用菌基因组研究进展
3.1已完成测序的食药用菌基因组
第一个真核生物酿酒酵母Saccharomycescerevisiae的基因组测序于1996年完成[54],引发了真核生物基因功能和表达的全球性研究。第一个食药用菌玉米黑粉菌(Ustilagomaydis)全基因组测序数据释放于2003年,基因组大小为20.5 Mb,共编码6 902个蛋白[55]。截至2017年6月,在线基因组数据库 NCBI Genome 已经公开的真菌基因组有1 254个,包括食药用菌中灵芝属、虫草属、银耳属、羊肚菌属等多个种类(表1)。
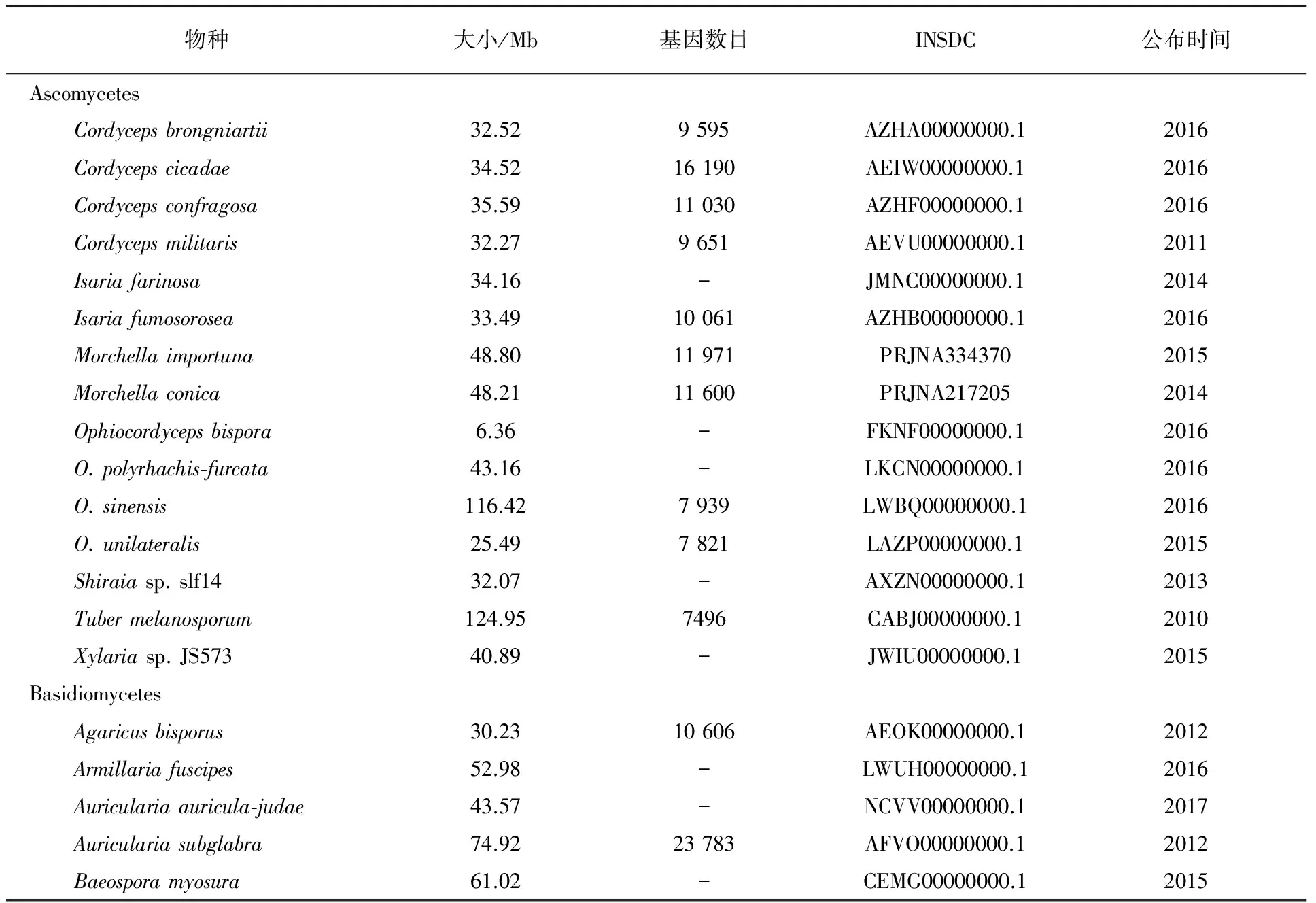
表1 部分已完成和发表的食药用菌基因组数据信息

续表1

续表1
注:INSDC为国际核苷序列联合数据库(International Nucleotide Sequence Database Collaboration)中代码;“-”:NCBI(https://www.ncbi.nlm.nih.gov/)数据库中没有查询到相关数据
3.2食药用菌基因组解析
重要食药用菌的基因组测序已经基本完成。灵芝基因组注释结果显示其具有作为次级代谢产生和调控研究模型生物的潜力,对其多样化木质素降解酶解析将促进其工业化应用[56]。蛹虫草基因组解析证实其基因组不编码对人类有害的已知真菌毒素,且其有性生殖为异宗配合[57],为蛹虫草人工栽培和产品开发提供了有力的支撑。冬虫夏草基因组解析显示其以同宗配合方式进行有性生殖,有助于在青藏高原高海拔环境中快速、低成本地繁衍后代,这可能也是造成冬虫夏草有效群体变小的原因[58-59]。香菇基因组解析揭示了香菇降解木质纤维素的遗传基础,对提高香菇栽培基质的利用效率和开发新型栽培基质具有重要的参考价值[60]。吴冰等[61]比较分析了46个食药用菌和3个降解木质纤维素模式真菌的基因组,发现降解木质纤维素相关酶系家族的平均数量:草腐真菌>白腐真菌>褐腐真菌>共生营养真菌;并且这个规律在纤维素、半纤维素、木质素、果胶、几丁质降解酶中都适用,这可能与不同生态类型的食药用真菌在生态环境中的作用相关。可见基因组解析将促进食药用菌遗传育种、基质利用、人工栽培和产品开发。
4 我国食药用菌基础研究与产业发展的思考
4.1食药用菌分类、系统发育研究与产业发展
