蛋白质组学方法评估转基因抗虫玉米非预期效应
2017-11-07郝文媛李飞武闫伟李葱葱郝东云郭长虹
郝文媛,李飞武,闫伟,李葱葱,郝东云,郭长虹
蛋白质组学方法评估转基因抗虫玉米非预期效应
郝文媛1,2,李飞武2,闫伟2,李葱葱2,郝东云1,2,郭长虹1
(1哈尔滨师范大学生命科学与技术学院,哈尔滨 150025;2吉林省农业科学院,长春130033)
【】转基因育种是将对人类有益的外源基因通过生物技术整合进受体植物的基因组中,使其获得通过自然进化无法获得优异性状,这是转入外源基因的“预期效应”。然而,外源基因插入受体植物还可能产生无法控制和预期的细胞、代谢或表型等方面的改变,即“非预期效应”。非预期效应是转基因植物安全评价的核心内容之一,也是近年来研究的热点和难点。本研究以抗虫转基因玉米为材料,利用差异蛋白质组学技术比较研究转基因玉米与其非转基因玉米之间的非预期效应,为蛋白质组学技术解析转基因植物非预期效应提供理论参考。【】选取4份已经进入中国农业部安全评价阶段转苏云芽孢杆菌(,Bt)内毒素基因的抗虫玉米材料,即SK12-5zd、IE034z、Bt799z、Bt799zd,以及它们的对照玉米材料郑单958和郑58,分别种植于可控人工温室中。待玉米幼苗生长至5叶1心期时,每份材料取长势一致的6个单株最上部完全展开叶片,混合作为一个样本进行取样,并提取叶片蛋白质。双向电泳技术分别分离4种转基因玉米和它们各自对应的非转基因材料高丰度蛋白质组,利用Image Scanner扫描仪扫描脱色后的蛋白质凝胶,PD Quest 8.0软件(Biorad,USA)分析蛋白质凝胶图谱,以蛋白点相对体积(% Vol)来表述每一个匹配蛋白质的表达丰度,将电泳凝胶上特异性蛋白质和相对体积大于2倍的差异蛋白质点取样,依据相应蛋白质数据库进行质谱鉴定,并对鉴定出的特异性和差异性蛋白质进行细胞功能富集分析(GO)和代谢途径富集分析(KEGG),以评价转基因抗虫玉米的可能非预期效应。【】通过比较4种不同转基因抗虫玉米及其非转基因对照的高丰度蛋白质组,除目标抗虫基因外,共鉴定出61个蛋白质,其功能主要是富集于光合作用、碳固定、能量转运等基础细胞功能相关酶类,如核酮糖二磷酸羧化酶、ATP合成酶、丙酮酸磷酸双激酶等。仅有少数几个与光合作用、碳固定和ATP合成途径等基础代谢过程相关基因出现上调表达。KEGG分析表明,与对照ZD958相比,SK12-5zd的差异蛋白在光合生物碳固定途径中显著富集;Bt799zd的差异蛋白则分别在光合生物碳固定途径、光合作用和碳代谢途径中显著富集。与对照Z58相比,IE034z和Bt799z的差异蛋白均在光合作用代谢途径中显著富集。证明4种转基因材料与对照蛋白质组相似性较高,未见基因的异常表达。【】测试的4种转基因材料在蛋白质水平未发生影响其安全性的非预期效应;当前结果表明蛋白质组学技术可以用来验证转基因植物的非预期效应。
转基因玉米;蛋白质组学;非预期效应;双向电泳;差异表达蛋白
0 引言
【研究意义】植物转基因技术的目标是将有利于植物自身或满足人类需求的特定外源基因整合进入受体植物基因组中,使其获得外源基因所表现的特定性状并能够稳定遗传。这些性状就是转基因植物的目标性状,也称为预期效应。由于外源基因整合位点不确定,加之外源基因产物本身也可能使受体植物的能量代谢和物质代谢发生变化,从而影响其固有的生理代谢过程,乃至改变受体植物的固有性状。这些性状改变均是转基因育种设计过程中无法控制和预期的,这些无法控制和预期的性状改变均称为非预期效应[1-2]。可见,转基因植物实际上是外源基因发挥预期的目标效应和受体植物基因组因外源基因的导入,而产生的非预期效应的综合体。转基因植物是否会产生非预期效应,以及对非预期效应的安全性评估一直是科学家和公众对转基因植物广泛关注的话题。长期以来,各国相关部门都是基于1993年欧洲经济发展与合作组织OECD(Organization for Economic Co-operation and Development)提出的“实质等同性原则”对转基因植物进行安全性评价的,评价的核心是比较转基因植物与其非转基因受体植物间在农艺性状、环境适应性、营养及物质组成变化等的差异,以确认或排除转基因植物是否存在非预期效应。传统的检测方法不仅难以全面涵盖转基因植物可能出现的非预期效应,且检测指标有限、检测费用高、费时、费力。因此,建立全面、快速、准确、灵敏的检测技术是分析鉴定转基因生物安全性的关键所在[2-3]。【前人研究进展】近年来,蛋白质组分析技术迅速发展,双向电泳技术、iTRAQ技术等已成为当今蛋白质组学研究的核心技术[4]。研究人员已经在转基因玉米、水稻、大豆等材料中,对蛋白质组水平上的非预期效应的进行了评估,取得了较好的效果[5-11]。转基因玉米商业化生产以来,已占全球转基因作物种植总面积的30%,居转基因作物的第二位,涉及的农艺性状主要为玉米螟抗性,转入的基因为种类繁多的来自苏云芽孢杆菌(,Bt)的昆虫内毒素基因[12-15](本文统称“Bt基因”)。Coll等[5]利用2D凝胶电泳、质谱技术分析比较,发现在同一地块种植的2种转Bt基因玉米MON810与其对照株系的籽粒蛋白质组几乎一致,只有几个蛋白质点有1—1.8倍的变化,结果表明,转基因玉米MON810与其对照非转基因材料具有实质等同性;Albo等[6]也发现MON810与其非转基因对照的籽粒蛋白质组之间非预期差异基本等同;Zolla等[7]发现同一田间生长的MON810及其对照同一品系的不同世代材料的蛋白质组存在显著差异,且在蛋白质组学水平上,环境因素对非预期变异效应的贡献比转基因插入更大。上述研究结果表明,蛋白质组学技术已逐渐成为检测转基因作物非预期效应的有效方法[16]。【本研究切入点】不同转Bt基因的抗虫玉米材料不但在目标基因上明显不同,而且由于受体材料差异和回交转育过程的不同,造成了最终的抗虫转基因玉米材料间存在着巨大的差异。这些遗传背景上的差异是否会产生非预期效应仍研究甚少,目前的研究还主要集中在Bt基因的抗虫目标预期效应上[17-22]。转基因抗虫玉米SK12-5、IE034、Bt799z和 Bt799zd是中国自主研发的具有潜在产业化前景的转基因新材料,围绕上述新材料已开展了大量育种研发工作及所获相应育种材料的环境安全[23-25]和食用安全评价研究,但是,对这些材料的非预期效应研究未见报道。【拟解决的关键问题】利用双向电泳技术对上述转基因抗虫玉米的4种不同遗传背景材料的转基因材料进行了比较分析,一方面在蛋白质组水平比较不同转基因材料的差异;另一方面从组学的水平初步评估了实验材料蛋白水平上是否发生了影响转基因材料安全性的非预期效应,为非预期效应评价提供数据支撑。
1 材料与方法
1.1 材料
供试材料:编号1#为具有转并有ZD958遗传背景的杂交种SK12-5zd,由浙江大学提供。编号2#为转并有Z58遗传背景的自交系IE034z,由中国农业科学院提供。编号3#为转并有Z58遗传背景的自交系Bt799z,编号4#为转并有ZD958遗传背景的杂交种Bt799zd,均由中国农业大学提供。选取非转基因的自交系Z58和杂交种ZD958作为对照材料,编号分别为5#和6#。编号2#和3#的转基因自交系是目标遗传转化体经2代自交后,再与Z58材料进行6代回交转育后获得了具有纯合、稳定遗传背景的转基因自交系。上述转基因材料均已采用国家发布的转基因检测标准进行了外源基因分子特征鉴定,证明均为独立转化事件,并已进入环境释放以上的转基因安全评价试验阶段。
2014年6月,将试验材料种植在吉林省公主岭市的吉林省农业科学院的人工温室中,幼苗长至5叶1心期进行取样,每种试验材料取长势基本一致的6个单株的最上部完全展开叶片,混合作为一个样本。液氮冷冻后,迅速放入干冰中运输至实验室,-80℃保存。
1.2 玉米叶片总蛋白提取