分类地位和生物学种的鉴定是食用菌野生种质资源评价的第一步,物种的准确鉴定是有效利用的重要基础,科学的物种名称是化学、药理学研究的保障,也是产业壮大、国际化的基础。最典型的例子是虫草属真菌及其相关产业。年总产值超过300亿的虫草及其产品,在我国已经形成了一个巨大的产业,虫草产业快速发展的同时也带来了诸多乱象,这些乱象的形成和物种名称、分类地位的混乱息息相关[62]。最近的研究表明我国广泛栽培的灵芝不是欧洲的G.lucidum,中国的桑黄不是美洲的I.linteus,木耳不是欧洲的A.auricula-judae。总之,面对我国如此丰富的资源宝库,食用菌产业从业人员应该和从事分类、系统发育研究的专业人员紧密合作,共担责任,共享成果,保障产业的健康发展。
4.2食药用菌生物学研究与产业发展
冬虫夏草、羊肚菌和暗褐网柄牛肝菌人工培植和驯化的成功都与其生物学研究的进展密不可分。冬虫夏草培植的成功建立在冬虫夏草菌、蝙蝠蛾幼虫及虫菌关系等生物学研究的基础上,其人工培植技术的成功再次证实了生物学研究给产业发展带来了重要的影响。另一方面,现有的科学研究不能满足产业发展的需求。如羊肚菌虽然人工栽培成功,但是生活史不明,菌核和菌霜在生活史中的作用、营养袋的作用、原基分化和发育的条件等诸多生物学问题均未研究清楚,这些生物学问题也是目前栽培不能稳产的重要因素[45]。由此可见,生物学研究是食药用菌驯化栽培技术突破的关键,同时产业发展为生物学研究提出了更多的科学问题。
4.3食药用菌组学研究与产业发展
食药用菌中半数以上物种的基因组数据是在近3年内获得的(表1),食药用菌正在迎来基因组测序的高潮,更多有价值物种的测序数据将共享。除了基因组解析外,转录组、蛋白组、线粒体组等研究也在不断深入。因为技术问题,在过去很长一段时间对食药用菌进行遗传操作比较困难,随着组学技术的不断完善和新技术的应用,将为食药用菌的基础研究提供有力的工具。2016年轰动食药用菌学术界和产业界的双孢蘑菇基因编辑以达到抗褐变的目的,美国农业部宣布将不会对这种利用CRISPR-Cas9基因组编辑进行遗传修饰的蘑菇进行监管。随着组学研究的深入,特别是组学研究直接结合产业发展,注重解决产业中遇到的实际问题,将有力促进产业发展。
食药用菌以其营养保健的双重功效,肩负着增进国民健康的重任,已成为大健康产业不可或缺的部分;食药用菌产业有助于调整种植业结构,促进农民增收,助力精准扶贫和解决“三农问题”;食药用菌有效利用农林废弃物,是农业循环经济的主力军,是现代生态农业产业化的重要组成部分。总之,食药用菌产业遇到了极好的发展机遇,食药用菌研究人员势必抓住机遇,扎实研究,助力产业发展。
致谢北京林业大学戴玉成教授对本文进行了审阅并提出了修改建议,云南省热带作物科学研究所张春霞研究员提供暗褐网柄牛肝菌工厂化栽培照片并审阅相关内容,广东东阳光药业有限公司提供冬虫夏草人工栽培图片,在此致以衷心的感谢!