将叶片去主脉后剪成1 cm的小段,称取0.5 g叶片加入50 mg PVPP进行充分研磨;加入7.5 mL经-20℃预冷的蛋白提取缓冲液Ⅰ,冰浴研磨5 min,转入预冷的10 mL离心管中,-20℃放置1 h;4℃,13 000 r/min离心45 min,弃上清;加入7.5 mL经-20℃预冷的蛋白质提取液Ⅱ,悬浮振荡,-20℃放置1 h;4℃,13 000 r/min离心45 min,弃上清,重复2次;加入7.5 mL 80%预冷的丙酮,充分悬浮后,离心去上清;沉淀真空干燥并称重。每种材料各重复3次,获得的蛋白用Bradford法[26]测定浓度后准备进行蛋白质双向电泳分析。
1.3 蛋白质等点聚焦
采用长度24 cm,pH 3—10的线性干胶条(Biorad,美国),取蛋白提取液与水化缓冲液(8 mol·L-1Urea、2 mol·L-1硫脲、4% CHAPS、0.002%溴酚蓝,用前加入65 mmol·L-1DTT和0.2% IPG Buffer)混合至总体积450 μL(含蛋白质约为1 200 μg),采用被动水化的方法处理18 h。将水化后的胶条转移到等电聚焦槽上(胶条酸性端置聚焦盘的“+”端,碱性端置聚焦盘“-”端)。每根胶条加入3 mL矿物油,将润湿的滤纸1/3处放在胶条的两端夹上电极夹,扣紧聚焦盘,等点聚焦程序和参数设置参照Biorad产品说明书。等点聚焦结束后,将胶条置于平衡缓冲液(6 mol·L-1Urea、75 mmol·L-1Tris-HCl(pH 8.8)、2% SDS、25%甘油、0.002%溴酚蓝)振荡平衡2次,每次15 min。第一次平衡在缓冲液中加入1% DDT;第二次平衡在缓冲液加入2.5%碘乙酰胺(IAM)。平衡完毕后取出胶条轻轻用平衡缓冲液润洗,然后去除多余的平衡缓冲液,准备SDS-PAGE电泳。
1.4 SDS-PAGE
将平衡好的胶条置于第二向凝胶上方,排除气泡使二者紧密接触,用1%琼脂糖凝胶封固胶条,接通电源。第一阶段1 W/gel运行1 h;第二阶段13 W/gel运行直至溴酚蓝到达凝胶底端。双向电泳结束后,SDS-PAGE凝胶置于固定液(10%冰醋酸、40%甲醇、50%超纯水)中固定3 h;凝胶固定后,超纯水冲洗3次,每次15 min;加入考马斯亮蓝染色液染色(0.12% G-250、10%硫酸铵、10%磷酸、20%甲醇)振荡过夜;超纯水脱色至背景满意为止。
1.5 质谱鉴定与数据库分析
Image Scanner扫描仪扫描脱色后的凝胶,用Biorad PDQuest 8.0软件分析不同玉米叶片材料的凝胶图谱,采用Yang等[4]的方法,使用蛋白相对表达体积(% Vol)来表述每一个匹配蛋白质点的表达丰度。差异蛋白质的表达丰度分析分别以Z58和ZD958材料的凝胶作为参考胶,只有蛋白质点在至少2个重复内被发现才被认为是真实的蛋白质点。定义平均相对表达丰度上调(出现)或下调(消失)差异大于2倍(ratio>2),并且在统计学T检验上<0.05的蛋白质为差异蛋白质点。差异蛋白质点使用液质联用质谱仪LC-Q-TOF-MS 6520(Agilent Technologies,USA)进行分析。使用NCBI玉米数据库(NCBI,Taxonomy:,Entry:212235,2015/01/20)对蛋白质进行检索。蛋白检索结果的显著性判别标准:一般认为PMF(Peptide Mass Fingerprinting,肽指纹图谱)在MS/MS检索中离子分数≥88分说明该蛋白的鉴定结果有效,其他一些辅助标准包括至少10%的序列覆盖率以及需要有3个肽段的匹配[27]。
1.6 GO和KEGG分析
采用BLAST(https://blast.ncbi.nlm.nih.gov)方法对质谱鉴定出的特异性蛋白(E value≤10−5)与NCBI non-redundant(Nr)蛋白质数据库进行比较,确定相应的蛋白质并获得对应的基因信息,用DAVID方法对这些表达有差异的基因进行基因功能注释(gene ontology,GO)分析[28],对差异蛋白功能及代谢途径进行KEGG通路分析[29]。
2 结果
2.1 差异表达蛋白的分离及鉴定
为了比较不同转基因抗虫玉米蛋白质组差异,对供试材料的叶片总蛋白进行了双向电泳分析。结果显示,各受试材料均可检测到超过1 200个清晰蛋白点(图1)。
将电泳胶图扫描后使用Biorad PDQuest8.0软件对每个转基因材料及其对照(1#和6#、4#和6#、2#和5#、3#和5#)进行比较,对电泳凝胶上相对表达体积超过2倍的差异蛋白质点进行质谱仪分析,成功鉴定玉米基因组蛋白61个(图2、表1)。进一步分析特异蛋白点(只在转基因与对照的一个材料中出现)和差异蛋白点发现,这些蛋白点中核酮糖二磷酸羧化酶相关蛋白点15个,ATP合酶类蛋白点4个,放氧增强蛋白点13个,丙酮酸磷酸双激酶相关蛋白点3个,GADPH酶相关蛋白点2个,类萌发素蛋白相关蛋白点4个,草铵膦乙酰转移酶相关蛋白点2个(3#和4#),其他蛋白点6个,未知功能蛋白点10个。这些蛋白所涉及的生化反应过程主要集中在光合作用、碳固定和能量代谢等方面。根据不同蛋白的功能以及质谱检测结果,对各类蛋白在不同转基因材料中变化的趋势进行了汇总分析(表2)。结果显示,在4种转基因玉米材料中发现被认为参与植物对逆境胁迫响应的类萌发素类蛋白均有显著的表达升高;而只有质体蓝素钠一类蛋白在4#材料中发生了表达显著降低;涉及光合作用及碳固定的相关酶类,如核酮糖二磷酸羧化酶、ATP合成酶、丙酮酸磷酸双激酶等均发生了表达显著上调。在同为ZD958背景的1#和4#材料中,包括类萌发素蛋白、核酮糖二磷酸羧化酶、PSⅡ的放氧增强蛋白的表达均发生了显著的升高;而在同为Z58背景的2#和3#材料中,包括ATP合成酶、放氧增强蛋白、类萌发素蛋白、肽基脯氨酰顺反式异构酶等蛋白的表达发生了显著升高。可以看出遗传背景相同的转基因玉米材料(1#和4#、2#和3#)在转入不同抗虫Bt基因后,其所引起的蛋白水平上的变化也存在着一定的一致性。而不同遗传背景的转基因材料(3#和4#)转入相同抗虫Bt基因后,其蛋白水平上的表达变化则差异较大。

图2 转基因玉米及其对照叶片蛋白质的双向电泳对比分析
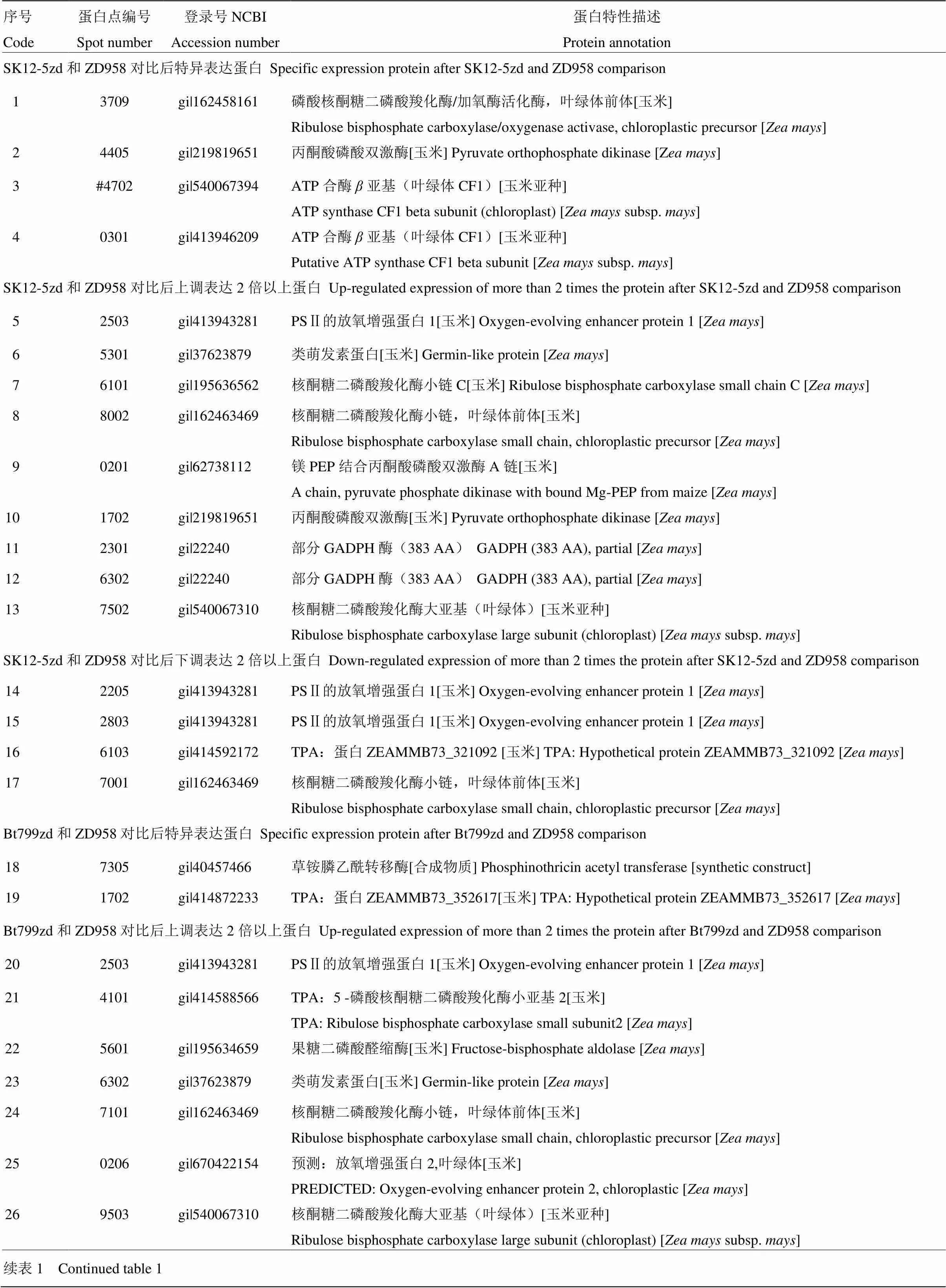
表1 转基因玉米叶片与对照的差异表达蛋白
此外,对4份转基因材料获得的特异蛋白点与转基因材料中所转入的Bt蛋白序列进行比对,发现了特异蛋白点#3802、#4719、#3707、#3708分别对应Bt蛋白cry1Ab、cry1Ie、mcry1Ac、mcry1Ac(图1)。说明转基因材料中转入的Bt基因在玉米中能够正常表达,并且这些Bt蛋白能够通过双向电泳被分离和鉴定,最终体现在蛋白质组分析所获得的特异蛋白点中。
2.2 差异蛋白的GO分析和KEGG分析
虽然通过BLASTx对试验所得的差异蛋白点进行了初步鉴定,获得了蛋白信息,但这些蛋白在是否在细胞组成、生物途径和分子功能上具有共性,是否富集于某一代谢途径,仍无法准确推断。通过对这些差异蛋白质的生物学功能和代谢通路富集分别进行GO(图3)和KEGG分析(图4)。发现转基因材料与对照材料相比虽有差异,但也主要集中在光合作用相关蛋白,并未发现特殊蛋白或特殊代谢途径的明显改变。SK12-5zd与ZD958比较发现,共有5个不同生物途径的相关蛋白发生了差异表达,分别为光合作用(photosynthesis)相关蛋白3个、还原型戊糖磷酸循环途径(reductive pentose-phosphate cycle)相关蛋白2个、钙离子结合(calcium ion binding)蛋白2个、不同细胞组成相关蛋白4个(具体为膜外蛋白(extrinsic component of membrane)2个和光合系统Ⅱ放氧复合体(photosystem Ⅱ oxygen evolving complex)相关蛋白2个。