[1]张金霞, 陈强, 黄晨阳, 等. 食用菌产业发展历史、现状与趋势[J]. 菌物学报, 2015, 34(4): 524-540.
[2] 戴玉成, 周丽伟, 杨祝良, 等.中国食用菌名录[J]. 菌物学报, 2010, 29: 1-21.
[3] 戴玉成, 杨祝良. 中国药用真菌名录及部分名称的修订[J]. 菌物学报, 2008, 27: 801-824.
[4] 李玉, 李泰辉, 杨祝良, 等. 中国大型菌物资源图鉴[M]. 郑州: 中原农民出版社, 2015.
[5] 戴芳澜. 中国真菌总汇[M]. 北京: 科学出版社, 1979: 1-1527.
[6] 赵继鼎, 张小青. 中国真菌志 第十八卷 灵芝科[M]. 北京: 科学出版社, 2000: 1-204.
[7] 刘波. 中国药用真菌[M]. 太原: 山西人民出版社, 1984: 1-228.
[8] 应建浙, 卯晓岚, 马启明. 中国药用真菌图鉴[M]. 北京: 科学出版社, 1987: 1-579.
[9] 卯晓岚. 中国经济真菌[M]. 北京: 科学出版社, 1998: 1-762.
[10] Smith BJ, Sivasithamparam K. Internal transcribed spacer ribosomal DNA sequence of five species ofGanodermafrom Australia [J]. Mycological Research, 2000, 104: 943-951.
[11] Hong SG, Jung HS. Phylogenetic analysis ofGanodermabased on nearly complete mitochondrial small-subunit ribosomal DNA sequences [J]. Mycologia, 2004, 96: 742-755.
[12] Cao Y, Wu SH, Dai YC. Species clarification of the prize medicinalGanodermamushroom"Lingzhi"[J]. Fungal Diversity, 2012, 56: 49-62.
[13] 戴玉成, 曹云, 周丽伟, 等. 中国灵芝学名之管见[J]. 菌物学报, 2013, 32(6): 947-952.
[14] Wang XC, Xi RJ, Li Y, et al. The species identity of the widely cultivated Ganoderma,G.lucidum′ (Ling-zhi), in China [J]. PLoS ONE, 2012, 7(7): e40857.
[15] 邓叔群. 中国的真菌[M]. 北京: 科学出版社, 1963: 1-808.
[16] Wu F, Yuan Y, Malysheva VF, et al. Species clarification of the most important and cultivatedAuriculariamushroom “Heimuer”: evidence from morphological and molecular data [J]. Phytotaxa, 2014, 186(5): 241-253.
[17] 吴芳. 木耳属的分类与系统发育研究[D]. 北京:北京林业大学, 2016: 1-107.
[18] Looney BP, Birkebak JM, Matheny PB. Systematics of the genusAuriculariawith an emphasis on species from the southeastern United States [J]. North American Fungi, 2013, 8(6): 1-25.
[19] 李晓, 张阔谭. 木耳属白色变异菌株的研究进展[J].食药用菌, 2016,24(4): 230-233, 238.
[20] 张小青, 戴玉成. 中国真菌志-锈革孔菌科[M].北京: 科学出版社, 2005: 117-191.
[21] 吴声华, 黄冠中, 陈愉萍, 等. 桑黄的分类及开发前景[J]. 菌物研究, 2016, 14(4): 187-200.
[22] Wu SH, Dai YC, Hattori T, et al. Species clarification for the medicinally valuable "sanghuang" mushroom [J]. Botanical Studies, 2012, 53: 135-149.