Bt799zd与ZD958比较发现,有7个不同生物途径相关蛋白发生了差异表达,分别为光合作用相关蛋白4个,光呼吸(photorespiration)相关蛋白质2个和碳固定途径(carbon fixation)相关蛋白2个;发现4个不同分子功能的蛋白发生了差异表达,分别为2个核酮糖-二磷酸羧化酶的活性蛋白(ribulose- bisphosphate carboxylase activity)和2个单加氧酶活性蛋白(monooxygenase activity);按照细胞组成分析也发现了4个蛋白发生了差异表达,分别为膜外蛋白(extrinsic component of membrane)2个和光合系统Ⅱ放氧复合体(photosystem II oxygen evolving complex)相关蛋白2个。
IE034z与Z58比较发现,有4个不同生物途径相关蛋白发生了差异表达,均属于光合途径相关蛋白;有5个细胞组成相关蛋白发生了差异表达,分别为膜外蛋白3个和光合系统Ⅱ放氧复合体相关蛋白2个;按照分子功能分析则发现了钙离子结合的蛋白发生了差异表达3个。

表2 转基因玉米差异表达蛋白汇总
Bt799z与Z58比较发现,有4个生物途径相关蛋白发生了差异表达,均属于光合途径相关蛋白;按照细胞组成分析有6个蛋白发生了差异表达,分别为膜外蛋白3个和光合系统Ⅱ放氧复合体相关蛋白3个;还发现有3个分子功能为钙离子结合的蛋白发生了差异表达。
通过2倍上下调控表达水平来筛选转基因材料与其非转基因对照材料的差异蛋白质进行KEGG分析则发现,在SK12-5zd与ZD958的比较中,差异蛋白在光合生物碳固定途径(carbon fixation in photosynthetic organisms)中显著富集;而在Bt799zd和ZD958的比较中,差异蛋白则分别在光合生物碳固定途径、光合作用和碳代谢(carbon metabolism)几个代谢途径中显著富集。在IE034z和Z58以及Bt799z和Z58的比较中均为光合作用代谢途径中的差异蛋白显著富集(图4)。

图3 转基因玉米叶片与其对照的差异蛋白GO分析
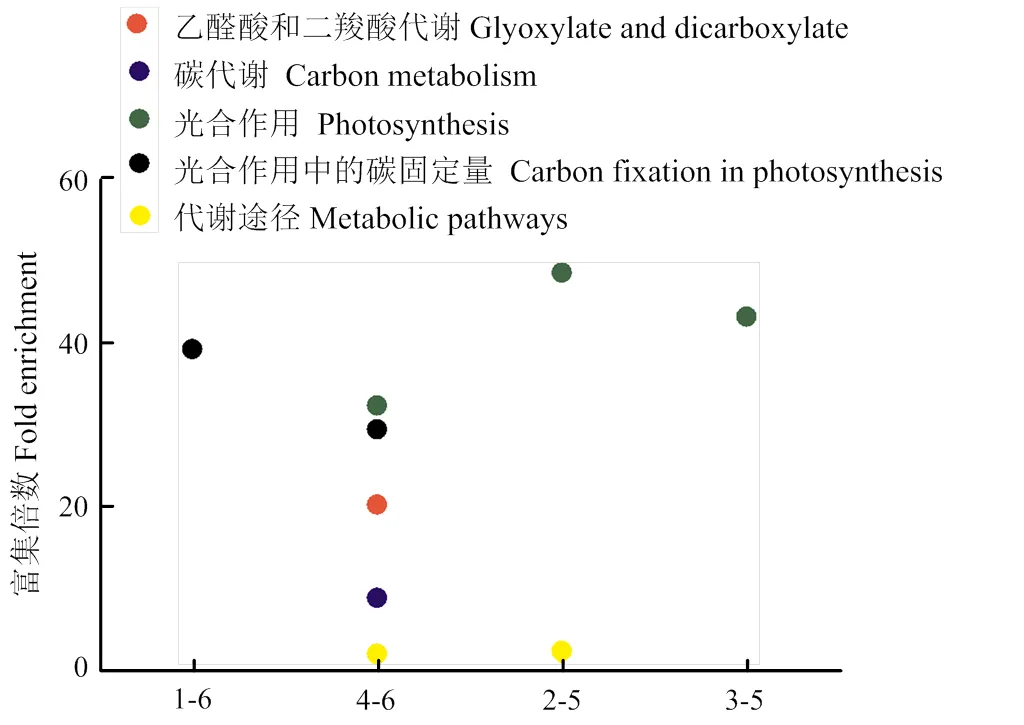
图4 转基因玉米叶片与其对照的差异蛋白KEGG分析
3 讨论
随着转基因植物的研发和推广,转基因生物及其产品的分析鉴定,以及转入基因的直接表达产物和间接调控产物对转基因生物生长发育、生态环境、食物链及人类食品安全的影响越来越受到关注。由于现有转基因生物中多数外源基因的表达或调控产物都是蛋白质,因而在蛋白质水平上对转基因生物进行评价显得尤为重要,蛋白质组学的兴起和发展为在蛋白质水平上对转基因生物进行评价提供了可能[30-31]。
本研究对4种转基因抗虫玉米材料及其非转基因对照玉米材料进行了蛋白质组分析。每份材料经双向电泳凝胶分离后均可检测到1 200个蛋白质点,利用PDQuest8.0软件对蛋白质组表达谱进行分析,已检测到4份转基因材料与对照之间差异表达的Bt杀虫蛋白,还鉴定出61个表达量发生显著变化的蛋白质点,主要集中于光合作用、同化物的积累及能量代谢过程。不同转基因材料中均检测到光合生物碳固定途径中的差异蛋白,如光合作用中决定碳同化速率的关键酶——核酮糖1,5二磷酸羧化酶、C4植物光合作用途径中二氧化碳固定的关键酶——丙酮酸磷酸双激酶、放氧增强蛋白;植物中对逆境响应的类萌发素蛋白、以及ATP合成酶类等在4个转基因材料中均有显著的上调表达,这些差异蛋白不仅是遗传背景的差异,可能也与转基因玉米具有抗虫性相关,而GO分析和KEGG分析的结果也证实了上述差异的存在。理论上光合作用的增强,生物体抗逆能力增强,生长会更健壮,植物的光合能力也会提高,这有待于后期对材料的观察和分析;同时光合作用和碳固定过程增强,均涉及大量的能量物质流动,这些能量物质的去向与外源蛋白的表达是否存在关联,有待深入分析。
本研究发现类萌发素蛋白在所有转基因材料中均有显著的上调表达,该蛋白是在植物中普遍存在的位于胞外基质的可溶性糖蛋白,绝大多数为稳定的低聚物,其作用为参与植物对逆境胁迫的响应。这类蛋白的上调表达可能预示着抗虫转基因玉米材料中,除依赖表达针对靶标生物的毒蛋白已达到抗虫的目的外,还可能存在某种共有的变化机制,对其他逆境胁迫响应产生作用,而具体作用机制需通过后续的逆境胁迫响应评估及分析深入研究。