[23] Zhou LW, Vlasák J, Decock C, et al. Global diversity and taxonomy of the Inonotus linteus complex (Hymenochaetales, Basidiomycota):Sanghuangporusgen. nov.,TropicoporusexcentrodendriandT.guanacastensisgen. et spp. Nov., and 17 new combinations[J]. Fungal Diversity, 2016, 1(77): 335-347.
[24] Berkeley MJ. On some entomogenousSphaeriae[J]. London Journal of Botany, 1843, 2: 205-211.
[25] Saccardo PA. Enumeratio pyrenomycetum hypocreaceorum hucusque congitorum systemate carpologico dispositorum [J]. Michelia, 1878, 1: 277-325.
[26] Sung GH, Hywel-Jones NL, Sung JM, et al. Phylogenetic classification ofCordycepsand the clavicipitaceous fungi [J]. Studies in Mycology, 2007, 57: 5-59.
[27] 王波. 几种食用菌品种名称订正[J]. 中国食用菌, 2006, 25(5): 27-28.
[28] 邓旺秋, 李泰辉, 陈枝南, 等. 栽培食用菌猪肚菇的学名考证[J]. 食用菌学报, 2006, 13(3): 71- 74.
[29] Karunarathna SC, Yang ZL, Ko TWK, et al.Lentinusgiganteusrevisited: new collections from Sri Lanka and Thailand [J]. Mycotaxon, 2011, 118: 57-71.
[30] 廉添添, 杨涛, 孙军德, 等. 人工栽培绣球菌的鉴定及其子实体β-葡聚糖含量的酶法测定[J]. 菌物学报, 2014, 33(2): 254-261.
[31] 李文佳, 董彩虹, 刘杏忠, 等. 冬虫夏草培植技术研究进展[J]. 菌物学报, 2016, 35(4): 375-387.
[32] 吕延华, 梁关海, 朱志钢, 等. 冬虫夏草不同部位来源的菌株及多次传代固体发酵特性[J]. 菌物学报, 2016, 35(4): 433-439.
[33] 张宗耀, 梁关海, 梁蕾, 等. 培养基及培养条件对冬虫夏草菌固体发酵产分生孢子的影响[J]. 菌物学报, 2016, 35(4): 440-449.
[34] 李全平, 贺媛, 刘杰明, 等. 冬虫夏草寄主昆虫选育及生殖退化研究[J]. 菌物学报, 2016, 35(4): 456-466.
[35] 李文佳, 李全平, 魏再鸿, 等. 土壤和植物对冬虫夏草寄主昆虫规模化饲养的影响[J]. 菌物学报, 2016, 35(4): 467-475.
[36] 李文佳, 韦瑞升, 夏金明, 等. 蝙蝠蛾幼虫肠道中的冬虫夏草菌菌体研究[J]. 菌物学报, 2016, 35(4): 450-455.
[37] 魏江春, 魏鑫丽, 郑维发, 等. 现代工业化培植的冬虫夏草物种鉴定与成分检测[J]. 菌物学报, 2016, 35(4): 404-410.
[38] 钱正明, 李春红, 李文庆, 等. 冬虫夏草蛋白图谱及干燥条件对超氧化物歧化酶活性影响[J]. 菌物学报, 2016a, 35(4): 424-432.
[39] 钱正明, 李文庆, 孙敏甜, 等. 冬虫夏草化学成分分析[J]. 菌物学报, 2016b, 35(4): 476-490.
[40] 张晗星, 钱正明, 苏尧, 等. 冬虫夏草不同发育时期蛋白质组iTRAQ 质谱分析[J]. 菌物学报, 2016, 35 (4): 411-423.
[41] Ower RD. Notes on the development of the morel ascocarp [J]. Mycologia, 1982,74:142-144.
[42] Ower RD, Mills GL, Malachowski JA. Cultivation ofMorchella[P]. US, US4594809, 1986.
[43] Ower RD, Mills GL, Malachowski JA. For culturing ascocarps of species of the genusMorchella[P]. US, US4757640, 1988.
[44] Ower RD, Mills GL, Malachowski JA. Cultivation ofMorchella[M]. US, US4866878, 1989.
[45] Liu QZ, Ma HS, Zhang Y, et al. Artificial cultivation of true morels: current state, issues and perspectives[J]. Critical Reviews in Biotechnology, 2017. doi: 10.1080/07388551.2017.1333082.
[46] 谭方河. 羊肚菌人工栽培技术的历史、现状及前景[J]. 食药用菌, 2016(3): 140-144.
[47] 纪开萍, 张春霞, 曾雁, 等. 盆栽条件下暗褐网柄牛肝菌人工菌塘及其子实体的培养 [J]. 植物分类与资源学报, 2007, 29(5): 554-558.