转基因作物的蛋白质组差异一方面是由于外源基因的插入造成的,另一方面还有可能由环境因素造成[7]。本研究材料均在温室内培养至5叶1心期,极大程度地避免了环境因素的影响,其结果所反应出的差异应主要为外源基因插入造成的蛋白质组差异。在此基础上,发现受体材料的遗传背景可能影响转基因材料的蛋白表达差异。本研究中所用的转基因自交系材料均是经过高世代选育获得的稳定材料,其基因组差异极小,因此,在与其回交亲本材料比较中所发现的蛋白质差异,应主要来源于外源基因的插入及表达所引起的蛋白质组变化。研究结果也证明在相同遗传背景的转基因材料中,其蛋白表达变化的一致性较高,而经过杂交配制的转基因杂交种材料,由于杂交过程中的不确定性,外源基因的表达和杂交过程均会对蛋白质组产生影响,因此导致检测出了更多的差异表达蛋白,本研究结果也再次证实了不同遗传背景但转入相同基因的材料间(3#和4#)其蛋白水平上的表达差异较大。这与抗虫玉米Mon810的相关蛋白质组的比较分析结果基本一致,即在转基因玉米个体的蛋白质表达谱之间没有显著的个体差异,其主要差异源于基因组遗传背景的差异[5-7]。
抗虫转基因玉米生物安全评价过程中非常重视其食用和饲用安全性。综合分析本研究结果,在4份转基因玉米材料中并没有发现影响其食用或饲用安全的新蛋白,未发现植物生理、生化和代谢过程中发挥重要功能的蛋白质或致敏和毒素蛋白质产生或发生明显的表达变化。由于蛋白质是具有生物功能的特殊大分子物质,转基因植物的食用安全性变化会直接体现在蛋白质水平上。本研究表明抗虫转基因玉米在蛋白质水平上并未产生可检出的影响其安全性的蛋白成分。转基因材料中发生变化的蛋白多集中于光合作用、同化物的积累及能量代谢过程,这些过程也与转基因材料安全性关系小,并非影响转基因材料安全性的非预期效应。
4 结论
SK12-5zd、IE034z、Bt799z、Bt799zd等4种转基因抗虫玉米材料与对照蛋白质组相似性较高,仅有少数几个与光合作用、碳固定和ATP合成途径等基础代谢过程相关基因出现上调表达,但未见基因的异常表达。测试的4种转基因材料在蛋白质水平未发生影响其安全性的非预期效应。
[1] Cellini F, Chesson A, Colquhoun I, Constable A, Davies H V, Engel K H, Gatehouse A M, Kärenlampi S, Kok E J, Leguay J J. Unintended effects and their detection in genetically modified crops., 2004, 42(7): 1089-1125.
[2] 赵艳, 李燕燕. 组学技术评价转基因农作物的非预期效应. 遗传, 2013, 35(12): 1360-1367.
Zhao Y, Li Y Y. Unintended effects assessment of genetically modified crops using omics techniques., 2013, 35(12): 1360-1367. (in Chinese)
[3] Arthur J W, Wilkins M R. Using proteomics to mine genome sequences., 2016, 3(3): 393-402.
[4] Yang P, Li F, X J, Wang X, Chen H, Chen F, Shen S H. Proteomic analysis of rice () seeds during germination., 2007, 7(18): 3358-3368.
[5] Coll A, Nadal A, Rossignol M, Puigdomènech P, Pla M. Proteomic analysis of MON810 and comparable non-GM maize varieties grown in agricultural fields., 2011, 20(4): 939-949.
[6] Albo A G, Mila S, Digilio A G, Motto M, Aime S, Corpillo D. Proteomic analysis of a genetically modified maize flour carrying Cry1Ab gene and comparison to the corresponding wild-type., 2007, 52(4): 443-455.
[7] Zolla L, Rinalducci S, Antonioli P, Righetti P G. Proteomics as a complementary tool for identifying unintended side effects occurring in transgenic maize seeds as a result of genetic modifications., 2008, 7(5): 1850-1861.
[8] Wang Y, Xu W, Zhao W, Hao J, Luo Y, Tang X, Zhang Y, Huang K. Comparative analysis of the proteomic and nutritional composition of transgenic rice seeds with Cry1ab/ac genes and their non-transgenic counterparts., 2012, 55(2): 226-233.
[9] Xue K, Yang J, Liu B, Xue D. The integrated risk assessment of transgenic rice: A comparative proteomics approach., 2012, 135(1): 314-318.
[10] Barbosa H S, Arruda S C, Azevedo R A, Arruda M A. New insights on proteomics of transgenic soybean seeds: evaluation of differential expressions of enzymes and proteins., 2012, 402(1): 299-314.
[11] Storer N P, Thompson G D, Head G P, Hutchison W, Gassmann A. Application of pyramided traits against Lepidoptera in insect resistance management for Bt crops., 2012, 3(3): 154-162.
[12] Xu L, Wang Z, Zhang J, He K, Ferry N, Gatehouse A M R. Cross-resistance of Cry1Ab-selected Asian maize borer to other Cry toxins., 2010, 134(5): 429-438.
[13] Li H, Olson M, Lin G, Hey T, Tan S Y, Narva K E.Bacillus thuringiensis Cry34Ab1/Cry35Ab1 interactions with western maize rootworm midgut membrane binding sites., 2013, 8(1): e53079.
[14] 周宗良, 林智敏, 耿丽丽, 苏军, 束长龙, 王锋, 朱彦明, 张杰. 水稻中cry1Ah1基因密码子优化方案的比较. 生物工程学报, 2012, 28(10): 1184-1194.
Zhou Z L, Lin Z M, Geng L L, Su J, Shu C L, Wang F, Zhu Y M, Zhang J. Comparison of codon optimizations of cry1Ah1 gene in rice., 2012, 28(10): 1184-1194. (in Chinese)
[15] Guo Y Y, Tian J C, Shi W P, Dong X H, Romeis J, Naranjo S E, Hellmich R L, Shelton A M. The interaction of two-spotted spider mites, Tetranychus urticae Koch, with Cry protein production and predation by Amblyseius andersoni (Chant) in Cry1Ac/Cry2Ab cotton and Cry1F maize., 2016, 25(1): 33-44.
[16] 李欣, 黄昆仑, 朱本忠, 唐茂芝, 罗云波.利用“组学”技术检测转基因作物非期望效应的潜在性. 农业生物技术学报, 2005, 13(6): 802-807.
Li X, Huang K l, Zhu B z, Tang M z, Luo Y b. Potentiality of “Omics” techniques for the detection of unintended effectsin genetically modified crops., 2005, 13(6): 802-807. (in Chinese)
[17] Mathur C, Kathuria P C, Dahiya P, Singh A B. Lack of detectable allergenicity in genetically modified maize containing “Cry” proteins as compared to native maize based on in silico & in vitro analysis., 2015, 10(2): e0117340.
[18] Agapito-Tenfen S Z, Vilperte V, Benevenuto R F, Rover C M, Traavik T I, Nodari R O.Effect of stacking insecticidal cry and herbicide tolerance epsps transgenes on transgenic maize proteome., 2014, 14: 346.
[19] Hoss S, Menzel R, Gessler F, Nguyen H T, Jehle J A, Traunspurger W. Effects of insecticidal crystal proteins (Cry proteins) produced by genetically modified maize (Bt maize) on the nematode., 2013, 178: 147-151.
[20] Hendriksma H P, Kuting M, Hartel S, Nather A, Dohrmann A B, Steffan-Dewenter I, Tebbe C C. Effect of stacked insecticidal Cry proteins from maize pollen on nurse bees () and their gut bacteria., 2013, 8(3): e59589.
[21] Zhang L, Huang F, Rogers Leonard B, Chen M, Clark T, Zhu Y C, Wangila D S, Yang F, Niu Y. Susceptibility of Cry1Ab maize-resistant and susceptible strains of sugarcane borer (Lepidoptera: Crambidae) to four individual Cry proteins., 2013, 112(3): 267-272.
[22] Li Y, Meissle M, Romeis J. Use of maize pollen by adult(Neuroptera: Chrysopidae) and fate of Cry proteins in Bt-transgenic varieties., 2010, 56(2): 157-164.
[23] 王江, 武奉慈, 刘新颖, 冯树丹, 宋新元. 转基因抗虫耐除草剂复合性状玉米‘双抗12-5’对亚洲玉米螟的抗性及对草甘膦的耐受性研究. 植物保护, 2016, 42(1): 45-50.
Wang J, Wu F C, Liu X Y, Feng S D, Song X Y. Evaluation of transgenic maize ‘Shuangkang 12-5’ with complex traits of insect- resistance and glyphosate-resistance for the resistance toand tolerance to glyphosate., 2016, 42(1): 45-50. (in Chinese)
[24] 刘新颖, 王柏凤, 周琳, 冯树丹, 宋新元. 转基因抗虫玉米IE09S034种植对田间大型土壤动物多样性的影响. 作物杂志, 2016(1): 62-68.