[48] 曹旸, 纪开萍, 刘静, 等. 瓶栽条件下覆土方法对暗褐网柄牛肝菌子实体生长的影响[J].食用菌学报, 2010, 17(3): 29-32.
[49] 张春霞, 何明霞, 曹旸, 等. 暗褐网柄牛肝菌是外生菌根菌吗?--暗褐网柄牛肝菌与思茅松和栲树的菌根合成[J]. 食用菌学报, 2016, 23(1) :37-41.
[50] 刘静, 曹旸, 张春霞, 等. 柚子树下暗褐网柄牛肝菌的仿生栽培研究[J]. 中国热带农业, 2013, 51: 53-55.
[51] 刘静, 何明霞, 王文兵, 等. 暗褐网柄牛肝菌仿生栽培研究[J]. 西南农业学报, 2017, 30(5): 1176-1182.
[52] 程显好, 冯路瑶, 刘静, 等. 一种采用苹果木栽培牛樟芝子实体的方法及其培养基[M]. 中国, CN201610238278.1, 2016.
[53] 黄阿贤. 牛樟芝的椴木栽培法[M]. 中国, CN201310017165.5, 2013.
[54] Goffeau A, Barrell BG, Bussey H, et al. Life with 6000 genes [J]. Science, 1996, 274: 546-567.
[56] Chen L, Gong Y, Cai Y, et al. Genome sequence of the edible cultivated mushroomLentinulaedodes(Shiitake) reveals insights into lignocellulose degradation [J]. PLoS ONE, 2016, 11(8): 1-20.
[57] Zheng P, Xia Y, Xiao G, et al. Genome sequence of the insect pathogenic fungusCordycepsmilitaris, a valued traditional Chinese medicine. Genome Biol, 2011, 12: R116.
[58] Hu X, Zhang YJ, Xiao GH, et al. Genome survey uncovers the secrets of sex and lifestyle in caterpillar fungus [J]. Chin Sci Bull, 2013, 58: 2846-2854.
[59] Xia EH, Yang DR, Jiang JJ, et al. The caterpillar fungus,Ophiocordycepssinensis, genome provides insights into highland adaptation of fungal pathogenicity [J]. Scientific Reports, 2017, 7(1): 1806.
[60] Chen S, Xu J, Liu C, et al. Genome sequence of the model medicinal mushroomGanodermalucidum[J]. Nature Communications, 2012, 3: 1-9.
[61] 吴冰, 章小灵, 崔宝凯, 等. 食(药)用真菌比较基因组分析揭示其生态特性[J]. 菌物学报, 2015, 34(4): 742-760.
[62] 董彩虹, 李文佳, 李增智, 等. 我国虫草产业发展现状、问题及展望—虫草产业发展金湖宣言[J]. 菌物学报, 2016, 35 (1): 1-15.
ResearchprogressonimportantedibleandmedicinalfungiinChinaoverthelastdecade
DONG Cai-hong1, LIU Qi-zheng1, ZHANG Jiao-jiao1,2
(1.StateKeyLaboratoryofMycology,InstituteofMicrobiology,ChineseAcademyofSciences,Beijing100101;2.UniversityofChineseAcademyofSciences,Beijing100039)
Edible fungi are now the fifth largest crops in China and the annual output in China has accounted for more than 70% of the total of the world. Here, we review the important progress including name revision, breakthrough in artificial cultivation and genomics of edible and medicinal fungi. The names of the important species are corrected asGanodermalingzhi,Auriculariaheimuer,Auriculariacornea,SanghuangporussanghuangandOphiocordycepssinensis, respectively. Some species have been cultivated successfully for the first time throughout the world includingOphiocordycepssinensis,Morchellaspp.,PhlebopusportentosusandTaiwanofunguscamphoratus. Genome sequencing and annotation have been completed for most of the common species. The fundamental research and industry development promote each other.
edible and medicinal fungi; artificial cultivation; name revision; genomics; industry
国家自然科学基金项目(31572179);山西省煤基重点科技攻关项目(FT2014-03-01);广西重点研发计划项目(2016AB05317)
董彩虹 女,博士,研究员,博士生导师。主要从事食药用菌生物学研究。Tel:010-64806138,E-mail:dongch@im.ac.cn
2017-07-08
Q93
A
1005-7021(2017)04-0001-09
10.3969/j.issn.1005-7021.2017.04.001