Liu X Y, Wang B F, Zhou L, Feng S D, Song X Y. Effects ofon soil macrofauna diversity in transgenic corn IE09S034 fields., 2016(1): 62-68. (in Chinese)
[25] 刘新颖, 王柏凤, 王江, 冯树丹, 宋新元. 转基因抗虫玉米叶片残体降解对土壤动物群落结构的影响. 植物保护学报, 2016, 43(3): 384-390.
Liu X Y, Wang B F, Wang J, Feng S D, Song X Y. Effects of leaf residue decomposition of-transgenic insect-resistant maize on community structure of soil animals., 2016, 43(3): 384-390. (in Chinese)
[26] Zor T, Selinger Z. Linearization of the Bradford protein assay increases its sensitivity: theoretical and experimental studies., 1996, 236: 302-308.
[27] 于涛, 李耕, 刘鹏, 董树亭, 张吉旺, 赵斌, 柏晗.玉米早期发育阶段粒位效应的蛋白质组学分析. 中国农业科学, 2016, 49(1): 54-68.
Yu T, Li G, Liu P, Dong S T, Zhang J W, Zhao B, Bai H.Proteomics analysis of grain position effects during early developmental stages of maize.,2016, 49(1): 54-68. (in Chinese)
[28] Huang D W, Sherman B T, Lempicki R A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources., 2009, 4(1): 44-57.
[29] Kanehisa M, Goto S, Kawashima S, Okuno Y, Hattori M. The KEGG resource for deciphering the genome., 2004, 32(Database issue): D277-D280.
[30] Pan Y H. Application prospects of proteomicsin detection and study of genetically modified organisms., 2010, 12(1): 31-34.
[31] Cho J I, Lee G S, Park S C.Global status of GM crop development and commercialization., 2016, 43(2): 147-150.
(责任编辑 李莉,岳梅)
assessment of the unintended effects of four genetically modified Maize varieties by Proteomic approach
Hao WenYuan1,2, Li FeiWu2, Yan Wei2, Li CongCong2, Hao DongYun1,2, Guo ChangHong1
(1College of Life Sciences and Technology, Harbin Normal University, Harbin 150025;2)
【】Unintended effects of genetically modified (GM) crops are defined as the unexpected trait changes in GM breeding and research. Over the last decade, much attention has been taken to GM crops’ intended effects that are attributed to the insertion of foreign gene, leaving the unintended effects of GM crops less investigated. In this study, a comparative proteomic study using two-dimensional electrophoresis and mass spectrometry was conducted to assess the high-abundant protein variation, so as to explore possible occurrence of their unintended effects.【】Four insect resistant GM maize varieties, i.e SK12-5zd, IE034z, Bt799z and Bt799zd, which were under approval of GM safety assessment by the Ministry of Agriculture, were planted in a condition-controlled greenhouse together with their corresponding CKs, i.e Zheng58 and Zhengdan958. When seedling growing into 5-leaf stage, the top-leaf blades of each maize variety were sampled and their leaf proteins were extracted. After profiling by two-dimensional electrophoresis, on-gel leaf protein spots of each GM maize were compared against their corresponding CKs using Imager Scanner and PD Quest 8.0 (Biored, USA). The resulted unique and differential protein (with a relative volume greater than 2-fold) spots were isolated from the gel and applied for protein identification by mass spectrometry against the relevant protein database. The identified proteins were analyzed in gene ontology (GO) and KEGG for their cellular functionalities and metabolism pathway enrichment. 【】Through comparison of the unique and differential proteins between each GM maize and its corresponding no-GM CKs among 4 insect resistant GM maize varieties, a total of 61 differential proteins were identified in maize PDB (NCBI) alongside the 4 intended insect resistant gene products. The gene ontology analyses of the identified proteins revealed that they are popular cellular enzymes of fundamental metabolism, i.e Rubisco, ATP synthase, pyruvate orthophosphate dikinase, etc. Only a few genes relating to photosynthesis, ATP synthesis and carbon fixation were observed to be up-expressed, possibly due to the genetic background difference between the GM maize and its CK. KEGG analyses uncovered that, in comparison with the corresponding CKs, the differential proteins of SK12-5zd and Bt799zd were enriched similarly into photosynthesis, carbon fixation and metabolism pathways. In comparison with the CK Z58, the differential proteins of both IE034z and Bt799z were enriched into the photosynthetic metabolism pathway.Four GM maize varieties exhibited extensive similarity over their corresponding wild-type controls. It appeared to be no unintended effects observed. 【】The preliminary results revealed no obvious unintended effects in 4 insect resistant GM maize varieties in respect to high-abundant proteomic assessment.
genetically modified maize; proteomics; unintended effects; two-dimensional electrophoresis (2-DE); differentially expressed protein
10.3864/j.issn.0578-1752.2017.19.003
2017-03-20;接受日期:2017-05-03
国家转基因生物新品种培育科技重大专项(2015ZX08013003)
郝文媛,E-mail:wenyuan_h@163.com。通信作者郭长虹,E-mail:kaku2008@hotmail.com。通信作者郝东云,E-mail:dyhao@cjaas.com
